
甘蓝、菜豆和玉米不同轮作组合对淡褐土细菌群落和作物产量的影响
2022-03-18 08:36王天乐王晓娟刘恩科史向远
农业环境科学学报 2022年2期
王天乐,王晓娟,刘恩科,史向远
(1.山西农业大学农学院,山西 太谷 030801;2.山西农业大学山西有机旱作农业研究院,太原 030031;3.省部共建有机旱作农业国家重点实验室(筹),太原 030031;4.有机旱作农业山西省重点实验室,太原 030031)
土壤是作物生长繁育的基地,土壤中有着种类繁多、数量巨大的微生物类群,作为土壤生态系统重要组成部分之一,这些微生物联系着作物与土壤,在适应土壤环境的同时影响着作物生长,形成土壤−作物−微生物影响机制,因此,土壤微生物与农业可持续发展关系紧密。细菌在土壤微生物中所占比例最大,是土壤微生物的重要组成部分,是土壤中最活跃的因素,具有数量多、分布广等优势。土壤细菌的群落结构和多样性会因作物种植模式不同产生变化,因此在研究轮作和连作两种重要的作物种植模式时,常用土壤细菌群落的变化来具体说明种植模式不同而产生的差异。
连作障碍多发生于旱地,作物经济效益好且需求量大,导致种植面积大、复种指数高,连作障碍问题日渐突出。结球甘蓝和菜豆因具有较强的生态适应性,而成为我国种植面积较大的蔬菜作物,同时也成为十字花科和豆科作物中的连作障碍高发物种。土壤连作障碍是作物与土壤两个系统之间多种因素相互作用的结果,所产生的障碍主要表现为土壤病虫害严重、作物生长受限、产量下降等。有研究表明,连作障碍是作物与特异微生物共同作用的结果,土壤微生物群落结构变化、有害微生物含量增加是产生连作障碍的主要原因。有研究发现,相较短期连作,长期连作可提高细菌群落多样性指数与OTU数量,这与陈雪丽等研究黑土大豆种植和刘素慧等研究大蒜种植得出的结论不同,说明连作年限和种植作物不同对结果存在较大影响。
轮作是目前国际公认的防治作物连作障碍应用最广泛、最经济有效的措施。相对连作处理,轮作会提高细菌群落多样性指数,显著改变群落结构。旱地多采用禾谷类或豆类作物参与轮作种植,以增加土壤有机质含量、改良土壤团粒结构,缓解连作障碍。玉米是我国重要的粮食作物之一,具有强耐旱性、耐寒性、耐贫瘠性以及极好的环境适应性,也是畜牧业和养殖业的重要饲料来源。糯玉米除具有普通玉米抗逆性强的优点外,还具有较高的营养价值,因此常被用于轮作研究。已有研究表明菜豆与绿叶菜短期轮作可获得高效生产。豆科与禾本科作物适当配合轮作可增加土壤有机质,有利于后作叶菜类作物的种植。
1 材料与方法
1.1 试验区自然条件与试验地概况
试验在山西农业大学旱地农业研究中心阳曲县凌井店河村基地进行,该地海拔1 248.5 m,年平均降水量450 mm,降雨多集中于夏季,其他三季雨水较少,且蒸发量大,为典型的半干旱区,属温带大陆性季风气候,年平均气温6~7 ℃,昼夜温差大,≥10 ℃的活动积温约2 600 ℃,年日照时数2 662 h,无霜期约120 d。该地地势平坦,土壤为黄土质淡褐土,0~20 cm 土层土壤基本理化性质为:有机质14.41 g·kg,全氮1.19 g·kg,全磷0.7 g·kg,全钾20.7 g·kg,pH 7.72。该区域农田种植主要依靠自然降水,主要种植玉米、结球甘蓝、菜豆等作物,一年一种,普遍连作。
1.2 试验材料
供试结球甘蓝(L.L.)品种为“世农307”,菜豆(L.)品种为“红珍珠”,糯玉米(L.)品种为“京科糯2000”。
1.3 试验设计
2018—2020 年连续3 a 定位进行单因素试验,形成玉米−甘蓝−菜豆轮(连)作体系,共10 组处理(表1),每组3 次重复,随机区组排列,每小区种植面积均为30 m。根据当地大田施肥情况施用复合肥,结球甘蓝施用化肥600 kg·hm,并多施粒状过磷酸钙375 kg·hm,总计纯氮108 kg·hm、纯磷77.58 kg·hm、纯钾89.62 kg·hm;玉米和菜豆施用化肥1 200 kg·hm,总计纯氮216 kg·hm、纯磷94.32 kg·hm、纯钾179.24 kg·hm。种植密度为甘蓝33 333 颗·hm,菜豆64 000 株·hm,玉米60 000 株·hm。2020 年试验玉米、菜豆于5月13日播种,甘蓝于6月10日移栽,期间菜豆多次采收(8 月2 日和13 日),所有作物于9 月11日收获,试验田旱作不灌水。

表1 试验的处理设置Table 1 Processing settings for the experiment
1.4 测定项目与方法
于2021 年作物播种前使用小型土钻通过五点取样法采集各处理小区内表层(0~20 cm)土壤,土壤混合后去除石块、根等杂质,使用2 mm 孔径土壤筛过筛,装入封口袋,干冰封存。利用Illumina HiSeq 高通量测序平台对细菌基因V3~V4 区测序,用双末端测序(Paired−End)的方法,构建小片段文库进行测序,通过对Reads 拼接过滤、聚类或去噪,并进行物种注释及丰度分析,揭示样品的物种构成;测定土壤中细菌群落结构、多样性,挖掘样品之间的相似性和差异性。甘蓝每小区随机取20 颗,测定其球状体实际质量;菜豆每小区随机摘取8 株枝上成熟豆角,不同时期3次测量相加后计算其产量;玉米每小区随机取20个,去掉雌穗苞叶连同玉米穗轴一同称质量,最后依次换算成所需面积产量。
1.5 数据处理
采用Excel 2003软件处理数据,SPSS 18.0软件进行统计分析,利用R语言工具绘图。
2 结果与分析
2.1 作物不同轮作组合下土壤细菌16S r RNA 的V3~V4区原始数据分析
通过统计各阶段的序列数、序列长度等参数对数据进行评估。10个处理,3次重复,共30个样品,测序共获得2 398 950 对原始序列(Raw reads),2 386 493条高质量序列(Clean reads),2 334 112 条有效序列(Effective reads),每个样品至少产生232 425 条有效序列,平均产生233 411 条有效序列,平均长度在418~421 bp 之间。稀释曲线用于验证测序数据量是否足以反映样品中的物种多样性,并间接反映样品中物种的丰富程度。由图1 可知,在各样品所含有效序列数范围内,随着样品测序数目的增加,曲线趋于平缓,表明样品序列充分,可以进行数据分析。

图1 样品稀释曲线Figure 1 Each samples rarefaction curves
2.2 作物不同轮作组合下土壤细菌OTU聚类分析
对所有处理在97%序列相似度下得到的土壤细菌OTU 数目进行统计,并绘制Venn 图用以直观地找出不同处理中的共有特征和特有特征的数目,从而结合特征所代表的物种,找出不同环境中的特有微生物。由图2a可知,各处理的OTU数无显著差异,10组处理共同拥有OTU 数目为2 299 个,前茬种植同种作物无特有OTU,所测得细菌分别归属于36 门、97 纲、199目、318科、588属、653种。

图2 各样品特征韦恩图Figure 2 Venn diagram of characteristics of each sample
前茬种植作物为甘蓝时三者共有2 324 个OTU数(图2b);CCC 处理特有OTU 数 为2,分别是OTU1372 Firmicutes 门Bacilli 纲Lactobacillales 目Lactobacillaceae 科属种(厚壁菌门芽孢杆菌纲乳杆菌目乳杆菌科乳杆菌属棒状乳杆菌种)和OTU1893 Firmicutes 门Bacilli纲Bacillales 目Paenibacillaceae 科属uncultured_bacterium 种(厚壁菌门芽孢杆菌纲芽孢杆菌目类芽孢杆菌科类芽孢杆菌属下未定名种);BMC处理特有OTU数为1,是OTU2307 Proteobacteria门Gammaproteobacteria 纲Betaproteobacteriales 目Burkholderiaceae科属uncultured_bacterium 种(变形菌门γ−变形菌纲β−变形菌目伯克氏菌科属下未定名种);两两共有OTU数为9。前茬种植作物为菜豆时三者共有2 315 个OTU 数(图2c);BBB和BMB 处理无特有OTU,CMB 处理特有OTU 数为1,是OTU2133 Actinobacteria 门Actinobacteria 纲Frankiales 目Sporichthyaceae 科属_SCGC_AAA278−O22 种(放线菌门放线菌纲弗兰克氏菌目鱼孢菌科hgcl分支属放线菌SCGC_AAA278−O22 种);两两共有OTU 数为20。前茬种植作物为玉米时四者共有2 314个OTU数(图2d);除BBM处理无特有OTU 外,其他处理均有1 个特有OTU,CCM 处理是OTU2307 伯克氏菌科属下未定名种,CBM处理是OTU2133鱼孢菌科hgcl分支属放线菌SCGC_AAA278−O22种,BCM处理是OTU1893类芽孢杆菌科类芽孢杆菌属下未定名种;其余为两两共有或三者共有。
2.3 作物不同轮作组合下土壤细菌群落Alpha多样性分析
由表2 可知,不同处理土壤细菌多样性指数覆盖率均在99.8%以上。Ace 指数与Chao1 指数变化相同,前茬种植同种作物的处理丰富度指数无显著差异,说明在本试验条件下细菌群落丰富度与轮作末期种植的作物有关联。CCC 处理丰富度指数显著高于BCM 处理,CCC、BCM 处理与其余处理以及其余处理之间均无显著差异;除BMC 和BCM 两处理丰富度指数值低于BBB 处理外,其余处理丰富度指数值均高于BBB 处理。Shannon 指数值显示,前茬种植作物分别为甘蓝和菜豆的处理间均无显著差异;前茬种植作物为玉米的处理间存在显著差异,CCM 处理与BBM处理显著高于CBM 处理;CMC、BMC、CCC、BMB、CCM与BBM 处理均显著高于CBM 处理,BMC、CCC、BMB处理较CBM 处理提高3.6%,CMB、BBB、BCM 处理高于CBM 处理但差异不显著,CMB 处理较CBM 处理提高2.1%。各处理Simpson 指数均在0.997 左右趋近于1,说明细菌群落普遍优势度较高、多样性较低,指数值变化规律与Shannon指数相似。

表2 各处理下土壤细菌Alpha多样性指数Table 2 Bacterial alpha diversity index of soil under different treatments
上述结果说明,不同连作、轮作组合对土壤细菌群落多样性所造成的影响不同,相对连作处理,CBM处理细菌群落多样性指数和BCM 处理细菌群落丰富度指数显著下降,其余处理影响不显著。
2.4 作物不同轮作组合下土壤细菌群落门水平组成结构分析
不同处理土壤细菌门水平的群落组成如图3 所示。相对丰度大的10 个细菌门分别为变形菌门Proteobacteria(25.0%~31.1%)、酸杆菌门Acidobacteria(18.5%~23.8%)、拟 杆 菌 门Bacteroidetes(10.1%~13.7%)、厚壁菌门Firmicutes(9.0%~16.8%)、绿弯菌门Chloroflexi(6.3%~9.1%)、放线菌门Actinobacteria(5.4%~7.1%)、芽单胞菌门Gemmatimonadetes(3.7%~4.9%)、疣微菌门Verrucomicrobia(2.7%~3.6%)、硝化螺菌门Nitrospirae(1.8%~2.4%)、浮霉菌门Planctomycetes(1.3%~1.4%),其在10个处理土壤样品中的相对丰度比例之和均占土壤细菌总数的95%以上,其他相对丰度低的细菌门合并为Others。总体上看,变形菌门、酸杆菌门、拟杆菌门和厚壁菌门4 种细菌类群是各处理土壤中的主要菌群,其相对丰度之和占细菌总数的70%以上。连作和轮作处理对细菌门类的组成无差异,但在群落相对丰度上存在差距。
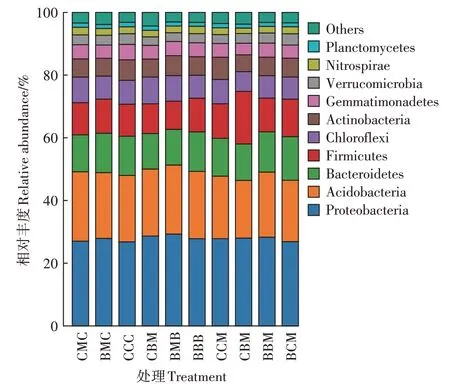
图3 不同处理土壤在细菌门水平的群落组成Figure 3 Community composition of bacteria at phylum level inthe different treatment soils
将不同处理土壤细菌优势门类排名前6 的细菌进行分析,比较不同处理土壤细菌群落组成差异,结果如表3所示。前茬种植作物相同时,CMC 处理绿弯菌门相对丰度显著高于CCC 处理;CMB 处理放线菌门相对丰度显著低于BBB 处理;BCM 处理变形菌门相对丰度显著低于CCM、CBM 和BBM 处理,CBM 处理放线菌门相对丰度显著低于CCM、BBM 和BCM 处理,其余处理间菌门丰度均无显著差异。
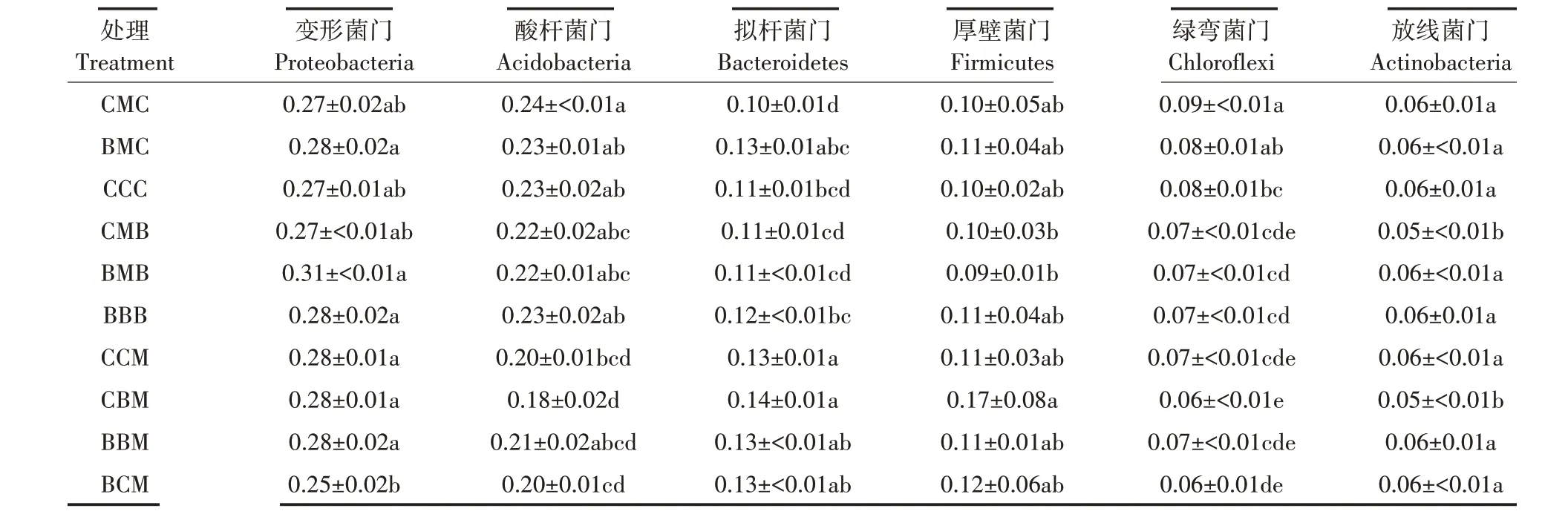
表3 门水平上排名前6的细菌相对丰度差异Table 3 Comparation of relative abundance of Top 6 bacteria on phylum level
变形菌门相对丰度在各处理间差异较小,除BCM 处理显著低于BMC、BMB、BBB、CCM、CBM 与BBM 处理外,其余处理两两间均无显著差异;CMC、BMC 处理略高于CCC 处理。CCC 处理略低于BBB 处理,相对连作处理,CMB、BCM 轮作处理降低变形菌门相对丰度,其余轮作处理(除CMC 处理)均能够小幅度提高菌门相对丰度。
酸杆菌门CBM 处理相对丰度显著低于CMC、BMC、CCC、CMB、BMB 与BBB 处理,BCM 处理显著低于CMC、BMC、CCC 与BBB 处理,CCM 处理显著低于CMC 处理,其余处理两两间均无显著差异;CMC 处理略高于CCC 处理,CMB、BMB 处理略低于BBB 处理。相对连作处理,CBM、BCM 轮作处理能够显著降低酸杆菌门相对丰度,其余轮作处理(除CMC、BMC 处理)降低菌门相对丰度的效果不显著。
拟杆菌门CMC 处理显著低于BMC、BBB、CCM、CBM、BBM 与BCM 处理,CMB、BMB 处理显著低于CCM、CBM、BBM、BCM 处理,CCC、BBB 处理显著低于CCM、CBM 处理,其余处理两两间均无显著差异;CMB、BMB 处理略低于BBB 处理。连作CCC 处理略低于BBB 处理,相对连作处理,CCM、CBM 轮作处理显著提高拟杆菌门相对丰度,BMC、BBM、BCM 处理能够小幅度提高菌门相对丰度,CMC、CMB、BMB 处理能够小幅度降低菌门相对丰度。
厚壁菌门CMB、BMB处理显著低于CBM处理,其余处理两两间均无显著差异,CMC、BMC 处理略高于CCC 处理,CMB、BMB 处理略低于BBB 处理。连作CCC处理略低于BBB处理,相对连作处理,CMB、BMB轮作处理降低厚壁菌门相对丰度,其余轮作处理均能够小幅度提高菌门相对丰度。
绿弯菌门CBM 处理显著低于CMC、BMC、CCC、BMB 与BBB 处理,BCM 处理显著低于CMC、BMC、CCC 处理,CMB、BMB、BBB、CCM、BBM 处理显著低于CMC、BMC 处理,CCC 处理显著低于CMC 处理,其余处理两两间均无显著差异,CMC、BMC 处理略高于CCC 处理。连作CCC 处理略高于BBB 处理,相对连作处理,CMC、BMC 轮作处理提高绿弯菌门相对丰度,CBM 处理显著降低菌门相对丰度,其余轮作处理(除BMB处理)均能够小幅度降低菌门相对丰度。
放线菌门相对丰度在各处理间差异较小,CMB、CBM 处理显著低于其余处理,连作CCC 处理略高于BBB 处理,相对连作处理,CMB、CBM 轮作处理显著降低放线菌门相对丰度,CCM 处理小幅度降低菌门相对丰度,其余轮作处理(除BMB 处理)均能够小幅度提高菌门相对丰度。
2.5 作物不同轮作组合下土壤细菌属水平分布
由图4可以看出,在属水平上10个处理明显分为两大类,CBM处理轮作组合单列为一支,其余9个组合细菌群落结构较为接近,为另一支,说明CBM 处理土壤细菌群落结构显著不同于其余处理,其对土壤细菌群落属水平组成结构的影响大于其他组合。区别于其他轮作组合,CBM 轮作处理乳酸杆菌属()、魏斯氏菌属()、未培养肠杆菌属类群(uncultured_bacterium_f_Enterobacteriaceae)、艾克曼菌属()、大肠杆菌志贺菌(−)、气单胞菌属()和副拟杆菌属()数值较大,超半数细菌属相对丰度较低。

图4 不同处理土壤在细菌属水平的相对丰度聚类热图Figure 4 Abundance cluster heatmap of bacteria at genus level in the different treatment soils
除CBM 处理外,其余9 个组合又可分为3 组,CMC、BBB 和CCM 处理为第一组,BMC、BBM 和BCM处理为第二组,CCC、CMB 和BMB 处理为第三组。相对于第三组,第一组和第二组细菌群落结构差异较小,各组内3 种处理细菌属群落结构分别具有一定的相似性,同时也说明3 组间土壤细菌属群落组成相似性较低。
结果表明,取土样前最后种植作物、同种作物不同轮作顺序、同种作物轮作年限对土壤细菌属群落结构和各物种相对丰度均有影响,其中CBM 轮作处理显著不同于其他9种处理。
2.6 不同轮作组合对产量的影响
在前茬种植作物相同时,分析不同处理对作物产量的影响,结果如表4所示,前茬种植作物相同的3个处理间均无显著差异。前茬种植作物为甘蓝时,CMC处理产量最高,相对CCC、BMC 处理分别增产0.76%、8.30%;前茬种植作物为菜豆时,BMB 处理产量最高;前茬种植作物为玉米时,BBM 处理产量最高,其次为BCM处理,CBM处理产量最低。

表4 不同处理对作物产量的影响Table 4 Effects of different treatments on crop yield
2.7 不同组合的主成分分析
为明确不同处理对土壤细菌群落结构产生的差异,基于OTU 水平绘制主成分分析图。如图5 所示,解释度最大的两个变量命名为PC1 和PC2,两者加和为82.63%,可在此累计方差解释度上解释土壤细菌属群落结构的变化。图中CMB、CBM处理和其余8个处理距离较远,表明CMB、CBM 处理与其余处理间群落结构差异较大,且CMB 和CBM 处理因受不同环境因素影响也存在显著区别。

图5 不同处理下土壤细菌群落结构PCA分析Figure 5 PCA analysis of soil bacterial community structure under different treatments
3 讨论
轮作对土壤细菌群落种类组成无差异,但在群落相对丰度上存在差异。高通量测序结果表明,作物轮作、连作处理中共鉴定出细菌36门、97纲、199目、318科、588 属、653 种,同时还发现许多未被分类的细菌(unclassified bacteria)。
综合所得结果提出合理推测,厚壁菌门乳杆菌属棒状乳杆菌种、厚壁菌门类芽孢杆菌属下未定名种、变形菌门属下未定名种和放线菌门分支属放线菌SCGC_AAA278−O22种等4个种会对土壤产生不利影响,进而阻碍作物生长。放线菌门分支属放线菌SCGC_AAA278 −O22 种存在于CMB 和CBM 处理中,且这两个处理与其他处理相比作物产量最低,此菌种的产生可能是导致轮作产量低于连作的原因。
基于Illumina HiSeq 测序平台,利用双末端测序(Paired−End)的方法,在各处理的覆盖率指数均在99.8%以上的条件下,对不同轮作处理的土壤细菌16S rRNA 序列V3~V4 区进行高通量测序,以揭示样品物种组成的变化。马瑞瑞通过研究芸豆短期连作、轮作对土壤微生物的影响表明,不同连作年限对土壤微生物的影响不完全相同。吴冬琴通过研究甘蓝2、10、15 a不同连作年限对根际土壤微生物的影响进一步表明,土壤中细菌数量呈现先降低再升高再
降低的周期性变化趋势。倪苗等的研究表明豇豆连作与豇豆叶菜轮作处理对土壤细菌多样性的影响较小,轮作能够提高土壤细菌群落丰度。吴宏亮等的研究表明,不同作物轮作与西瓜连作之间的土壤微生物群落多样性指数变化不同,轮作多样性指数既有上升也有下降。连作使百合、黄瓜、辣椒、大豆等作物土壤细菌群落多样性指数与丰富度增加。通过对比前人的研究发现,不同作物轮作、不同连作年限对土壤细菌数量、多样性的影响机制不同。因无关于3 a 内甘蓝连作、菜豆连作以及甘蓝、菜豆、玉米不同轮作组合对土壤细菌多样性方面的影响研究,因此无法形成精准的对比分析。
本研究结果表明,各处理间土壤细菌OTUs 无显著差异;细菌丰富度指数在BCM 处理中数值最低,显著低于CCC处理;细菌Shannon指数在CBM 处理中数值最低,显著低于CMC、BMC、CCC、BMB、CCM 和BBM 处理。综合而言,相对连作处理,轮作CBM 处理细菌群落多样性显著下降,BCM 处理细菌群落丰富度显著下降,其余轮作处理影响不显著,可能是因为CBM、BCM 轮作处理中放线菌门hgcl 分支属放线菌SCGC_AAA278−O22 种的存在导致土壤有益细菌群落大量减少,其原因还需进一步进行分析。
在门水平上,各处理细菌优势门均为变形菌门、酸杆菌门、拟杆菌门、厚壁菌门、绿弯菌门。土壤细菌优势菌门组成与王进闯研究的大豆连作、陈丹梅等研究的作物轮作、徐新雯等研究的烟蒜轮作较为相似。
前茬种植玉米相对于种植甘蓝、菜豆有利于提高拟杆菌门、厚壁菌门相对丰度,降低酸杆菌门、绿弯菌门相对丰度。相对于连作处理,前茬种植玉米时能够在一定程度上提高变形菌门、拟杆菌门、厚壁菌门相对丰度,降低酸杆菌门、绿弯菌门、放线菌门相对丰度;CBM 处理显著降低酸杆菌门、绿弯菌门、放线菌门相对丰度,显著提高拟杆菌门相对丰度;BCM 处理显著降低酸杆菌门相对丰度,CCM 处理显著提高拟杆菌门相对丰度,CMC 处理显著提高绿弯菌门相对丰度,CMB处理显著降低放线菌门相对丰度。
CCM 处理拟杆菌门相对丰度显著高于CCC 处理,CMC 处理绿弯菌门相对丰度显著高于CCC 处理,说明在3 a内甘蓝连作和甘蓝与玉米轮作对土壤菌门影响差异较大。BMC、CBM、BCM 与CMB 处理土壤优势菌门存在差别,说明甘蓝、菜豆、玉米3 种作物不同轮作组合对土壤优势菌门组成存在影响,且前茬种植作物为菜豆时影响较大。丁俊男等研究玉米−大豆不同轮作模式对土壤的影响,结果表明前茬种植豆科有利于改善土壤微生物群落结构,提高微生物群落多样性,玉米−大豆−大豆轮作模式最优。
南丽丽等研究苜蓿后茬轮作4 种不同作物对土壤细菌门水平产生的影响发现,除酸杆菌门相对丰富呈增加趋势,其余优势菌门相对丰度均有下降。有研究表明酸杆菌门适宜生活在酸性环境中,而种植玉米会降低土壤酸度,提高土壤pH,这可能与本试验中CCM、CBM、BBM、BCM 处理酸杆菌门相对丰度下降而其余处理无明显变化有关。CBM、BCM 处理相对丰度显著低于CCC、BBB 连作处理,说明甘蓝、菜豆、玉米3 种作物轮作且前茬种植作物为玉米时能够较大程度提高土壤pH。酸杆菌可降解植物残体,在贫瘠土壤中相对丰度更高,属于寡营养型细菌,可用来判断土壤环境瘠薄程度,这极大可能与CMB、BMB处理酸杆菌门相对丰度低于CCC、BBB连作处理存在相互关联。
变形菌门和酸杆菌门是衡量土壤营养状况的指标,通常土壤中酸杆菌门细菌与变形菌门细菌数量相当,在陆地土壤微生物群落中分布最广、含量最丰富。变形菌门在土壤细菌群落中相对丰度最高,是轮作处理较为常见且重要的细菌门,具有固氮作用和环境适应能力强、抗病能力强等优点,环境变化对其影响较小,属于富营养菌。变形菌门主要研究α−变形菌纲、β−变形菌纲和γ−变形菌纲,在种植豆科作物中有效固氮菌主要是α−变形菌和γ−变形菌[37]。
土壤酸杆菌门相对丰度下降使得其他细菌门竞争压力减小,有助于提高其他细菌门相对丰度如拟杆菌门,拟杆菌门与变形菌门都是富营养型细菌,在营养状态好的土壤中含量较高。整体比较,前茬作物种植玉米有助于改善土壤营养状况。
有研究表明,厚壁菌门随土壤pH 由4 升高到8,其相对丰度呈明显增加趋势。CBM 处理厚壁菌门相对丰度显著高于CMB、BMB 处理,这与适宜生活在酸性环境中的酸杆菌门相对丰度变化相反,说明CBM 处理土壤pH 最高,这可能是造成CBM 处理土壤细菌属显著区别于其他处理的原因。玉米适宜生活在土壤pH 为5.8~7.5 的环境中,糯玉米抗酸能力强耐盐碱能力差。
绿弯菌门作为自养型有益细菌,具有较强的生态恢复能力和生存优势,可降解受污染沉积物中的多环芳烃,驱动生物地球化学循环。本研究轮作组合中CMC 处理绿弯菌门相对丰度最高,CBM 处理最低。放线菌门适宜生长在中性偏碱的土壤环境中。有研究表明苜蓿轮作降低了土壤pH,从而降低放线菌门相对丰度。CMB、CBM 处理放线菌门相对丰度显著低于其他处理,结合厚壁菌门相对丰度的变化,说明CBM 处理可能导致土壤pH 升高,CMB 处理可能导致土壤偏酸性。
在属水平上,CBM 处理显著区别于其余9 个处理,CMC、BMC 处理物种组成和丰度与CCC 处理更为接近,CMB、BMB 处理物种组成和丰度与BBB 处理更为接近。主成分分析显示,CBM 处理土壤细菌群落结构显著区别于其他处理,CMB 处理次之,其余8 个处理差异较小。以上说明除CBM 处理土壤细菌群落变化较大外,前茬种植同种作物时,轮作处理细菌属较为相似,并与连作处理差异较大。
前茬种植作物为甘蓝时,3 个处理产量表现为CMC 处理>CCC 处理>BMC 处理,这与种植后期BMC处理小区虫害严重、测产误差较大有关。前人研究表明,玉米−菜豆−马铃薯−甘蓝轮作组合的甘蓝产量最高,菜豆加入甘蓝轮作组合有利于提高产量,甘蓝连作导致产量下降,因此常规产量大小应为BMC处理>CMC处理>CCC处理。
前茬种植作物为菜豆时,产量高低为BMB 处理>CMB 处理>BBB 处理,轮作处理产量均高于连作,这与丁素荣等研究得到玉米大豆轮作相对大豆连作增产的结果相同。与传统粮豆轮作会增加产量相同,CMB 处理放线菌门相对丰度变化较大且显著低于BMB、BBB 处理,这可能是第一年种植甘蓝导致土壤酸度增加引起,这也是CMB 处理产量低于BMB 处理的原因。
前茬种植作物为玉米时,产量高低为BBM 处理>BCM 处理>CCM 处理>CBM 处理,这与前人研究大豆玉米轮作产量显著高于玉米连作产量,且大豆−玉米产量高于大豆−大豆−玉米的结果不同。BBM 处理产量最高,这与每茬菜豆种植前均施适量肥料避免第二茬菜豆吸收第一茬菜豆残余养分导致土壤养分降低相关;CBM 处理产量最低,与土壤细菌大部分属缺失且7种属含量较高有关,也可能与CBM处理使土壤pH 提高较多,超过玉米适宜生长范围,进而影响土壤营养状况相关,这在6 种土壤细菌门相对丰度变化上有所体现,CBM 处理酸杆菌门、放线菌门相对丰度最低,其次为绿弯菌门、拟杆菌门、厚壁菌门相对丰度最高。酸杆菌门适宜生活在酸性环境,放线菌门适宜生活在中性偏碱环境,厚壁菌门在一定范围内随土壤pH 增大而增加。前人研究表明,土壤pH 是造成细菌群落结构变化的最大影响因子。
4 结论
(1)10 组处理共有OTUs 为2 299,分别归属于36门、97 纲、199 目、318 科、588 属、653 种,不同处理间土壤细菌OTUs无较大差异。
(2)BCM 处理细菌丰富度指数值最低,显著区别于CCC 处理;前茬种植玉米可降低细菌多样性指数值,其中CBM处理多样性指数最低。
(3)各处理优势菌门主要为变形菌门、酸杆菌门、拟杆菌门、厚壁菌门、绿弯菌门、放线菌门,CBM 处理土壤细菌相对丰度和群落结构对比连作变化最大,显著降低酸杆菌门、绿弯菌门、放线菌门相对丰度,显著提高拟杆菌门相对丰度;在属水平上CBM 处理与其余9种处理细菌群落差异最大。
(4)前茬种植作物相同时,各处理产量无显著差异。
(5)10 组处理间对比无特有OTU,前茬种植作物相同的处理间对比存在特有OTU。
(6)菜豆−菜豆−玉米对作物产量和土壤富营养优势细菌综合影响最好,因此推荐该轮作组合。但本试验只针对轮作第3 年作物进行了分析,还需进一步从土壤及作物其他角度进行比较,以明确轮作带来的效果和效果所能持续的年限。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
中国农学通报(2022年14期)2022-06-01
干旱地区农业研究(2022年1期)2022-01-28
动漫界·幼教365(中班)(2020年8期)2020-06-29
湖北农业科学(2019年22期)2019-12-23
幼儿教育·父母孩子版(2018年8期)2018-12-28
饮食科学(2016年9期)2016-11-18
故事大王(2014年3期)2014-05-21
探索财富(2013年10期)2013-11-18
作文大王·中高年级(2008年5期)2008-07-31
