
金沙江上段浮游藻类和着生藻类群落格局及其与环境因子的关系比较研究
2022-03-10 03:06胡建成宋志斌周佳诚
生态学报 2022年2期
胡建成, 宋志斌, 周佳诚, 谭 路, 唐 涛,*
1 中国科学院水生生物研究所,淡水生态与生物技术国家重点实验室,武汉 430072 2 中国科学院大学,北京 100049 3 西藏大学理学院,拉萨 850000
藻类是河流生态系统的主要初级生产者,它们具有生活周期短,对环境变化响应敏感等特点[1]。一般可按生活习性差异将它们分为浮游藻类和着生藻类两大类。浮游藻类主要在水体中悬浮生活,易受风浪和水流的影响而被动运动;而着生藻类主要营附着或固着生长在浸没于水中的各种基质上。这种生活习性的差异使得浮游藻类和着生藻类具有不同的生境偏好。在源头、低级别或浅水河流中,着生藻类占优势地位;而随着河流级别及水流量的增加,浮游藻类多样性及生物量也会增加[2]。实际上,浮游藻类和着生藻类共存于河流中,它们彼此联系、相互竞争,处于一种动态平衡状态[3—4]。一方面,在特定条件下,浮游藻类和着生藻类可以通过沉降和再悬浮相互转化[5—6]。例如,当水流对基质的冲刷足够大时,着生藻类会被冲刷到水体中成为浮游藻类,而当水流较小时浮游藻类可能沉淀到基质上成为着生藻类。另一方面,由于生长环境不同,浮游藻类和着生藻类对一些环境因子可能存在竞争关系。有研究表明着生藻类对于营养盐具有竞争优势,而浮游藻类对光照具有竞争优势[7—8]。在高度贫营养水体中,着生藻类可能是主要的初级生产者,他们能够吸收水体中的营养盐,并有效减少沉积物中的营养盐向水体中释放,从而影响浮游藻类的生长[8—9]。而随着水体中营养盐的增加,浮游藻类可能会逐渐成为优势类群,通过遮蔽减少着生藻类对于光的吸收[10—11]。然而,以往的研究很少比较同一条河流中浮游藻类和着生藻类的多样性格局。
河流藻类群落的时间动态与环境的季节性变化高度相关[12—14]。水温、光照强度、流速、流量、水质等因素的季节性变化均会引起藻类群落结构的改变。如,雨季洪水冲刷会导致藻类多样性及生物量下降[15—16],同时非点源污染也更容易通过地表径流进入河流[17—18]。而在旱季水流量的降低有利于藻类的稳定和繁殖。此外,浮游藻类和着生藻类对季节性变化的响应也可能存在差异。浮游藻类能借助水流、风浪迁移,避免长时间处于不利环境中;而着生藻类由于生境相对固定,难以躲避环境胁迫的影响[19—20]。因此,相比较而言,着生硅藻对水体环境的持续性变化可能具有更好的响应。然而,浮游藻类和着生藻类与环境因子的关系是否存在季节性的动态变化,相关的研究并不多见。
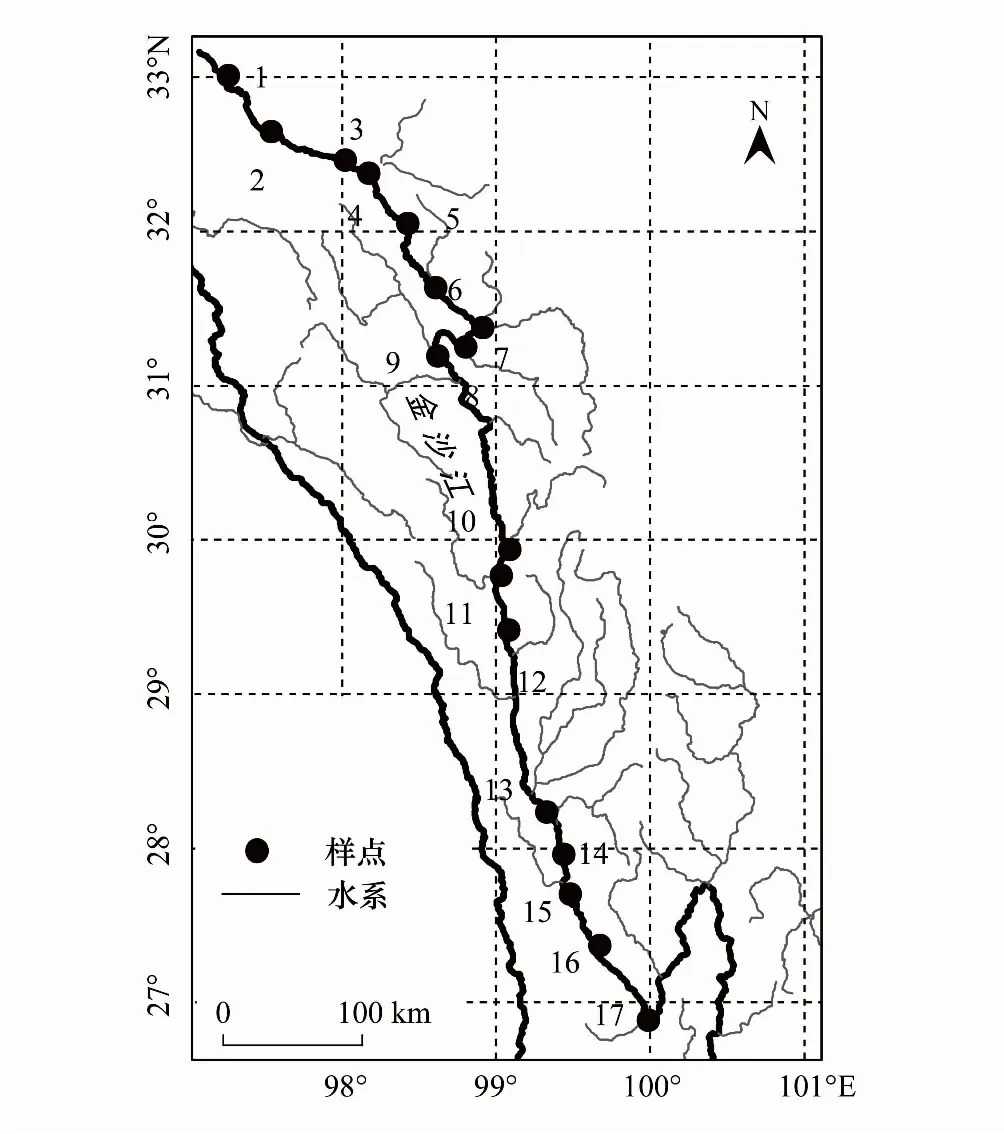
图1 金沙江上段干流调查样点分布图Fig.1 Locations the sampling sites in the upper reaches of Jinsha River采样点:1直门达,2奔达乡, 3 洛须镇, 4麻呷乡,5卡松渡乡,6岗托水文站,7赠曲河口,8金沙乡,9波罗乡,10巴塘河口,11竹巴龙乡,12苏洼龙,13奔子栏,14江东村,15五境乡,16上江,17石鼓镇
金沙江上段地处横断山区,人类干扰较少,河流基本处于自然状态,是生物多样性的热点区域。该区域内河段径流主要以冰雪融化和降雨为主,存在明显季节变化[21],这为探讨河流藻类群落格局的季节性差异提供了理想场所。本研究旨在探究金沙江上段干流浮游藻类和着生藻类的群落结构是否存在差异,以及环境因子的时间动态是否会导致浮游藻类和着生藻类群落具有不同的季节性变化。回答这些问题有助于了解浮游藻类和着生藻类在河流生态系统中季节动态的差异,为河流生物多样性的管理实践提供科学依据。
1 材料与方法
1.1 研究区域及样点设置
金沙江上段始于青海省玉树直门达,流经四川、西藏,至云南石鼓镇,全长965 km,流域面积2.6×105km2[22]。该区域内地貌复杂多样,河谷较深,两岸陡峭,高差较大,分布有高山冰川、高寒湿地等。流域水系发达,具有含沙量大、水能资源丰富等特点,降水多集中在夏季(7—8月)[21, 23]。该地区以藏族居民为主,经济产业以农牧、畜牧、民族手工业、旅游业为主,工业发展水平不高。
本研究分别于2019年9月底(秋季)和2020年7月中旬(夏季)对金沙江上段干流的17个样点展开调查。金沙江上段夏季处于洪水期,水体含沙量大,藻类生物量较低;而到了秋季水流速减缓,含沙量降低,藻类生物量也逐渐增加。所有17个样点中(图1),1个位于青海省境内、11个位于四川省境内,5个位于云南省境内,经纬度跨度为97°14′—99°57′ E, 26°53′—33°00′ N,海拔范围在1816—3490 m。每个样点采集浮游藻类和着生藻类标本,并测定相应的水体理化参数。
1.2 样品采集与测定
考虑到金沙江水体流速、含沙量,以及不同季节水流量差异等特点,适当加大了采水量,并且根据不同季节实时调整采样量,确保能够尽可能全面地采集到浮游藻类和着生藻类。
浮游藻类使用5 L采水器于距水面0.5m处取2—5 L水现场加鲁哥氏液固定。带回实验室经48 h沉淀后浓缩至50 mL,加4%的福尔马林溶液保存以备镜检。着生藻类以纵向100 m河段作为每个样点的调查范围。在河段内随机选取5个位点,每个位点随机挑选3个代表性基质,每个样点共计采集15个基质。具体而言,使用一个半径为2.4cm的铁圈确定采样区域,然后将圈内的藻类用刷子刷下并清洗数次。每个样点的藻液集中装入同一标本瓶中,并记录刷液体积,现场添加4%福尔马林液保存标本。藻类鉴定首先在400倍光学显微镜下对非硅藻分类以及硅藻总数的统计,非硅藻一般鉴定到属水平;再将部分藻液进行酸处理制作成永久性硅藻干片,并在1000倍油境下对硅藻进行鉴定和计数,一般鉴定到种或变种水平,并计算藻类的分类单元数、密度及相对丰度数据。由于浮游藻类是在水样中获取,因此密度计算单位为“个/L”;而着生藻类是在基质表面获取,因此密度计算单位为“个/cm2”。藻类鉴定参照国内外相关文献和书籍[24—27]。
在秋、夏季分别测定了15项环境因子。海拔(Elevation)用手持式GPS仪 (佳明 Drive51)测定,采样点流速用Global Water FP101 流速仪测定,水温(WT)、溶解氧(DO)、浊度(Turb)电导率(Cond)、pH用便携式多参数水质分析仪(YSI ProPlus)测定。采集 1L水样带回实验室,参照《水和废水监测分析方法》测定氨态氮 (NH3-N)、硝态氮(NO3-N)、总氮 (TN) 、总 磷 (TP)、磷酸盐(PO4-P)、化学需氧量(COD)、可溶性有机碳(DOC)、硅酸盐(SiO2-Si)的含量[28]。
1.3 数据分析
藻类群落结构分析使用基于相对丰度数据的Bray-Curtis相异度距离矩阵进行,分析前根据Pearson相关系数剔除相关性较高的环境因子(r>0.8),并对环境因子进行对数转换,对物种数据进行反正弦平方根转换。
首先,用Mann-Whitney U检验分析各样点浮游藻类和着生藻类密度和物种分类单元数的季节差异;用非度量多维尺度分析(Non-metric Multidimensional Scaling,NMDS)解析各个季节浮游藻类和着生藻类的群落结构特征,并进一步使用置换多元方差分析(Permutational multivariate analysis of variance,PERMANOVA)检验不同类群(浮游、着生)和不同季节(秋、夏季)藻类的差异显著程度。该方法本质是基于F统计的方差分析,依据距离矩阵(Bray-Curtis相异度)对总方差进行分解的非参数多元方差分析方法,使用置换多元方差分析(PERMANOVA)可分析不同分组因素对样品差异的解释度,并使用置换检验进行显著性统计[29]。另外,还用Mann-Whitney U检验分析了各环境因子的季节性差异。
接着,用BIO-ENV分析筛选出与藻类群落显著相关的环境因子组合。BIO-ENV分析能够计算出每一个可能的环境因子组合的欧式距离矩阵,并计算出群落相异性矩阵和不同变量组合之间的相关性,从而得出与群落相关性最高的环境因子组合[30]。
最后,使用Mantel test分析进一步检验藻类群落差异与环境距离的相关性,即群落结构差异沿着环境梯度的距离衰减[31]。Mantel相关性指数r>0,表示样点间藻类群落结构的差异随着环境距离的增加而增加,而当P<0.05时,则表示这种关系是显著性的。考虑到调查样点之间空间跨度较大,因此也考虑了地理距离的影响,使用Partial Mantel test分析检验排除地理距离对环境因子的影响后,环境距离与藻类群落差异的关系。环境距离使用欧氏距离,样点间的地理距离是基于各样点经纬度数据计算得到的最短直线距离。为了保持BIO-ENV分析和Mantel test分析之间的一致性,这里使用BIO-ENV筛选出的环境因子进行分析。
以上分析均在R语言(Version 3.6.3)分析平台中进行,Mann-Whitney U检验使用stats包,置换多元方差分析(PERMANOVA)、非度量多维尺度分析(NMDS)、BIO-ENV和Mantel检验均使用vegan包[32]。
2 结果
2.1 环境因子分析
参照国家《地表水环境质量标准》对金沙江水体状况进行了分析[33]。就氮、磷含量而言,多数样点达到Ⅲ类水质标准及以上;溶解氧含量普遍达到Ⅱ类水质标准及以上。通过差异性检验发现,大部分环境因子在夏、秋季存在显著差异。浊度、流速、溶解性有机碳、水温在夏季显著高于秋季;而总磷、磷酸盐、溶解氧、pH、化学需氧量则在秋季显著高于夏季(表1)。
2.2 浮游藻类和着生藻类群落特征差异
2.2.1藻类物种组成及优势种
本次研究共观察到浮游藻类3门32属126个分类单元,其中秋季有3门25属82个分类单元,夏季有1门19属71个分类单元,硅藻门均为主要类群,相对丰度分别为99.5%、100%。浮游藻类分类单元数在秋、夏季无显著差异(W=165.5,P=0.48)(图2),其中秋季各样点平均分类单元数为18种,金沙乡样点最高,为31种,岗托水文站样点最低,为5种;夏季各样点平均分类单元数为16种,巴塘河口样点最高,为31种,竹巴龙和苏洼龙样点最低,均为10种。着生藻类共观察到3门34属121个分类单元,其中秋季共有3门32属102个分类单元,夏季共有3门20属57个分类单元,硅藻门均为主要类群,相对丰度分别为92.1%、89.4%。着生藻类分类单元数在秋季显著高于夏季(W=244.5,P<0.001)(图2),其中秋季各样点平均分类单元数为26种,金沙乡、增曲河口、奔子栏样点最高,均为33种,奔达乡最低,为16种;夏季各样点平均分类单元数为15种,岗托水文站样点最高,为32种,江东村最低,仅有1种。
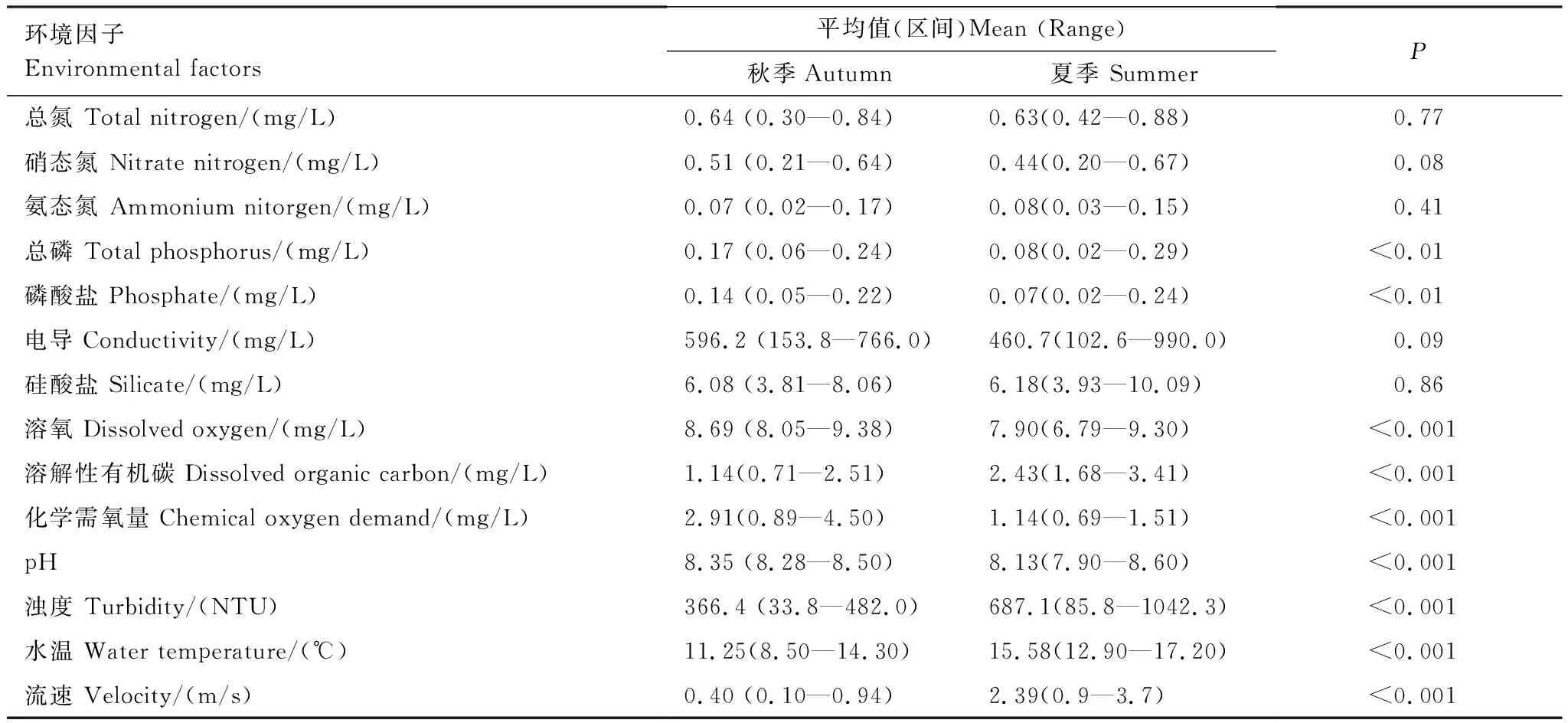
表1 金沙江上段水环境因子季节性差异
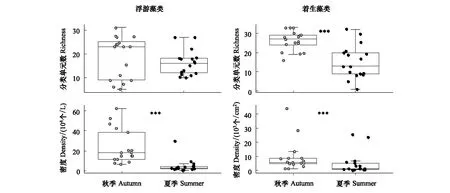
图2 金沙江上段藻类分类单元数和密度箱线图Fig.2 Boxplots of richness and density of algal communities in the upper reach of the Jinsha River“***”代表差异性显著,即P<0.001
分析金沙江上段藻类群落的物种组成特征,发现浮游藻类相对丰度>5%的物种在秋、夏季均有6种,相对丰度>1%的物种分别有14、13种,其余62、52种浮游藻相对丰度<1%。着生藻类相对丰度>5%的物种在秋、夏季分别有3、5种,相对丰度>1%的物种均有20种,其余79、32种着生藻相对丰度<1%。比较不同类群、不同季节藻类群落结构,发现金沙江上段藻类群落物种组成差异较大。浮游藻类在秋、夏季的共有物种为28种,着生藻类在秋、夏季的共有物种为37种;就两类藻而言,秋季浮游藻类和着生藻类共有物种为32种,夏季浮游藻类和着生藻类共有物种为34种。不同藻类的优势种差异也较大,其中只有极小曲壳藻在4个藻类群落中均为优势种,分散菱形藻在秋、夏着生藻类群落中均为优势种(表2)。

表2 金沙江上段浮游藻类和着生藻类优势种相对丰度/%
2.2.2藻类密度
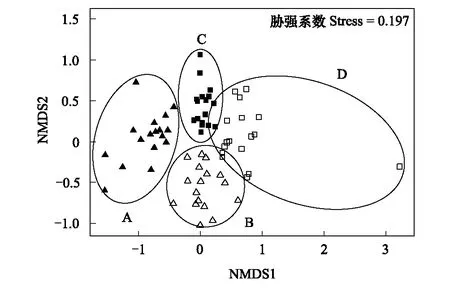
图3 基于藻类群落Bray-Curtis相异度距离的非度量多维尺度分析(NMDS)排序图Fig.3 Non-metric Multidimensional Scaling (NMDS) plot based on Bray-Curtis dissimilarity distances of algal communities A:秋季浮游藻类群落 Planktonic algal community in autumn;B:夏季浮游藻类群落 Planktonic algal community in summer;C:秋季着生藻类群落Periphytic algal community in autumn;D:夏季着生藻类群落 Periphytic algal community in summer
浮游藻类密度在秋季显著高于夏季(W=273,P<0.001)(图2),其中秋季各样点平均密度为2.41×105个/L,波罗乡样点最高,密度为6.25×105个/L,奔子栏样点最低,密度为6.72×104个/L;夏季各样点平均密度为4.84×104个/L,波罗乡样点最高,密度为2.98×105个/L,奔达乡样点最低,密度为6.14×103个/L。着生藻类密度在秋季显著高于夏季(W=223.5,P< 0.001)(图2),其中秋季各样点平均密度为9.43×103个/cm2,金沙乡样点最高,密度为4.41×104个/cm2,奔达乡样点最低,密度为1.25×103个/cm2;夏季各样点平均密度为4.84×103个/cm2,石鼓镇样点最高,密度为2.57×104个/cm2,奔子栏样点最低,密度为3.14×102个/cm2。
2.2.3群落相似度
从非度量多维尺度分析(NMDS)排序图可见,不同类群、不同季节的藻类群落相似度存在差异。不同季节的同一类群在NMDS1轴上分离,同一季节两个类群在NMDS2轴上分离(图3)。置换多元方差分析(PERMANOVA)表明,浮游藻类和着生藻类群落在季节间均存在显著差异(浮游藻类:R2=0.244,P<0.001;着生藻类:R2=0.176,P<0.001),秋季和夏季的藻类群落也存在显著差异(秋季:R2=0.259,P<0.001;夏季:R2=0.179,P<0.001)。
2.3 浮游藻类和着生藻类群落与环境因子的关系差异
用BIO-ENV首先筛选出了与不同季节浮游藻类群落最为相关的环境因子组合,其中秋季为总氮、氨态氮、海拔、流速和可溶性有机碳,夏季为总氮、氨态氮、电导、海拔。Mantel test和partial Mantel test分析结果显示,秋季浮游藻类群落差异随着环境距离的增加而显著增加,但是当排除了地理距离对环境距离的影响后,相关性则不显著(图4,表3)。夏季浮游藻类群落与环境距离、排除了地理距离影响的环境距离均显著相关(图4,表3)。
对于着生藻类群落,BIO-ENV筛选出的环境变量组合在秋季为海拔和硅酸盐,在夏季为总氮、pH和化学需氧量。Mantel test和partial Mantel test分析结果显示,与浮游藻类类似,秋季着生藻类群落差异也与环境距离显著相关,且当排除了地理距离对环境距离的影响后,相关性则不显著(图4,表3)。夏季着生藻类群落差异也与环境距离、排除了地理距离影响的环境距离均显著相关。不同的是,秋、夏季着生藻类群落差异与环境距离的Mantel相关系数均高于浮游藻类(图4,表3)。
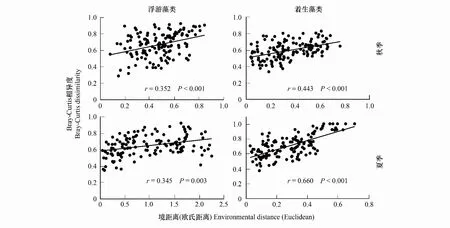
图4 藻类群落差异(Bray-Curtis 相异度)沿着环境距离的变化图(图中显示了每种关系的Mantel相关系数r和显著性P)Fig.4 Changes of algae community dissimilarity (Bray-Curtis dissimilarity) along the environmental distance (Mantel correlation coefficient (r) and significance (P) of each relationship are shown)
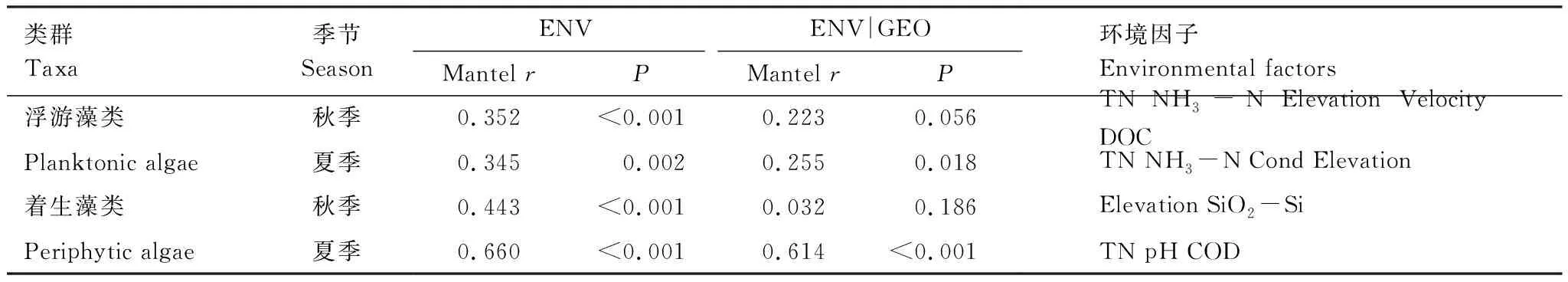
表3 藻类群落差异性(Bray-Curtis相异度)与环境地理距离的相关性(Mantel分析)
3 讨论
本研究发现,金沙江上段不同季节的浮游藻类和着生藻类群落结构均存在显著差异。同时,本研究发现虽然两个类群共有物种较少,但是这些共有物种的平均相对丰度却较高,达到62.7%—85.8%,这从侧面反映了浮游藻类群落和着生藻类群落也存在着紧密联系,即浮游藻类和着生藻类可能相互转化[34]。此外,本研究还发现,金沙江上段浮游藻类和着生藻类群落在秋、夏季的变化既有相似之处也存在差异。相似之处在于硅藻门无论是在物种数还是密度上均为优势门类,且秋季藻类密度均显著高于夏季。在金沙江干流其它河段也有类似的发现[35—37]。不同的是,着生藻类群落的分类单元数在夏季明显低于秋季,而浮游藻类分类单元数的季节变化不显著。导致这一差异的原因可能是由于夏季金沙江水流量大、流速急(调查江段夏季平均流速高达2.39 m/s),大量着生藻类被水流冲刷离开其生境,且难以获得来自上游水域或浮游藻类的补充。另外,夏季异常浑浊的江水(调查江段夏季平均浊度为687.1NTU)使得着生藻类难以获得生长所需的光照,从而限制了它们的生长。相反,浮游藻类却能够获得来自再悬浮的着生藻类,或上游和支流藻类的补充[6,38]。
金沙江上段浮游藻类和着生藻类群落均与环境距离存在显著相关性,即随着环境距离的增加,藻类群落结构差异性(即β多样性)也随之增加。但同时本研究也发现着生藻类群落结构的差异与环境距离的相关性更高,这表明着生藻类可能对于水质变化更为敏感[39]。此外,本研究还发现与秋、夏季浮游藻类群落差异显著相关的环境因子组成较为相似,而与着生藻类显著相关的环境因子在季节间差异较大。这说明,影响着生藻类的主要环境胁迫因子存在季节性差异。地理距离也会对河流环境产生一定的影响[40—41]。当剔除地理距离对环境距离的交互作用后,环境距离与藻类群落结构的相关性发生了变化,即环境距离与夏季藻类群落依然存在显著相关性,而与秋季的相关性却不显著,这是因为秋季环境因子可能存在更显著的空间相关性。实际上,任何季节的环境因子都存在空间相关性,金沙江上段各样点间的海拔梯度可能使得秋季环境因子的空间化格局更突出,秋季藻类群落均受到海拔因素的影响证明了这一点。
本研究结果可为金沙江上段生态管理提供科学依据。研究发现该江段秋、夏季的藻类优势种均以出现在中、贫营养型水体中的清洁指示物种为主,如微小曲壳藻、分散菱形藻、橄榄绿色异极藻、念珠状等片藻等[42],这反映了金沙江上段水体营养水平不高,根据金沙江上段氮、磷含量也能发现这一结果。相较于浮游藻类,本研究发现着生藻类群落差异与金沙江上段环境因子的变化更为相关,因此,宜以着生藻类为主开展河流生态状况调查与评价。此外,金沙江上段雨季流速、可溶性有机碳等环境因子显著增加,因此雨季应重点关注农业非点源污染,并保护河岸带的完整性,减少污染物被冲入河道中;旱季江水中磷营养盐、溶氧等环境因子显著增加,此时应重点控制点源污染的排放。
4 结论
通过比较金沙江上段浮游藻类和着生藻类的群落格局及其与环境因子的关系发现,尽管浮游藻类和着生藻类的群落均以硅藻为主,但是它们的物种组成差异显著,并且在季节间的动态变化也十分不同。着生藻类具有更为明显的季节动态,且对于环境变化具有更好的响应,这表明着生藻类可能比浮游藻类更适用于河流生态系统的管理和评估。此外,也不能忽略地理距离对于藻类群落的影响,在以后的研究中还需要进一步探讨空间过程对于藻类群落构建过程的贡献。
致谢:感谢何智博士、池仕运博士在野外工作中给予的帮助。
猜你喜欢
湖北植保(2022年4期)2022-08-23
湖南水利水电(2021年6期)2022-01-18
建材发展导向(2021年13期)2021-07-28
土壤(2021年1期)2021-03-23
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
中国农村水利水电(2020年4期)2020-06-12
流行色(2019年10期)2019-12-06
福建农业学报(2016年6期)2016-11-01
大众考古(2015年2期)2015-06-26
