
冷胁迫方式对太平洋牡蛎无水保活期氧化应激及能量消耗的影响
2022-03-31 09:04林恒宗高加龙梁志源范秀萍林海生曹文红黄艳平秦小明
广东海洋大学学报 2022年2期
林恒宗,高加龙,梁志源,范秀萍,林海生,曹文红,黄艳平,秦小明
冷胁迫方式对太平洋牡蛎无水保活期氧化应激及能量消耗的影响
林恒宗1,2,高加龙1,2,梁志源1,范秀萍1,2,林海生1,2,曹文红1,2,黄艳平1,秦小明1,2
(1.广东海洋大学食品科技学院,广东 湛江 524025;2.国家贝类加工技术研发分中心(湛江)// 广东省水产品加工与安全重点实验室 // 广东省海洋食品工程技术研发中心 // 广东省海洋生物制品工程重点实验室 // 水产品深加工广东普通高校重点实验室,广东 湛江 524088)
【目的】系统分析太平洋牡蛎() 保活运输前的预冷方式及温度胁迫对其氧化免疫系统、能量物质代谢的影响,筛选出较好的无水保活前预冷方式。【方法】分别采用散冰自然降温、急性连续降温、线性匀速降温、梯度降温4种低温诱导休眠方式处理牡蛎,并探讨不同诱导休眠方式对牡蛎生态冰温无水保活期的存活率、失重率、抗氧化能力以及能量物质代谢的影响。【结果】1)梯度和线性降温牡蛎冰温保活9 d其存活率及失重率优于散冰和急性降温组。2)各组牡蛎在整个保活期内抗氧化酶活力均升高,且散冰降温组显著高于急性、线性和梯度降温组(< 0.05);散冰和急性降温组丙二醛(MDA) 含量呈明显上升趋势,而线性和梯度降温组先降后升高;各不同降温组过氧化氢(H2O2) 含量则先升后降;总抗氧化能力(T-AOC) 随胁迫时间延长呈上升趋势。3)散冰降温组在整个保活期内能量物质均显著低于各组(< 0.05),保活9 d后线性和梯度降温组水分、糖原、脂肪含量高于各组,乳酸含量则低于各组。【结论】采用线性和梯度降温方式诱导牡蛎进入生态冰温休眠状态后开始无水保活,有利于减缓牡蛎在冷胁迫下抗氧化酶活力及脂质过氧化的影响,且保活时间更长,失重率更低,能量代谢物质损失较少,更有利于牡蛎长时间无水保活。
太平洋牡蛎;休眠方式;生态冰温;无水保活;氧化应激;能量代谢
太平洋牡蛎 () 是我国产量最高的牡蛎品种,主要分布在辽宁、山东等长江以北地区。牡蛎不仅味道鲜美且富含蛋白质、糖原、氨基酸、微量元素及维生素等营养物质[1-2],深受消费者青睐。牡蛎在我国主要以鲜销为主,但当前产业中牡蛎无水保活流通作业时缺乏系统的降温预冷工艺技术,鲜活牡蛎采捕后大多采用泡沫箱加冰强迫其进入休眠状态,后进入保活流通工序。然而,大幅降温会导致机体产生大量活性氧 (Reactive Oxygen Species,ROS),ROS会对细胞造成损伤、组织正常的生理机能和免疫防御能力产生不可逆的损害,从而影响其存活质量[3]。在虾夷扇贝 ()[4]、皱纹盘鲍 (HannaiIno)[5]、溢蛏 ()[6]、岩扇贝 ()[7]、魁蚶 ()[8]等贝类的研究中都已证实温度急剧变化对抗氧化系统及生理代谢产生直接影响。因此,合理的降温方式对延长贝类保活时间和保持贝类鲜度品质非常重要。目前关于太平洋牡蛎保活运输前的预冷方式及温度胁迫对其氧化免疫系统、能量物质代谢的研究鲜有报道。因此,本研究模拟当前国内太平洋牡蛎流通模式,将新鲜捕捞的牡蛎运抵实验室后暂养净化,采用不同降温方式对净化后的牡蛎进行诱导休眠处理,探索降温方式对牡蛎生态冰温保活过程中氧化应激及能量物质的影响,以期为双壳贝类保活前处理工序、低温保活运输条件优化提供技术和理论参考。
1 材料与方法
1.1 材料
鲜活太平洋牡蛎 (),2021年3月购自山东威海灯塔水母海洋科技有限公司,采捕后挑选富有活力、双壳完整、规格相近的带泥个体作为研究对象,迅速分装于套有内膜保湿袋的泡沫箱中,装箱前及装箱完成后在其表面与底面平铺一层约3 ~ 5 cm海水冰,且用吸水海绵与贝体隔开,经专用牡蛎冷链运输车运输48 h后运抵广东海洋大学水产品保活流通实验室暂养净化,以消除捕捞和运输操作时应激反应。牡蛎的平均生物学指标见表1。

表1 太平洋牡蛎生物学指标
1.2 主要仪器设备与试剂
贝类暂养净化系统,广州创岭水产有限公司;GXG-1低温恒温层析柜,江苏盛蓝仪器有限公司; JK-8多点温度计,常州金艾联公司;Varioskan Flash全自动酶标仪,美国Thermo公司;VAP450全自动凯氏定氮仪,德国Gerhardt公司;SZF-06A粗脂肪测定仪,上海洪记仪器设备有限公司;HX204卤素水分测定仪,瑞士梅特勒公司。
超氧化物歧化酶 (SOD)、过氧化氢酶 (CAT)、过氧化物酶 (POD)、谷胱甘肽过氧化物酶 (GSH-PX)、丙二醛 (MDA)、过氧化氢 (H2O2)、总抗氧化能力 (T-AOC)、糖原 (Gn)、乳酸 (LD)、蛋白质含量检测试剂盒均购自南京建成生物工程研究所。其余化学试剂为分析纯。
1.3 设计与方法
1.3.1 人工海水配制 在贝类净化系统中按太平洋牡蛎养殖海域盐度 (31±0.5)[9],用海水晶配制人工海水,开启增氧设备持续曝气24 h,通臭氧至海水臭氧质量浓度为0.15 mg/L,以流速2.5 m3/h通过紫外灯管进行灭菌,设定水温25 ℃循环预冷备用。
1.3.2 牡蛎暂养净化 采用灭菌人工海水清洗牡蛎表面附着物,将清洗后的牡蛎放入塑料筐 (600 mm × 420 mm × 315 mm) 置于贝类净化系统中,参照慕翠敏等[10]和费星等[11]的人净化工艺条件进行净化:水温25 ℃,贝水质量(kg)体积(L)比1:20,臭氧质量浓度0.15 mg/L,于盐度31的紫外循环灭菌人工海水中净化24 h,塑料筐与槽底距离2.5 cm以上防止二次污染。
1.3.3 牡蛎生态冰温的确定 临界温度测定参照储建军等[12]方法。挑选净化后富有活力的牡蛎20只放入装有人工海水的透明玻璃容器内 (320 mm × 150 mm × 212 mm) 内,通过调节低温恒温设备及加入人工海水冰块进行缓慢降温,同时观察箱内牡蛎的存活状态和对外界刺激的反应情况。牡蛎双壳自然张开,剌激反应微弱甚至无反应作为临界温度点。
结冰点测定参照徐德峰等[13]实验方法。挑选净化后富有活力的牡蛎10只,将多点温度计温度探针插入牡蛎肌肉内部,置于-20 ℃低温冰箱中,每5 s记录肌肉内部的温度,直到个体完全冻结。以时间为横坐标,温度为纵坐标绘制冻结曲线,根据冻结曲线拐点确定结冰点。由上述临界温度和冻结点测定结果及生态冰温定义确定太平洋牡蛎生态冰温范围。
1.3.4 牡蛎诱导休眠方法 将净化后挑选的600只太平洋牡蛎随机分为4组,每组150只,参照郝爽等[7]降温方法并加以改进,分别采用4种降温诱导休眠方式。其中散冰降温处理方式为:将牡蛎分装于泡沫箱中(450 mm × 250 mm × 200 mm),约5 kg/箱,装箱前后在其四周及表层平铺淡水冰,加盖后放置在0 ℃的恒温层析柜中开始无水保活。另外三种降温方式则是将牡蛎分装于保活塑料筐中,塑料筐的四周及表面均覆盖一层浸足灭菌海水的海绵,为保证环境湿度,每12 h向海绵喷淋灭菌海水。急性降温的处理方式为直接将牡蛎置于0 ℃的层析柜中,开始无水保活;线性及梯度降温速率参照胡益鸣等[14]的实验结果,通过调节恒温层析柜,以5 ℃/h的降温速率从(25±0.5)℃降至(0±0.5)℃后开始无水保活,其中梯度降温组牡蛎温度每下降5℃停留90min。各实验组牡蛎在诱导休眠结束后开始无水保活工序,保活周期为9 d,分别在降温前(BC)、降温后(AC)及保活期每天监测牡蛎存活及失重情况,另取腮和肌肉组织检测相关生化指标。
1.4 样品采集、制备与指标测定
1.4.1 存活及失重率监测 检活方法参照高加龙等[15]实验方法,每24 h观测牡蛎的贝壳张开情况,能自由闭合为活贝,不能闭合为死贝,依据存活及死亡数量绘制存活曲线。失重率监测参照申淑琦等[16]实验方法,选取个体差异较小、规格相近、富有活力的牡蛎20只,每天记录总质量。失重率为保活前后牡蛎质量损失与初始质量之比。
1.4.2 抗氧化酶活力测定 每组随机取9只牡蛎,迅速开壳取鳃,用预冷质量分数为0.86%生理盐水漂洗,吸水纸拭干表面水分,按质量(g)体积(mL)比1:9的比例加入预冷生理盐水进行冰浴机械匀浆,将制备好的体积分数为10%的匀浆液在4 ℃下以12 000 r/min的转速离心10 min,取上清于洁净EP管中,立即测定或-80 ℃冻存待测。严格按照试剂盒操作规程检测腮组织中蛋白质含量、SOD、CAT、POD、GSH-PX、MDA、H2O2、T-AOC酶活力,结果以蛋白浓度计算。
1.4.3 能量代谢指标测定 每组随机取15只牡蛎,迅速开壳刨取牡蛎全脏器,用预冷的质量分数为0.86%生理盐水漂洗,吸水纸拭干表面水分,采用高速匀浆机搅碎成肉糜状,分装,立即测定或-80 ℃冻存待测。严格按照试剂盒操作规程检测肌肉组织中Gn、LD含量。水分、粗蛋白、粗脂肪含量分别按照国标GB 5009.3-2016、GB 5009.5-2016、GB 5009.6-2016测定。
1.5 数据分析
实验结果均平行测定三次,数据采用平均值±标准偏差表示,对非正态分布数据存活率采用平方根反正弦变换后进行统计分析,对试验结果采用统计分析软件SPSS 26.0进行单因素方差分析 (One-Way ANOVA) 及组间差异采用Duncan多重比较,显著性水平设为0.05。
2 结果与分析
2.1 太平洋牡蛎生态冰温零点测定
太平洋牡蛎在缓慢降温过程中的行为特征如表2所示。在5 ℃以上太平洋牡蛎生理活动正常;当温度降至5 ℃时,应对外界刺激反应缓慢;3 ℃时基本无滤水表征现象,置于室温条件下能缓慢闭壳,表明太平洋牡蛎此时处于半休眠状态;0 ℃时对外界刺激已无任何反应,但置于常温人工海水能恢复正常滤水现象;-1℃时对外界刺激无反应,壳边缘出现微冻现象,温度继续降低成活率下降。判定牡蛎生态冰温零点约为0 ℃。

表2 不同温度下太平洋牡蛎活动状态
2.2 太平洋牡蛎结冰点测定
只有确定太平洋牡蛎耐受低温极限温度,才能通过控制适当低温使牡蛎处于休眠状态且不至于温度过低导致死亡。由图1可见,当温度降至-1.7 ℃时,牡蛎体内释放出大量潜热,此阶段牡蛎肌肉降温速度缓慢,甚至温度维持不变,因此得出太平洋牡蛎结冰点为-1.7 ℃,温度继续下降将导致肌肉结冰而引起牡蛎死亡。结合牡蛎生态冰温零点,太平洋牡蛎在无水保活过程中温度范围控制在-1.7 ~ 0 ℃。根据研究结果,机体越接近冰点温度易造成冷胁迫损伤,因此,在生态冰温保活过程中温度应尽量远离冰点温度。
2.3 冷胁迫方式对太平洋牡蛎无水保活期生命体征的影响
2.3.1 存活分析 不同降温方式对太平洋牡蛎无水保活过程存活曲线变化如图2所示。在冷胁迫下太平洋牡蛎存活率随保活时间延长均呈明显右移趋势。散冰自然降温组在保活1 d开始出现死贝,急性连续降温组在保活2 d开始出现死贝,梯度和线性匀速降温组在保活3 d出现死贝;对比4种不同降温方式,梯度降温组存活率最高,散冰自然降温组最低,保活6 d全部死亡;梯度、线性、急性降温组保活9 d后存活率分别为90%、85%、79%。可见,降温预冷方式对双壳贝类无水保活过程中存活率有较大影响。同时说明胁迫强度越大会加速太平洋牡蛎生理代谢, 降低机体对环境的适应能力,引起死亡,并随着时间延长而加剧,本实验保活条件下太平洋牡蛎的存活率与香港牡蛎 () 保活结果接近(保活9 d,存活率95%)[15]。
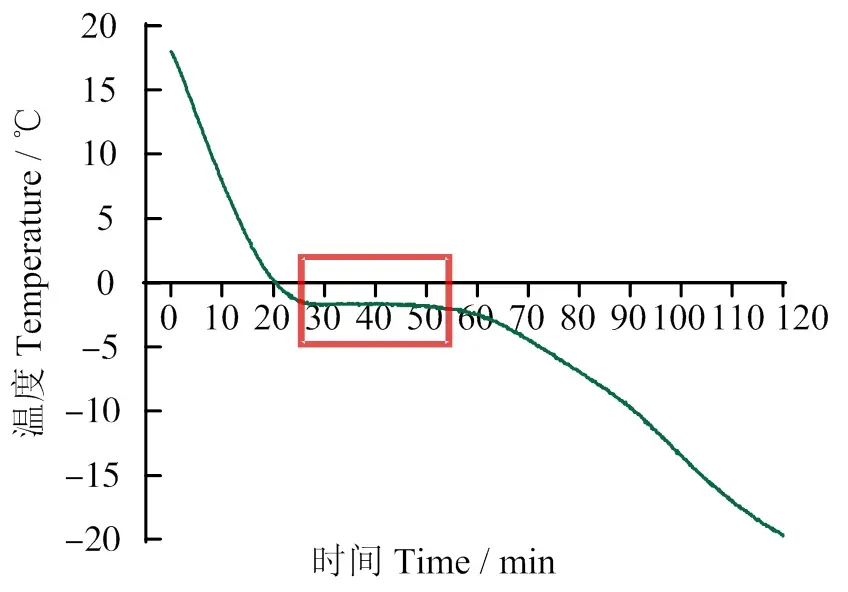
方框指示太平洋牡蛎冻结温度点
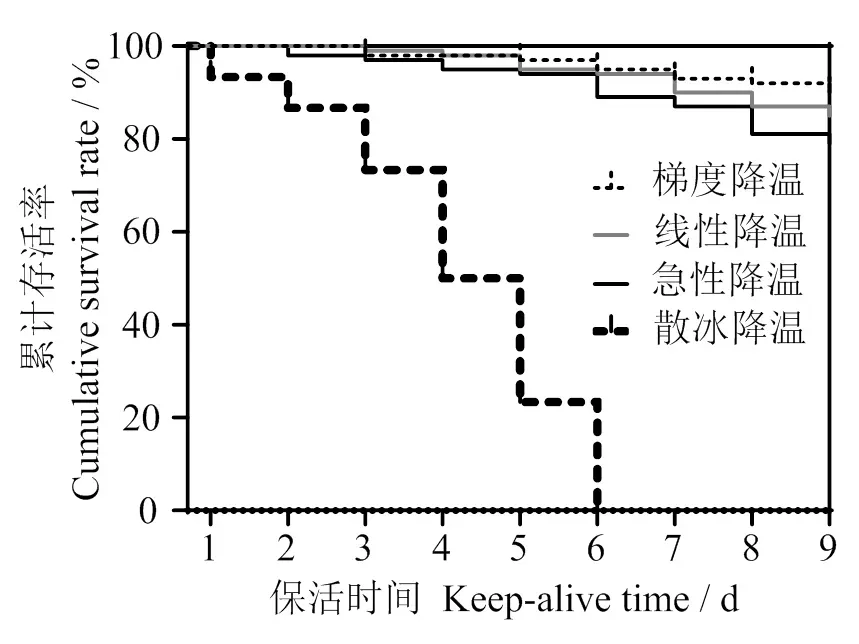
图2 不同冷胁迫方式下太平洋牡蛎无水保活期存活变化
2.3.2 失重率 不同降温方式对太平洋牡蛎无水保活期失重率的影响如图3所示,随着保活时间延长所有不同降温处理组牡蛎失重率均呈明显上升趋势,且不同降温方式对质量失重率的影响有一定差异。对比4种不同降温方式,散冰自然降温组在整个保活期内质量损失率明显低于其他3组,主要是由散冰融化,牡蛎长时间浸泡在冰水中所致。从保活2 d开始,梯度和线性降温组失重率明显低于急性连续降温组;保活5 d时失重率分别为11.98%和11.39%,而急性降温组保活5 d时高达15.78%;保活9 d后急性、线性和梯度降温组失重率分别为20.46%、16.82%、14.18%。结合存活率分析,可见采用线性和梯度的降温方式对牡蛎无水保活过程质量损失影响最小。
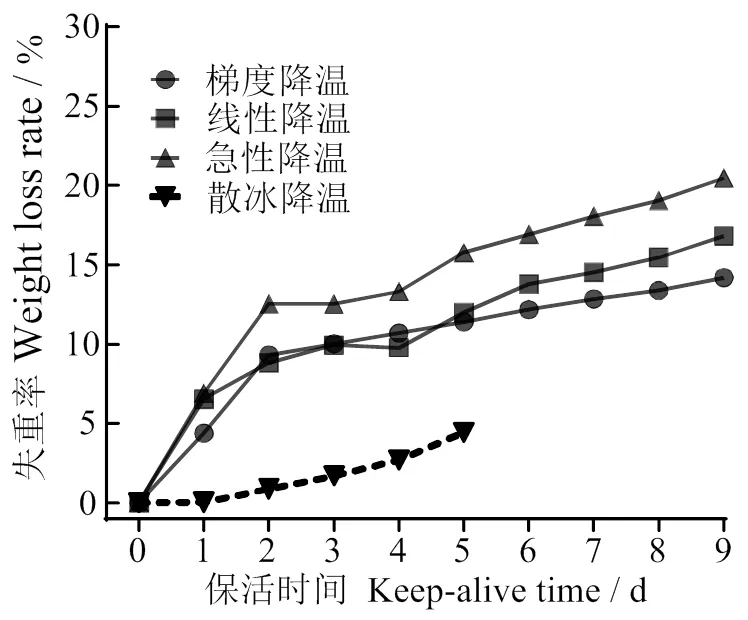
图3 不同冷胁迫方式对太平洋牡蛎无水保活期失重率的影响
2.4 冷胁迫方式对太平洋牡蛎无水保活期抗氧化酶活力的影响
图4(A-B)可知,在不同降温方式胁迫下,太平洋牡蛎腮组织中的SOD、CAT活力均随保活时间的延长呈上升趋势。降温后各组SOD、CAT活力较降温前明显升高,散冰自然降温组在保活前3 d均显著高于其他降温组 (< 0.05),在保活终期呈下降趋势;急性降温组SOD、CAT活力随保活时间延长呈直线上升趋势,且在保活7 ~ 9 d CAT活性显著高于线性和梯度降温组 (< 0.05);但线性和梯度降温组在保活0 ~ 3 d,SOD、CAT活力与降温后相比无明显增加 (> 0.05),随后呈缓慢增长趋势,除保活3 d外,线性和梯度降温组SOD、CAT活力在降温后保活1 ~ 9 d无明显差异(> 0.05)。表明,散冰与急性连续降温属于温度突变的降温方式,易导致牡蛎长时间处于应激状态,机体无法通过自身调节修复,氧化应激损伤明显高于线性和梯度降温组。温度作为主要的环境因子,对水生生物生理生化反应产生直接影响。在常温状态下,ROS处于动态平衡,但贝类在无水保活过程中受低温缺氧等因素胁迫,体内黄嘌呤脱氢酶可转化为黄嘌呤氧化酶,线粒体电子传递链载体失活,产生大量ROS,但贝类自身具备一套完整的抗氧化体系来维持机体ROS平衡[17-18]。由郝爽等[6]研究表明急性低温胁迫下溢蛏组织中SOD、CAT活力随处理时间延长呈不断升高趋势。陈丽梅等[19]研究发现,温度突变胁迫后毛蚶 () 组织中SOD、CAT活力明显升高。刘玲等[20]发现,渐变温度胁迫下对杂交石斑鱼(Valenciennes) 幼鱼抗氧化能力影响较小,这些研究结果均提示不良环境胁迫会诱导机体SOD、CAT活力升高,与本研究结果一致。
GSH-PX与POD它们在维持自由基平衡和减少应激损伤方面发挥着重要作用,在应激状态下GSH-PX将H2O2和有机过氧化物转化为H2O,而POD则具有降解消除H2O2和酚类、胺类、醛类、苯类毒性的双重作用[21]。由图4(C-D)可知,太平洋牡蛎腮组织中的GSH-PX与POD活力均随保活时间的延长呈上升趋势,除保活3 d外,散冰降温组牡蛎GSH-PX活性与急性降温组在降温后、保活1d、保活5 d无显著差异 (> 0.05),散冰自然降温组和急性降温组在降温后保活1 ~ 9 d均显著高于线性和梯度降温组 (< 0.05);线性和梯度降温组在降温后保活1 ~ 9 d POD活力变化不明显 (> 0.05)。可见GSH-PX、POD与SOD、CAT在清除自由基抵抗氧化损伤过程中发挥着相互协同及补充的作用,同时也表明在强应激条件下,牡蛎表现出更强的生理应激响应,线性和梯度渐变降温方式更有利于维持牡蛎抗氧化系统平衡。这一变化规律与李晓英等[22]对温度渐变下青蛤 () 肝胰腺组织中抗氧化酶活力的研究结果相似。
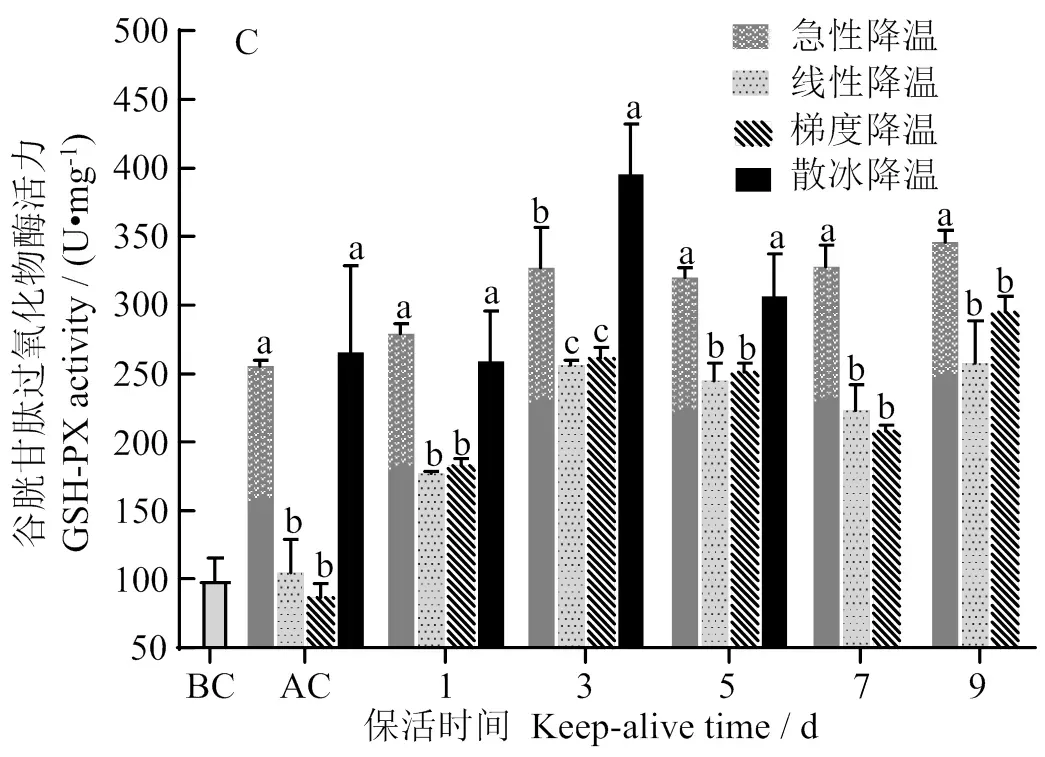
同一时间下,凡含一个相同字母表示组间无显著差异 (> 0.05)
At the same time, the data with a same letter means no significant difference between them(>0.05)
图4 不同冷胁迫方式对太平洋牡蛎无水保活期SOD(A)、CAT(B)、GSH-PX(C)、POD(D)的影响
Fig.4 Effects of differentcold stress methods on the SOD(A), CAT(B), GSH-PX(C), POD(D) ofduring the waterless keep alive
MDA含量代表脂质过氧化程度,间接反映组织细胞受自由基攻击的程度。一般来说,MDA含量越高,生物体所受的压力越大[23]。由图5(A)可见,太平洋牡蛎腮组织中的MDA含量均随保活时间的延长呈上升趋势且在降温后保活1 ~ 5 d散冰自然降温组MDA含量明显高于其他降温组 (< 0.05),急性和线性降温组在降温后MDA含量明显高于梯度降温组 (< 0.05),线性和梯度降温组在保活前期无明显变化,急性降温组MDA质量摩尔浓度在保活9 d达到最高值(13.1 nmol/g),显著高于线性和梯度降温组 (< 0.05)。表明,短期冰温胁迫下牡蛎抗氧化系统被激活,加速清除过多MDA以维持体内抗氧化系统平衡,但长时间冰温胁迫易导致牡蛎受到脂质过氧化损伤且强应激条件下牡蛎表现出更强的生理应激响应,散冰和急性降温组尤为明显。大量研究表明,机体在应激胁迫条件下MDA含量急剧上升,如岩牡蛎()[14]在急性温度胁迫下腮组织中MDA活力显著上升,华贵栉孔扇贝 ()[24]在遭受低温胁迫下MDA含量有所升高,以上结果均与本实验研究变化趋势相似。
需氧生物在氧化还原反应中产生大量的ROS,SOD将机体内产生的ROS分解成H2O2,而CAT、POD、GSH-PX又将H2O2还原成氧分子 (O2) 和水 (H2O),从而消除ROS对机体的影响[4]。由图5(B)可见,各不同降温组H2O2含量随保活时间延长总体呈先上升后下降趋势。在降温后保活1 ~ 9 d,散冰自然降温组H2O2含量均显著高于各降温组 (< 0.05);急性降温组H2O2含量在保活5、7、9 d显著高于线性和梯度降温组(< 0.05);线性和梯度降温组H2O2含量在降温后保活1 ~ 9 d无明显差异 (> 0.05)。结表明,散冰和急性降温组在受到直接冰温胁迫后通过SOD产生大量H2O2未被及时消解,且随着时间延长氧化应激逐渐加深,而线性和梯度降温组属温度渐变方式,机体H2O2能被SOD、CAT、POD、GSH-PX协同及时有效清除,有效阻止氧化应激的进一步损伤。本结果与郝爽等[25]报道的急性低温胁迫对溢蛏H2O2的变化趋势相似。
T-AOC是反映机体抗氧化能力的综合指标,它可以反映机体抗氧化防御系统应对外来胁迫的能力[26]。由图5(C)可见,散冰降温组在降温后保活1 ~ 9 d,T-AOC水平显著高于其他3组 (< 0.05),且在保活终期呈下降趋势;提示在长时强应激胁迫下由于牡蛎活力不足,致使体内总抗样化能力下降。除保活1 d外,急性降温组在降温后保活3 ~ 9 d内T-AOC含量均高于梯度降温组 (< 0.05),线性和梯度降温组无明显差异 (> 0.05),且梯度降温组在保活期内始终维持在降温前水平。
整体来看,T-AOC水平的变化情况与SOD和CAT活力变化呈现高度的一致性,同时也表明,散冰和急性冰温胁迫导致牡蛎ROS快速富集,抗氧化体系做出应答,表现为各种抗氧化酶活力以及总抗氧化能力显著升高,但线性和梯度缓慢降温方式在短时间内能通过自身调节来维持体内抗氧化系统平衡,长时间冰温胁迫会对机体造成一定的氧化应激损伤。曹善茂等[27]研究表明,岩牡蛎在低温胁迫下对其T-AOC能力有明显抑制作用。Gu等[28]研究表明,珍珠贝 () 在遭受干露胁迫时,总抗氧化能力呈先上升后下降趋势,以上结果均与本研究变化趋势相似。
同一时间下,凡含一个相同字母表示组间无显著差异 (P > 0.05)
At the same time, the data with a same letter means no significant difference between them(P>0.05)
图5 不同冷胁迫方式对太平洋牡蛎无水保活期MDA(A)、H2O2(B)和T-AOC(C)的影响
Fig.5 Effects of different cold stress methods on the MDA(A), H2O2(B) and T-AOC(C) of Crassostrea gigas during the waterless keep alive
2.5 冷胁迫方式对太平洋牡蛎无水保活期能量代谢的影响
不同低温诱导休眠方式对太平洋牡蛎主要能量物质变化见表3。可以看出,散冰自然降温组牡蛎受到冰温胁迫后脂肪和肌糖原变化明显 (< 0.05);急性降温组受到冰温胁迫后肌肉中水分、蛋白质、脂肪均无显著变化 (> 0.05),肌糖原含量则明显下降 (< 0.05);随着保活时间延长,散冰降温组在保活终期能量物质均显著低于其他降温组 (< 0.05);保活9 d后急性、线性和梯度降温组能量物质呈明显下降,乳酸则上升,能量代谢次序为糖原、粗脂肪、粗蛋白。急性降温组肌肉中水分、脂肪、糖原、乳酸含量与梯度降温组相比具有显著性差异 (< 0.05),但蛋白质则无明显差异 (> 0.05);线性和梯度降温组在保活9 d后脂肪、蛋白质、水分均无明显差异 (> 0.05)。结果说明,机体在受到强应激条件下主要消耗体内储存糖原来维持机体正常生理代谢,且胁迫强度越大,消耗糖原越多,但短期胁迫并不会启动脂肪、蛋白质来维持机体稳定。Bonga等[29]研究发现,机体一旦处于胁迫状态,肌糖原就会被优先调动起来用于供能。随着胁迫时间延长,机体受到低温、饥饿等多重胁迫后加速了水分的损耗,分解大量的糖原、脂肪来维持机体平衡,但对蛋白质利用较少,且胁迫强度越大,体内能量物质消耗的越快,这一现象在虾夷扇贝[30]、香港牡蛎[15]、菲律宾蛤仔()[31]无水流通过程中也有报道。同时也说明,线性和梯度的降温方式更有利于牡蛎在长时间保活中能量物质的维持。
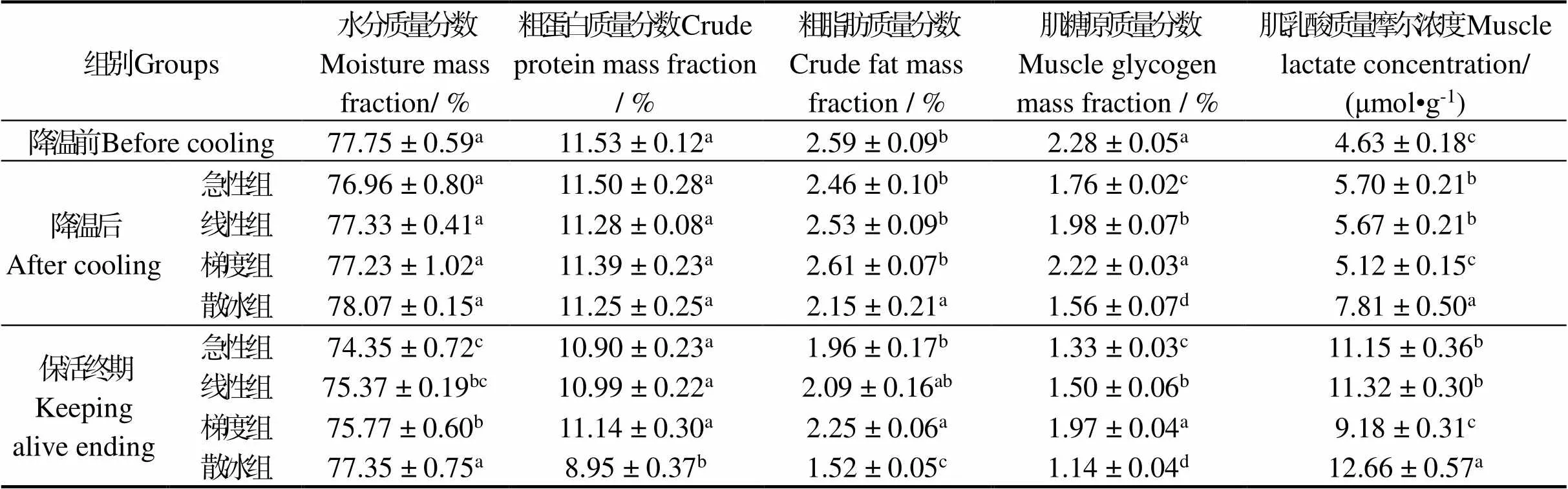
表3 冷胁迫方式对太平洋牡蛎无水期能量代谢的影响
注:相同指标下,不同处理方式间凡含一个相同字母表示差异不显著(>0.05)
Note: At the same index,the data with a same letter in different control groups means no significant difference between them(>0.05)
3 结论
散冰和急性降温胁迫对牡蛎抗氧化酶活力及脂质过氧化状态影响较大,加速了牡蛎抗氧化系统损伤,且随着应激时间的延长对其生命体征及能量代谢均产生显著影响。线性和梯度这两种较为缓和的降温方式能缓解牡蛎在冰温保活过程中氧化应激状态,短时间内有利于牡蛎抗氧化系统动态平衡的修复,且对其生命特征及能量代谢影响更小。综上,为减少运输损失和运输胁迫对牡蛎的健康造成影响,建议在采用线性和梯度的降温方式诱导牡蛎进入生态冰温状态后开始无水保活;同时可采用抗氧化指标(SOD、CAT、GSH-PX、POD、MDA) 作为贝类无水保活过程中的监控指标。
[1] LIU S, LI L, WANG W, et al.Characterization, fluctuation and tissue differences in nutrient content in the Pacific oyster () in Qingdao, Northern China[J].Aquaculture Research, 2020, 51(4): 1353-1364.[2] FUJITA K, SAITO M, VONGVICHITH B, et al.Analysis of the nutritional composition of aquatic species toward nutritional improvement in a Lao PDR rural area[J].Japan Agricultural Research Quarterly: JARQ, 2019, 53(3): 191-199.
[3] FAN X P, QIN X M, ZHANG C H, et al.Metabolic and anti-oxidative stress responses to low temperatures during the waterless preservation of the hybrid grouper (♀ ×♂)[J].Aquaculture, 2019, 508: 10-18.
[4] JIANG W W, LI J Q, GAO Y P, et al.Effects of temperature change on physiological and biochemical responses of Yesso scallop,[J].Aquaculture, 2016, 451: 463-472.
[5] 姜娓娓, 方建光, 李加琦, 等.温度胁迫对皱纹盘鲍生理和生化活动的影响[J].中国水产科学, 2017, 24(2): 220-230.
[6] 郝爽, 张敏.不同冷胁迫方式对缢蛏无水保活期抗氧化酶活性及脂质过氧化的影响[J].大连海洋大学学报, 2020, 35(4): 584-590.
[7] 王潇, 曹善茂, 刘钢, 等.温度对岩扇贝无水保活及抗氧化酶活力的影响[J].Marine Science Bulletin, 2018, 20(1): 1-20.
[8] XUE S Y, WANG J Y, LI J Q, et al.Effects of temperature on energy metabolism and antioxidant enzyme activities of[J].Journal of Fisheries of China, 2019, 43(3): 573-583.
[9] 山东省市场监督管理局.地理标志产品乳山牡蛎: DB37/T 3928—2020[S].北京: 中国标准出版社, 2020.
[10] 慕翠敏, 孔令峰, 于红, 等.太平洋牡蛎中大肠杆菌净化的实验研究[J].中国海洋大学学报(自然科学版), 2012, 42(S1): 83-86.
[11] 费星, 秦小明, 林华娟, 等.臭氧净化对近江牡蛎的存活率和主要营养成分的影响[J].食品工业科技, 2009, 30(7): 91-93.
[12] 储建军.冰温保活条件下缢蛏优势腐败菌生长动力学及耐冷机制研究[D].杭州: 浙江工商大学, 2017.
[13] 徐德峰, 王雅玲, 孙力军, 等.降温及生态冰温条件对凡纳滨对虾无水存活时间的影响[J].广东海洋大学学报, 2019, 39(6): 101-107.
[14] 胡益鸣, 李琪, 刘士凯, 等.温度和盐度急性胁迫对岩牡蛎存活及免疫指标的影响[J].中国水产科学, 2020, 27(3): 286-294.
[15] 高加龙, 章超桦, 秦小明, 等.不同温度无水保活对香港牡蛎微生物和基本营养成分的影响[J].广东海洋大学学报, 2020, 40(5): 90-96.
[16] 申淑琦, 万玉美, 申亮, 等.温度、湿度和氧气对海湾扇贝无水保活的影响[J].大连海洋大学学报, 2014, 29(5): 492-497.
[17] ZHANG G S, MAO J Q, LIANG F F, et al.Modulated expression and enzymatic activities of Darkbarbel catfish,for oxidative stress induced by acute hypoxia and reoxygenation[J].Chemosphere, 2016, 151: 271-279.
[18] LEVEELAHTI L, RYTKÖNEN K T, RENSHAW G M C, et al.Revisiting redox-active antioxidant defenses in response to hypoxic challenge in both hypoxia-tolerant and hypoxia-sensitive fish species[J].Fish Physiology and Biochemistry, 2014, 40(1): 183-191.
[19] 陈丽梅, 刘利华, 胡宏辉, 等.温度突变对毛蚶不同组织抗氧化酶活性的影响[J].水产科学, 2019, 38(4): 435-442.
[20] 刘玲, 陈超, 李炎璐, 等.短期温度胁迫对驼背鲈(♀)×鞍带石斑鱼(♂)杂交子代幼鱼抗氧化及消化酶活性的影响[J].渔业科学进展, 2018, 39(2): 59-66.
[21] DUAN Y F, ZHANG J S, DONG H B, et al.Oxidative stress response of the black tiger shrimptochallenge[J].Fish & Shellfish Immunology, 2015, 46(2): 354-365.
[22] 李晓英, 董志国, 阎斌伦, 等.青蛤POD组织差异及温度骤升和窒息胁迫对青蛤POD的影响[J].海洋通报, 2010, 29(5): 521-525.
[23] VÍCTOR V M, ESPULGUES J V, HERNÁNDEZ- MIJARES A, et al.Oxidative stress and mitochondrial dysfunction in: a potential therapy with mitochondria-targeted antioxidants[J].Infectious Disorders Drug Targets, 2009, 9(4): 376-389.
[24] 杜可, 林岗, 李惠莲, 等.低温胁迫对三沙湾华贵栉孔扇贝生理指标的影响[J].福建师范大学学报(自然科学版), 2021, 37(6): 62-69.
[25] 郝爽, 张敏, 方佳琪.缢蛏在急性温度胁迫下的氧化应激响应及生理代谢变化[J].上海海洋大学学报, 2021, 30(2): 214-221.
[26] GIESEG S P, CUDDIHY S, HILL J V, et al.A comparison of plasma vitamin C and E levels in two Antarctic and two temperate water fish species[J].Comparative Biochemistry and Physiology Part B, Biochemistry & Molecular Biology, 2000, 125(3): 371-378.
[27] 曹善茂, 王潇, 刘钢, 等.温度胁迫对岩扇贝幼贝抗氧化酶活力的影响[J].大连海洋大学学报, 2018, 33(2): 223-227.
[28] GU Z F, WEI H J, CHENG F, et al.Effects of air exposure time and temperature on physiological energetics and oxidative stress of winged pearl oyster ()[J].Aquaculture Reports, 2020, 17: 100384.
[29] WENDELAAR B S E.The stress response in fish[J].Physiological Reviews, 1997, 77(3): 591-625.
[30] 陈文秀, 欧阳杰, 徐文其, 等.贮藏条件对活品虾夷扇贝营养及品质的影响[J].渔业现代化, 2019, 46(6): 83-89.
[31] BI S J, XUE C H, SUN C, et al.Impact of transportation and rehydration strategies on the physiological responses of clams ()[J].Aquaculture Reports, 2022, 22: 100976.
Effect of Cold Stress Methods on Antioxidant and Energy Metabolism ofin Water-free Live Storage Period
LIN Heng-zong1,2, GAO Jia-long1,2, LIANG Zhi-yuan1, FAN Xiu-ping1,2, LIN Hai-sheng1,2, CAO Wen-hong1,2, HUANG Yan-ping1, QIN Xiao-ming1,2
(1.,,524025,2.()////////,524088,)
【Objective】To systematically analyze the effects of pre-cooling and temperature stress on the oxidative immune system and energy metabolism ofbefore live transport, and select a better pre-cooling methods before waterless keep alive.【Method】Oysters were either treated by natural cooling, acute continuous cooling, linear uniform cooling and gradient cooling in crush ice.The survival rate, weight loss rate, antioxidant capacity, energy metabolism of the oyster were determined to evaluate the best method of dormancy induction.【Results】1) the survival rate and weight loss rate of gradient and linear cooling methods were better than the natural cooling and acute cooling.2) The antioxidant enzyme activity of each group increased during the whole live storage, ice cooling group was significantly higher than acute, linear and gradient cooling groups (< 0.05); MDA content in natural cooling and acute cooling group were increased significantly.In contrast, the MDA value showed an initial decreased then increased trend in the linear and gradient cooling groups; each groups.The H2O2level of all test groups showed an initial increased and then decreased pattern while the T-AOC was at an upward trend when the stress time was extended.3) The energy content of ice cooling group was significantly lower than other groups during the test period (< 0.05).The moisture, glycogen and fat content of the linear and gradient cooling groups were higher than the others groups, but lactic acid was lower than them.【Conclusion】When theunder ice are induced dormancy and are kept waterless after treatment by linear and gradient cooling methods (cooling rate ≤ 5 ℃/h), the antioxidant enzyme activity and lipid peroxidation will be reduced.The survival time is extended, the weight loss rate and the loss of energy metabolites is lower for the cold stress group, which is more conductive to the waterless keep alive retention of oysters.
; hibernation methods; ecological ice temperature; waterless keep alive; oxidative stress; energy metabolism
Q939.11
A
1673-9159(2022)02-0095-09
10.3969/j.issn.1673-9159.2022.02.012
2021-12-02
“十三五”国家重点研发计划“蓝色粮仓科技创新”重点专项(2019YFD0901601)
林恒宗(1996―),男,硕士研究生,研究方向为贝类净化及保活运输技术。E-mail:1074680995@qq.com
秦小明(1964―),男,教授,研究方向为贝类净化和水产品保活运输技术。E-mail: xiaoming0502@21cn.scom
林恒宗,高加龙,梁志源,等.冷胁迫方式对太平洋牡蛎无水保活期氧化应激及能量消耗的影响[J].广东海洋大学学报,2022,42(2):95-103.
(责任编辑:刘朏)
猜你喜欢
九江学院学报(自然科学版)(2022年2期)2022-07-02
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
福建基础教育研究(2019年3期)2019-05-28
西部资源(2018年1期)2018-11-01
卷宗(2018年14期)2018-06-29
中学生数理化(高中版.高一使用)(2018年2期)2018-04-04
小资CHIC!ELEGANCE(2018年8期)2018-04-03
环球时报(2017-12-25)2017-12-25
数学大王·低年级(2015年10期)2015-10-21
山海经(2015年18期)2015-04-19
