
为什么和声能诱发音乐情感?——音乐协和性的作用及其认知机制
2022-04-13 01:52张积家
心理科学进展 2022年4期
张 航 孟 乐 张积家
为什么和声能诱发音乐情感?——音乐协和性的作用及其认知机制
张 航1孟 乐1张积家2
(1中国人民大学心理学系, 北京 110872) (2广西师范大学教育学部, 桂林 541004)
音乐是高级意识活动产生的声音艺术, 对人类的情感表达和交流具有重要意义。作为连接音乐与情绪的核心要素, 协和性的形成原理至今仍然未有定论。人类如何加工多个音符构成的和声?为什么一些和声听起来协和(愉悦), 一些和声听起来不协和(不愉悦)?协和感究竟是自下而上的声学感知还是自上而下的审美体验?从古希腊时代至今, 这些问题就一直吸引着学者的目光。物理学家从协和与不协和的声学区别中寻找答案, 生理学家从听觉生理机制方面分析协和感的产生, 心理学家研究协和音程偏好是与生俱来的还是后天形成的。目前, 音乐协和性的理论内容主要以西方音乐为主, 中国传统民族音乐迫切需要开展相关的实证研究。
音乐协和性, 和谐性, 拍频, 音乐文化, 认知机制
1 引言
音乐是经由人类意识活动产生的声音组合的高级艺术, 只有对声音的频率和节奏等信息进行有意识编排才能够创造出优美的旋律。因此, 创作、演奏或聆听音乐是人类具有复杂声音事件处理能力的重要标志。作为人类的特有才能, 音乐活动贯穿人类文明史的始终。考古学证据显示, 人类至少在35000年前的旧石器时代就能够制作并且使用乐器(Conard et al., 2009;Killin, 2018)。作为人类的通用语言, 音乐存在于所有的文明社会(有文字的或无文字的)中(Mehr et al., 2019)。作为情感交流的艺术载体, 音乐伴随着人的一生, 从摇篮曲到葬礼哀乐, 都是人类释放情感和引发共鸣的重要方式(Belfi et al., 2021)。音乐对人类的非凡意义使得许多学科都重视对音乐基础理论的研究。在创刊125周年之际, 就将音乐和语言的起源列为125个最具挑战性的科学问题之一(Kennedy & Norman, 2005)。也以“科学与音乐”为专题, 探讨音乐理论与实践中亟待解决的重要问题, 包括现代和声音乐学的理论核心——音乐的协和性(musical consonance) (Ball, 2008)。
协和性是音乐构建的基本要素。无论何种音乐形式或题材, 都由音符组成, 音符同时发声形成和声(harmony), 相继发声构成旋律(melody)。音乐协和性是两个或两个以上音符同时演奏时所引发的心理感受(McDermott et al., 2016; Parncutt & Hair, 2011; Zentner & Kagan, 1996)。根据演奏效果, 可以将和声分为两类, 听起来愉悦、稳定或彼此融合的和声称作协和音程(consonant interval)或协和和弦(consonant chord), 听起来不愉悦、不稳定或彼此不融合的和声称作不协和音程(dissonant interval)或不协和和弦(dissonant chord) (Harrison & Pearce, 2020; McDermott et al., 2016)。迄今为止, 已经有大量的理论试图说明音乐中存在协和与不协和两种音响效果的原因, 但都存在着较大的争议。这些理论可以归纳为三种(见表1):(1)强调音乐刺激的物理属性, 认为和声音程的声学特征是影响音乐协和性的主要因素, 主张总结出人类普遍遵循的数理规律; (2)侧重于考察音乐加工的生理属性和心理属性, 认为音乐协和感是声音事件在听觉通道诱发的情绪体验, 主张采用生理声学实验和心理声学实验揭示出这种普遍的听觉加工机制。这两种理论不约而同地认为, 音乐协和性的感知能力是人类与生俱来的天赋才能。(3)聚焦音乐的文化属性, 认为音乐协和性知觉是后天形成的审美经验, 受文化熏陶和音乐训练影响。本文以上述三种理论为脉胳, 介绍不同学科的成果, 阐明人类音乐协和感的形成原理和认知机制, 为未来的相关研究尤其是中国民族音乐协和性的理论研究提供启示。

表1 音乐协和性的相关理论
2 音乐协和性的数理之美
音乐是一门艺术, 但当人们研究其原理、分析其性质、发现其规则时, 就成为一门科学。音乐协和性理论的发展正是基于对音乐美学的不懈追求, 这从根本上推动了人类使用理性的目光审视其背后的奥秘, 探测隐藏于音符间微妙的数理关系。从数理角度探讨音乐协和性的理论主要包括简单整数比学说(Small-Integer Ratios Theory)和泛音和谐理论(Harmonicity Theory)。
2.1 简单整数比学说
音乐协和性的早期研究可以追溯至古希腊时期。毕达哥拉斯是第一个触及该领域的学者。他发现, 同时拨动的两条琴弦长度之比越简单, 和声效果越协和, 弦长之比越复杂, 和声效果越不协和(Crocker, 1963; Purves, 2017)。据此, 把弦长关系1: 2 (纯八度)、2: 3 (纯五度)和3: 4 (纯四度)的音响组合划定为协和一致的愉悦音程, 并从和谐哲学观念赋予这些数值以特殊象征含义——“圣十结构” (Tetractys, 10 = 1 + 2 + 3 + 4), 认为美的事物总是有类似简单且和谐的数学特征, 甚至将“数”视为万物之源, 宇宙和谐本质上是“数”的和谐(Bowling & Purves, 2015; Crocker, 1963; Primavesi, 2016)。简单整数比学说提出促进了音乐协和性的数理规律研究。尤其是在文艺复兴时期, 大量作曲家在简单整数比思想指导下探索协和性的规律, 以丰富协和音程与和弦的数量。意大利作曲家扎利诺用“六数列”来说明协和性的产生原则(6 = 1 + 2 + 3 = 1 × 2 × 3), 认为所有的协和音程都由1到6几个数字组成(Bowling & Purves, 2015; Palisca, 1961; Sukljan, 2020), 从而把原来一直被毕氏学派看作是不协和音程的6: 5 (小三度)、5: 4 (大三度)和5: 3 (大六度)调整至协和音程范畴, 为推动和声音程系统分类做出重要贡献(Tenney, 1988)。至此, 现代和声音乐学中协和音程和不协和音程基本确立。
音乐形式的鼎故革新和音乐种类的丰富多彩表明, 音乐极具有社会历史性和文化多样性。但是, 通过比较不同时期甚至是不同地区的研究发现, 人对音程协和性的感知偏好存在着极高的相似性(图1a), 且极度符合简单整数比学说, 听众普遍地将比值简单的音符组合评定为协和音程, 将比值复杂的音符组合评定为不协和音程。这说明, 数理思想对解释协和性仍然具有较强的现实意义。即便在今天, 没有一种理论能够像简单整数比说那样对音乐艺术产生如此大的影响。由此也引发一个关键性的疑问, 为什么比值简单的和声听起来协和?
2.2 泛音和谐理论
声音科学的诞生为解答和声协和性提供了新的思考方式。首先, 声音是一种机械振动, 振动频率决定音高, 所以, 影响音乐协和性的本质因素是弦振动频率比而非弦长比(Konoval, 2018)。其次, 泛音(overtones)作为最基本的声学现象是音乐协和性形成的物理基础(Pesic, 2014)。因为传统乐器或人发出的声音都是复合音, 除了能够听到一个响亮的基音音高外, 还能够听到一系列相对柔弱却悦耳协和的“弦外之音” (Dostrovsky, 1975; Konoval, 2018)。以理想的弦振动为例(图1b), 被拨动的琴弦同时做整段振动和局部振动(二分之一、三分之一、四分之一…), 整段振动产生清楚响亮的基音(fundamental tone), 分段振动产生了一系列整数倍于基音频率(简称基频, frequency fundamental)的泛音。泛音现象发现为数学和音乐学之间搭建了桥梁, 包括拉莫在内的许多音乐学家主张运用声学理论而非纯粹数学来研究音乐协和性。拉莫认为, 在自然泛音(natural harmonics)中包含许多协和音程, 构成协和音程的泛音列与基音距离更近, 听起来更清晰, 构成不协和音程的泛音列距离基音较远, 不容易被察觉(Christensen, 2004; Kieffer, 2016), 如表2所示, 离基音较近的几个泛音无论是与基音的音程关系还是与前一个泛音的音程关系都非常协和, 表明和谐的泛音列本身就是一个协和和弦(chord)。所以, 简单整数比的音乐审美倾向在一定程度上反映了人类对自然和谐泛音的知觉偏好, 当两个音被同时演奏时, 其叠加形成的声学特征在泛音关系上越和谐, 构成的声音织体就越协和。

图1 a. 从1898年至2020年间, 来自6个国家受试者对12个和声音程音乐协和性的等级排列结果高度一致。数据分别选自:美国(Guernsey, 1928), 日本(Butler & Daston, 1968), 德国(Stumpf, 1898), 英国(Pear, 1911), 新加坡(Bowling & Purves, 2015), 加拿大(Weiss et al., 2020)。b. 吉他琴弦振动产生的泛音列。c. 协和音程(纯五度)和不协和音程(三全音)的频谱示意图。绿色实线代表根音音符A的频率成分, 基频是110 Hz, 泛音是220, 330, 440, 550, 660 Hz等。三全音中红色虚线代表冠音音符Eb, 基频是156 Hz, 泛音是 312, 468, 624, 780, 936 Hz等。纯五度中红色虚线代表冠音音符E, 基频是165 Hz, 泛音列是330, 495, 660, 852 Hz等。黑色虚线代表纯五度的虚拟音高。d. 基底膜上拍频效应的示意图。拍频效应通常由泛音列过于接近的不协和音程引发。

表2 自然倍音列中基音与泛音之间的协和关系
注:C代表协和英文缩写“consonance”, D代表不协和英文缩写“dissonance”.
人类对和谐泛音的偏好可能是一种适应性的加工机制, 并且有重要的进化意义。因为人类的嗓音、大多数动物的叫声以及物体的有规则振动都产生与基音呈倍频关系的和谐泛音特征(嗓音称共振峰, formants), 这些声音也是社交、择偶或生存警示意义的凸显性信号(Bowling et al., 2017)。所以, 在自然选择压力下, 人类进化出了一套可以解析和谐泛音的听觉系统, 能够在纷繁无序的声音世界中迅速地分辨泛音有序的重要信息。研究发现, 嗓音作为生物学意义极高的听觉刺激是有效地预测音乐协和性的指标, 那些与嗓音频谱特征相似的和弦听起来更协和(Bowling et al., 2018; Gill & Purves, 2009), 说明相较于泛音无序的噪音, 人类大脑更容易处理嗓音般的和谐声响进而引发积极的情绪体验。嗓音被喻为人类的“听觉面孔”, 闻声数秒, 就能够迅速地获取对方的性别、身份、情绪、吸引力甚至是体型大小等信息(Pisanski & Feinberg, 2019)。对嗓音相似特征的加工优势(processing advantage)是人脑的预装机制(Bowling & Purves, 2015; Harrison & Pearce, 2020)。不仅如此, Schwartz等人(2003)对上千段语音材料进行特征提取后发现, 人类言语信息的振幅−频率分布模式恰好对应于音程协和度的等级顺序, 表明人类在言语特征上的统计知觉经验训练了人耳对特定频率比的音程协和感。和谐泛音偏好也可能发轫于人类对于大自然和谐音响法则的崇尚与追求。例如, 大三和弦产生就是源于对自然泛音的观察(Christensen, 2004), 西方五度相生律和中国三分损益律的建立也受自然泛音的启发, 甚至最初的音乐形式就是人类对大自然声音律动的倾听和模仿(王小盾, 2017)。《淮南子》中有:“乐生于音, 音生于律, 律生于风, 此声之宗也。”大自然的风吟雨落、鸟叫虫鸣极可能是人类形成和谐音响审美法则的最初灵感来源。
但是, 持信息加工观点的研究者却认为, 泛音作为物理特征所以引发协和性的情绪体验主要与音高的表征机制有关(McLachlan et al., 2013)。音高是乐音(物体有规则振动产生)的基本听觉属性, 对音高的感知是理解音乐和语言的重要媒介(Norman-Haignere et al., 2019)。通常说来, 音高由基音决定, 但多声源复合信号的音高知觉就变得复杂并受泛音信息影响(Harrison & Pearce, 2020; Oxenham, 2018; Popham et al., 2018)。呈倍频关系的基音和泛音被自动地知觉为来自同一声源, 基音决定音高, 泛音影响音色, 但不具和谐关系的声音由于无法被听觉系统有效地分辨, 会造成知觉不流畅的不愉悦体验。研究证实, 听者对协和音程与不协和音程的音高知觉水平存在着差异, 大脑对不协和音程的音高编码比协和音程慢36ms (Tabas et al., 2019)。在不协和音程中完成音高匹配的错误率更高, 反应时更长, 说明大脑并不擅长处理泛音不和谐的声音(Seror & Neill, 2015)。甚至在不协和音乐背景中进行Stroop任务时, 由不一致条件引起的冲突效应增加, 表明不和谐泛音会占用大量的认知资源并且影响听者的执行功能(Bonin & Smilek, 2016; Masataka & Perlovsky, 2013; Seror & Neill, 2015)。而且, 模式匹配理论(Pattern Matching Theory)和时域自相关理论(Temporal Autocorrelation Theory)还强调, 泛音和谐性影响音高表征的准确度进而引发音乐协和性的知觉体验(de Cheveigné, 2005; Harrison & Pearce, 2020; McDermott & Oxenham, 2008)。模式匹配理论认为, 长时记忆储存和谐泛音的频谱特征, 如果听觉信息与之吻合, 即便基音缺失了(fundamental missing), 大脑也能够自动地利用泛音与基音的倍频关系补全并感知到音高(Feng & Wang, 2017; Oxenham, 2018)。所以, 和谐泛音的音高知觉起来清晰明确, 不和谐泛音在匹配中会出现多个“虚拟音高” (virtual pitch), 从而造成音高不可辨的杂乱音响(Milne et al., 2016; Parncutt & Hair, 2011; Stolzenburg, 2015)。如图1c所示, 协和音程P5(纯五度)的泛音列和谐度较高, 因而有明确的音高值(F0等于原来两个泛音列的最大公约数), 不协和音程Tritone (三全音)的泛音列错落不均, 无法被听觉系统有效地处理, 所以引起不愉悦的体验。因此, 音高知觉流畅性是引起音乐协和感的重要原因。时域自相关理论认为, 听觉系统根据声波在时域上的周期性变化来表征泛音的和谐性, 并且以求最小正周期的方式推测音高值。比值复杂的不协和音程由于自相关系数低导致泛音列的周期性差。例如, 小二度产生的几个虚拟音高值甚至低于人类的乐音感知阈限(30 Hz) (McDermott & Oxenham, 2008)。实际上, 两种理论争论的焦点在于听觉系统计算音高的方式, 根据傅里叶变换原理, 声波在时域上的周期性等价于在频域上的和谐性, 但两种理论在音高知觉流畅性是引发音乐协和性的观点上并无分歧。
总的来说, 17世纪的科学革命促进了物理声学发展, 在实证主义思想推动下, 研究者开始将目光从数理哲学转向声学, 发现泛音与音乐协和性的联系, 逐渐形成了泛音和谐性理论。
3 音乐协和性的感知之美
声学的物理特征是导致声音协和与否的外在条件, 人耳如何感知声学线索才是引发音乐协和性的关键。协和性归根结底是人类的听觉体验。所以, 到19世纪, 部分学者开始研究音乐引起的生理−心理反应机制, 而非只关注物理现象。赫尔姆霍兹就是这一领域的先驱者, 他提出的拍频理论(Beat Theory)目前仍然是现代音乐学教材中普遍推崇的观点。
3.1 拍频理论
在音乐知觉测量中, 赫尔姆霍兹同样意识到泛音特征的重要, 但并未像拉莫等人一样把音程协和性看作是泛音列的和谐自然, 而是转向研究泛音之间的相互作用及对听觉系统的影响(Bowling & Purves, 2015)。他认为, “自然泛音之和谐只能称叹宇宙法则之神奇, 再无它用, 因为自然界中美与丑的存在概率相同, 事物的自然性并不足以证明它是精美的。相反, 一些音程之所以不协和是因为听起来杂乱刺耳, 其原理是两个频率相近的声波相遇时会发生干涉效应, 出现振幅涨落、强弱不均的拍频现象, 由此引起基底膜上听觉感受器的间歇性兴奋从而导致不舒服之感” (Helmholtz, 1885)。这种令人不愉悦的体验可以定义为粗糙感(roughness)。粗糙感影响音程协和性。和声听起来协和是因为粗糙感不明显, 听起来不协和是因为粗糙感明显(如图1d所示, 在不协和音程三全音中, 频率相近的泛音列在基底膜相应位置引发了拍频效应)。所以, 音程使用应该遵循简单整数比原则, 防止泛音过于紧凑引起明显的粗糙感。
拍频理论提出后, 大量的生理声学和心理声学的实验结果都相继支持了这一假设。贝凯西的耳蜗振动研究对推动拍频生理机制探索具有里程碑的意义(von Békésy, 1960)。基于听觉位置理论, 贝凯西发现, 耳蜗基底膜以行波的形式振动, 不同的声波频率分别对应于基底膜不同位置的响应。这项工作不仅令他获得了1961年诺贝尔生理与医学奖, 更让大量的心理声学数据可以直接与基底膜振动图谱数据比较(Bowling & Purves, 2015)。在比较中, 研究者发现, 耳蜗就像是一个天然的分频滤波器, 上面布满了一系列相互交叠的负责响应特定频率范围的听觉感受器, 当两个声波频率相近时, 激励的感受器范围有很大一部分重叠, 这部分重叠区域就会受到双音拍频效应影响引起粗糙感(Greenwood, 1961; Vassilakis, 2005; Schneider 2018)。因此, 研究者将出现粗糙感的频率差值范围称为临界频带(critical bands), 如果两个声波的频率刚好落在相同临界频带内就会引起粗糙感, 落在临界频带外就不引起粗糙感(Greenwood, 1991; Plomp & Levelt, 1965; Vassilakis, 2005)。例如, 同时呈现两个频率值为1,2 (1>2)的纯音(正弦波), 其叠加形成的信号可表示为:
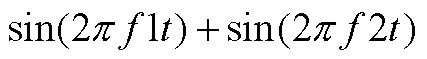
根据和差化积公式, 上式也可变为:
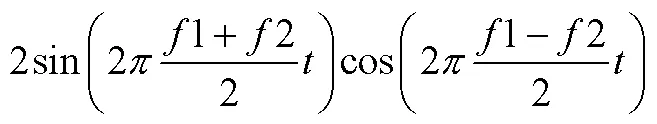
当双音之间的频率差值足够大时, 听者知觉到的是公式(1), 此时仍然是两个清晰可辨的频率为1、2的声音信号; 但当双音只有较小的频率差值时, 听者所知觉到的是公式(2), 这时叠加信号以频率为(1+2)/ 2, 振幅涨落为Δ=1−2的方式振动。其中, 较慢的振幅涨落(Δ= 0.1~5 Hz)在响度上不会被知觉为不愉悦, 甚至听起来像一个融合音(fusion), 但快速振荡的振幅涨落(Δ= 20~ 250 Hz)会引起忽强忽弱的刺耳抖动(Harrison & Pearce, 2020; Shapira & Stone, 2008)。所以, 临界频带范围实际就是双音可辨——粗糙感知觉——双音完全融合的变化范围。Plomp和Levelt (1965)计算, 纯音音程的最大协和度发生在临界宽带值的125%, 极度不协和音程发生在临界宽带值的25%。后续的研究也引入了新算法估计临界频带对复合音协和度的影响(Bernini & Talamucci, 2014; Vencovskỳ, 2016)。拍频理论的建立使得音乐审美感受和人耳内部生理构造及活动规律进行了有效的结合。与泛音和谐论的观点类似, 拍频理论也强调音乐协和性建立在一般感知水平上并且受听觉加工系统的约束。
3.2 拍频理论面临的挑战
虽然拍频理论是上世纪的主流观点, 但是, 随着研究的深入, 拍频理论受到了前所未有的挑战。首先是该理论的外部效度, 因为拍频值受发生干涉效应的声音频率成分的振幅和相位影响, 这些因素在不同乐器之间差异很大(Kohlrausch et al., 2000; Plack, 2010), 所以拍频不是衡量音乐协和性的可靠线索。其次, 拍频也不是引发协和感的关键原因。采用双耳分听(dichotic)方式给被试呈现音乐刺激时, 拍频效应导致的粗糙感被显著降低了(两个声音分别作用于双侧耳蜗从而避免了干涉效应), 却未影响音程协和性的评价, 说明拍频对音乐协和性的作用很小(Bidelman & Krishnan, 2009; McDermott et al., 2010)。更为有趣的是, 按照拍频理论, 构成和弦的音符越多, 理应造成更明显的粗糙感, 但三音(triads)甚至四音(tetrads)和弦高于双音(dyads)协和性的情况大量存在(Bowling & Purves, 2015)。听者对纯音(不含泛音)、只含少数泛音的复合音和含有较多泛音的复合音在协和度评价上没有显著差异, 说明泛音的有无及数量都不影响协和性知觉(McLachlan et al., 2013), 这显然与拍频理论的泛音干涉效应的假设相违。
尽管许多研究者从不同的方面指出了拍频理论的局限性, 但都未能够实质性地否认拍频对音乐协和性的作用, 因为在通常的情况下, 不协和音程同时包含了拍频较明显和泛音不和谐两种特征, 因此很难准确地计算某一种因素的贡献, 也就很难拒绝某一种理论假设。McDermont等人(2010)不但分离了两种因素的相互影响, 还直接计算了每种因素与音乐协和性的相关。该研究分析了三个重要的指标:(1)对音乐协和性的偏好, 以被试对协和音程与不协和音程的喜好程度差异来衡量; (2)对拍频的厌恶, 以在双耳分听与双耳同听条件下对纯音喜好程度差异来衡量; (3)对泛音和谐性的偏好, 以对和谐泛音与不和谐泛音(人为进行泛音扰动处理)喜好程度差异来衡量。结果表明, 尽管被试并不喜欢拍频明显的和声, 但对拍频的厌恶与对音乐协和性的偏好却没有相关性, 对和谐泛音的偏好显著预测了对音乐协和性的偏好, 充分说明影响音乐协和感的关键性因素并不是拍频的有无。另外, 对先天性失音症(congenital amusia)研究表明, 失音症病人不能像正常被试一样区分音程协和性(Cousineau et al., 2012; Zhou et al., 2019), 但对听觉粗糙感的知觉评价却与正常被试无异, 二者都喜欢拍频不明显的刺激而厌恶拍频明显的刺激, 说明失音症的音乐协和性加工缺陷与拍频因素无关(Cousineau et al., 2012)。总的来说, 拍频理论一直强调没有粗糙感是导致音程听起来协和的原因, 却忽略了协和音程本身可能具有的美学机理, 正如Bowling和Purves (2015)所评论的, 不苦并不是糖吃起来甜的原因。
4 生物与文化的融合之美
音乐是声音组合的艺术。不同国家和民族都能以独特的形式自由灵活地创作音乐, 进而形成了风格迥异、百家齐放的音乐文化。作为音乐审美的核心要素, 音乐协和性在多大程度上受生物学因素影响有很大的争论。许多研究者认为, 协和性偏好是由于人类听觉系统对协和音程的声学物理特征存在加工优势, 这是镶嵌在人类认知系统的先天适应性机制(Bidelman, 2013; Bowling et al., 2017; Virtala & Tervaniemi, 2017)。但是, 民族音乐学家(ethnomusicologist)和作曲家反对这一观点, 认为音乐协和性是后天习得的审美经验, 存在较大的文化差异和个体差异(Brown & Jordania, 2013; McDermott et al., 2016; Popescu et al., 2019)。由此, 引发了音乐协和性起源的先天与后天之争。
4.1 生物决定论
持生物决定论观点的学者认为, 音乐协和性知觉是人类与生俱来的天赋才能。首先, 发展心理学的证据表明, 19到40个月大的儿童对不同协和程度的音乐刺激表现出明显的偏好差异, 他们花费更长的时间探索那些发出协和音乐声响的玩具(di Stefano et al., 2017)。6个月(Crowder et al., 1991; Trainor & Heinmiller, 1998)、 4个月(Trainor et al., 2002; Zentner & Kagan, 1996)甚至2个月大的婴儿(Trainor et al., 2002)也都展现出音乐协和性的知觉反应能力和偏好行为。Zentner和Kagan (1996)采用转头偏好范式考察了4个月大婴儿的音乐协和性知觉能力, 发现在播放协和音乐时, 婴儿表现出更少的头部运动并且对声源刺激方向注视更长的时间。父母是聋哑人的婴儿对协和音程产生强烈的喜爱, 说明音乐知觉经验不是导致婴儿形成协和感的主要因素, 这就排除了教养或环境暴露等学习效应的影响(Masataka, 2006)。一些神经科学研究甚至发现, 新生儿就已经具有音乐协和感, 这无疑为音乐审美先天论的观点提供了强有力的证据(Perani et al., 2010; Virtala et al., 2013)。但是, 近期采用转头偏好范式的研究却未能够重复以往的实验结果, 只发现了熟悉性对协和性知觉的影响, 因此质疑早期使用类似范式的研究可能混淆了刺激材料的熟悉性和新异性(Plantinga & Trehub, 2014)。但不管怎样, 这些对儿童早期的实证研究至少说明音乐协和性知觉产生与人类的生物学基础存在着很大的关联。
不同文化对音阶(scale)的使用也佐证了这一猜测。人类在中等音阈范围内至少能够分辨240个音高, 在原则上应该有大量的音阶用来创作音乐, 但是, 无论是西方古典音乐、现代流行音乐、中国传统民乐及其它文化的音乐都只使用5到7个相似的音阶音级(Bowling et al., 2017; Gill & Purves, 2009), 并且不约而同地构成了相似的和声音程(Bowling et al., 2017; Bowling & Purves, 2015), 甚至不同国家出土的早期乐器也都只含有这几个音阶, 修缮后竟然还能演奏出协和音响(Conard et al., 2009; Killin, 2018)。这充分说明, 先天的生物学因素约束了人类对音阶的选择, 也约束了和声音程的知觉加工。最后, 两种主流的音乐协和性理论都支持音乐审美先天论的假设。泛音和谐论认为, 人脑预装有自然泛音特征识别器, 加工与之吻合的有简单频率比的协和音程时会引起流畅的愉悦体验, 加工与之不吻合的有复杂声学特征的不协和音程会引起负性体验。拍频论认为, 听觉器官的生理构造是导致复杂不协和音程引起粗糙感的原因。所以, 二者都强调音乐协和性是先天听觉机制在处理特定声音刺激时引发的情感效应, 只不过前者认为该过程发生在大脑的神经系统, 后者认为该过程发生在耳蜗的基底膜。因此, 人类的音乐协和性知觉可能是一种先天的音乐才能。
4.2 文化决定论
民族音乐学家和作曲家把音乐协和性看作一种审美经验, 认为文化背景和后天音乐训练塑造了个体的音乐协和性知觉。首先, 和声音乐的审美观念在不同文化之间差异较大。例如, 巴厘岛甘美兰(Gamelan)音乐在演奏时产生非常明显的拍频效应, 这种引起粗糙感的音乐形式在当地十分盛行(Harrison & Pearce, 2020; Plantinga & Trehub, 2014)。在中东、印度北部、立陶宛、波斯尼亚以及巴布亚新几内亚等地区, 人们更是主动追求器乐乃至声乐间的拍频复调(beat diaphony), 说明拍频在这些文化中被视为令人愉悦的协和音乐元素(Ambrazevičius, 2017; Harrison, 2021; Vassilakis, 2005)。事实上, 拍频效应的文化差异被民族音乐学家广泛考证, 但许多研究者在撰写结论时往往忽略了这些因素(Plantinga & Trehub, 2014)。其次, 听者对音乐协和性的知觉评价也受接触程度影响。Meyer (1903)报告说, 被试初次接触微分音乐(microtonal music)时会感到很不协和, 重复播放了12到15遍后, 大多数参与者都自发地表示听起来还不错。Valentine (1914)指出, 当把一些不协和音程重复听33遍, 被试会认为它是协和的。不过, 这些早期的研究报告并未考虑纯粹接触效应(mere exposure)的影响(Montoya et al., 2017)。后来的研究发现, 仅仅是短暂的实验室训练就能够显著地提高被试对协和音程的偏好效应(McLachlan et al., 2013), 对受过专业训练的音乐家来说, 偏好效应更加显著(Brattico et al., 2009; Dellacherie et al., 2011; McDermott et al., 2010)。即便是未受过音乐教育的儿童, 随着年龄的增长, 对音乐协和性的知觉也会提高(Weiss et al., 2020)。这些来自跨文化、音乐训练和毕生发展的研究结果都向音乐协和性的生物本能论发起了挑战。
把这场争论推向高潮的是2016年刊登在杂志的封面文章(McDermott et al., 2016)。该文系统考察了协和音程偏好在多大程度上是受现代和声音乐文化影响。作者选取了玻利维亚热带雨林中仍然过着原始狩猎采摘生活的提斯曼人(Tsimane)和生活在城市和乡镇过着现代生活的玻利维亚人以及完全生活在西方和声音乐文化下的美国人(音乐家和非音乐家)。结果发现, 虽然提斯曼人在声音情感知觉和音乐粗糙度的分辨能力上与对照组相差不大, 但提斯曼人却给予协和与不协和音程以相同等级的愉悦度评价。相反, 生活在城市及乡镇的玻利维亚人却显著地偏好协和音程, 偏好程度依次低于美国的非音乐家和音乐家。这说明, 音乐协和感的形成与接触现代和声音乐文化的程度有关。如果一个地区完全没有被和声音乐影响, 这里的人们就可能就缺乏音乐协和性知觉。所以, 音乐协和性不是先天的知觉偏好反应, 文化因素在音乐审美方面起到了巨大作用。提斯曼人没有聆听或演奏和声音乐的经历, 他们的音乐属于仅有一条旋律线的单音音乐(Monophony), 是最简单的音乐形式, 西方和声音乐至少有两条以上的旋律线交织, 所以, 提斯曼人没有形成音乐协和感可能是缺乏对纵向多线程音乐的知觉经验(McDermott et al., 2016; McPherson et al., 2020)。知觉磁体效应(perceptual magnet effect)也能够说明这一现象。婴儿早期能够分辨大量的语音范畴, 但所处的语言环境如果缺少对某些声音的分辨要求(例如日语中的“r”和“l”), 在特定的年龄后对这些声音的分辨能力就会消失(Werker & Hensch, 2015), 表明人类的听觉系统会根据环境的需要进行调整。一些音乐加工模型也认为, 文化和经验通过塑造大脑的皮层奖赏环路影响音乐奖赏系统(Zatorre & Salimpoor, 2013)。所以, 音乐协和性的形成极可能是神经系统与文化长期相互作用的产物, 而不是单方面的因素所致。
4.3 比较心理学的介入
音乐协和性起源问题所以悬而未决, 一个重要原因是无法完全避免先天生物学因素和后天环境变量的相互影响, 因为以人类为被试的研究无法真正做到长期隔绝一切声音刺激。比较心理学研究动物行为, 能够提供有效控制后天环境接触的手段(Harrison & Pearce, 2020; Toro & Crespo- Bojorque, 2017)。在实验室条件下, 动物在出生前就被隔绝外部声音刺激, 所以, 动物表现出来的任何行为偏好都可以排除后天接触特定听觉信息(如自然泛音接触)的影响。动物不具有产生音乐的能力, 因此, 任何音乐偏好性反应都可以视作是生物普遍性的一般听觉加工, 而不是音乐特有的认知加工。因此, 比较心理学是考察音乐协和性究竟是先天性行为还是习得性行为的重要方法, 也是理解音乐初始状态以及这种初始状态如何被相关经验塑造的重要途径(Toro & Crespo-Bojorque, 2017)。
相关的研究主要集中在分类能力和偏好行为上(图2)。研究表明, 部分鸟类和灵长类动物经过训练以后能够准确地区分协和音程和不协和音程, 如爪哇禾雀(Watanabe et al., 2005)、黑顶山雀(Hoeschele et al., 2012)、欧洲椋鸟(Hulse et al., 1995)和日本猕猴(Izumi, 2000), 并且能够将这种分类能力迁移至新的音乐刺激。猴子的初级听觉皮层(Fishman et al., 2001)以及猫的听神经(Tramo et al., 2001)和下丘(McKinney et al., 2001)对协和与不协和音程的神经反应有显著差异, 说明一般性的听觉机制是导致某些动物掌握音乐协和性分类能力的重要原因。研究还发现, 一些动物幼崽能够立即表现出音乐协和性的偏好行为。例如, 刚孵化的小鸡会对播放有协和旋律的视觉印记表现出明显的接近行为(Chiandetti & Vallortigara, 2011)。黑猩猩幼崽也更喜欢通过连接在它胳膊上的装置播放协和音乐旋律, 提示人类可能与同在系统发育上接近的黑猩猩共享音乐协和性的进化起源(Sugimoto et al., 2010)。但是, 当使用孤立的和声音程为刺激时, 棉花顶面绢毛猴在V型迷宫中并未在协和音程附近逗留更的长时间(Koda et al., 2013)。与之相反, 人类的音乐协和性偏好不仅在完整旋律条件下显著, 也在孤立和声的情况下存在(McDermott et al., 2010; Toro & Crespo-Bojorque, 2017), 说明较少的音乐信息对比可能不足以引起某些非类人猿灵长类动物的音乐协和性知觉。
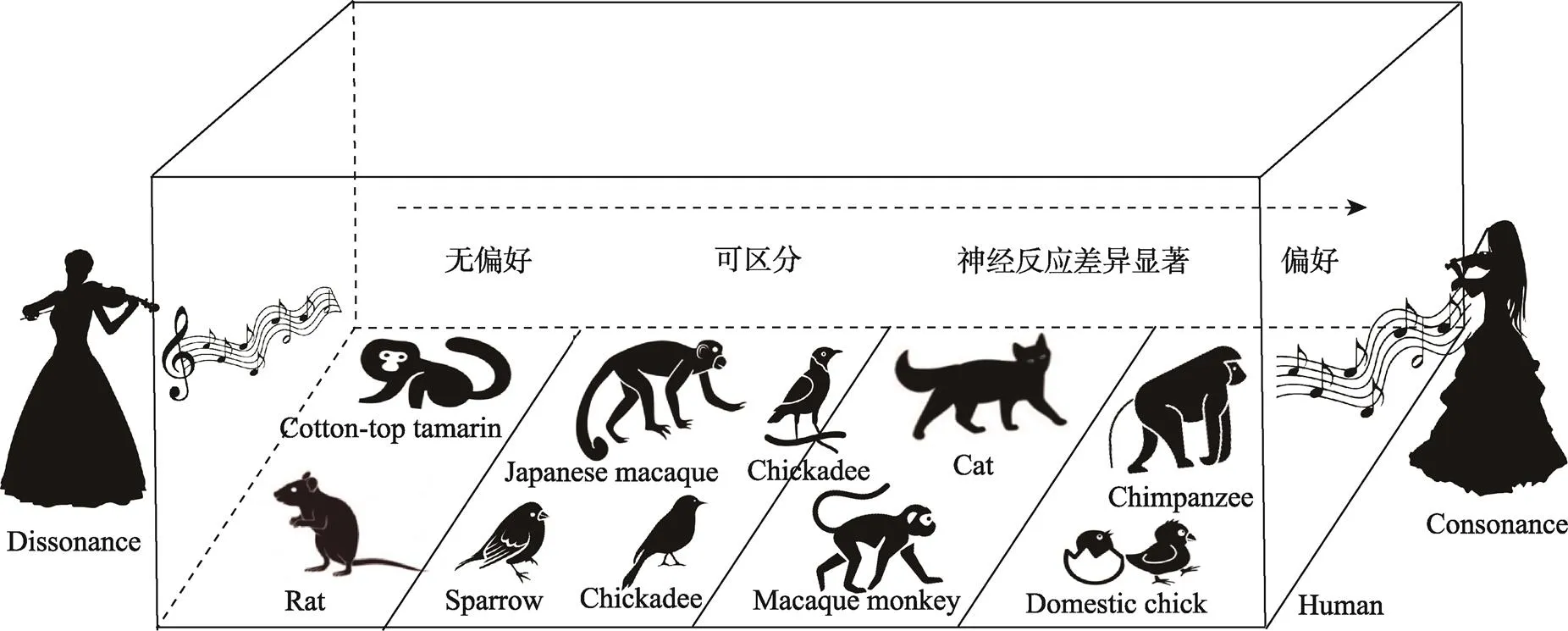
图2 不同物种对协和与不协和音乐的分辨能力和偏好反应的实验结果总结
为了排除动物自身叫声的影响, 研究者采用啮齿类动物如褐家鼠为研究对象(Crespo-Bojorque & Toro, 2015; Postal et al., 2020)。因为与大多数常发出和谐泛音叫声的动物不同, 褐家鼠的叫声一般不具有和谐泛音特征, 在多数情况下是纯音样的超声波, 这就为验证泛音和谐性的知觉接触理论控制了自身叫声这一混淆变量。研究发现, 即便褐家鼠在强化训练下习得了分辨协和音程与不协和音程的能力, 但不能泛化至其他新的音程刺激, 也不能迁移至倍频转换(octave transpositions)音程条件, 说明褐家鼠只是暂时记住了音程间的绝对频率差异。导致褐家鼠不具有音乐协和性知觉能力的原因可能是缺乏和声音乐或和谐泛音的接触经验(Crespo-Bojorque & Toro, 2015; Toro & Crespo-Bojorque, 2017)。未来研究可以让褐家鼠自出生前就暴露在这些声学线索中, 以考察接触经验是否对音乐协和性偏好有塑造作用。总之, 比较心理学是理解音乐知觉如何受后天文化环境熏陶的重要手段。而且, 音乐协和性也并非为人类所特有, 一些动物与人类在发音−听觉系统可能存在相似的进化起源。
4.4 多因素混合理论
为了弥合音乐协和性理论中的先天论与后天论之争, 一些研究者提出了整体性概念结构理论, 认为生物学因素与文化因素共同影响音乐协和性知觉。其中, 和谐泛音列是引起协和感的主要因素, 拍频效应是导致不协和感体验的主要原因, 经验和文化通过审美观念自上而下地调节音乐协和性知觉(Friedman et al., 2021; Parncutt & Hair, 2011; Parncutt et al., 2019)。Bowling等人(2017)认为, 忽视生物因素或环境信息的任何一方都无法准确说明音乐协和性的产生, 所以多因素混合理论是解释音乐协和性产生的最佳方式。最有说服力的证据来自Harrison和Pearce (2020)归纳的混合计算模型。他们重新评估了此前分别基于泛音和谐性、拍频效应以及文化特征三种因素理论的几种经典模型的预测力, 发现尽管依据不同理论建立的模型在预测力上存在着差异, 但三种因素都是衡量音乐协和性的有效指标。而且, 重新拟合一个同时包含这些因素的混合计算模型以后, 预测力提高至88.95%, 显著大于此前所有的单因素计算模型。作者还利用音乐语料库分析法, 考察了过去千年间的10000部古典、爵士和流行音乐作品的和弦流行性(chord prevalence)。结果发现, 泛音和谐性对和弦流行性的预测有显著的正相关, 拍频效应与和弦流行性呈显著的负相关, 说明一直以来作曲家都更青睐在创作中添加泛音和谐的音乐元素, 尽量避免使用那些可能引发听众不愉悦情绪的粗糙感元素, 再次证实音乐协和性同时受到拍频效应和泛音协和性影响。综上, 音乐协和性是由多种因素引起的音乐审美知觉, 未来研究应该试图分离这些因素的相互作用, 以确定加工这些信息是否存在共享的神经通路。
5 音乐协和性的认知神经机制
近20年来, 随着认知神经科学的发展, 借助神经电生理(如ERP)和脑成像(如fMRI)等技术, 研究者在解析音乐协和性的认知加工机制方面取得重要进展。大量的证据表明, 人脑对和声音程的加工遵循自下而上的加工模式, 与音乐协和性相关的音高信息在传入高级皮层之前就已经被初步处理, 这些初期感觉编码形式遵循高水平的音乐层级规则, 而非简单的声学物理特征, 说明音乐协和性的认知体验基于感觉水平的神经活动(Bidelman, 2013; Bidelman & Grall, 2014; Bidelman & Heinz, 2011; Bidelman & Krishnan, 2009; Bones et al., 2014; Itoh et al., 2010; Kim et al., 2017)。在早期注意阶段, 初级和次级听觉皮层负责音高信息的抽象表征(Bidelman & Grall, 2014; Itoh et al., 2010), 在更早期的前注意阶段(pre-attentive stages), 皮层下组织结构(如脑干和听神经)中神经元群的锁相活动是编码音高信息的认知通路起源(Bidelman, 2013; Bidelman & Heinz, 2011; Bidelman & Krishnan, 2009; Bones et al., 2014;Kim et al., 2017)。因此, 音乐协和性认知偏好在皮层和皮层下水平均具有生物学基础。Itoh等人(2010)利用ERP技术验证脑电信号反应与音乐协和性的等级顺序存在紧密关联, 该实验任务是向被试随机呈现跨度分布范围在0~13个半音的和声音程, 发现200~300 ms左右诱发的N2成分显著地受音程协和性高低调节, 不协和音程相较于协和音程诱发了波幅更大的N2, 构成和声音程的两个音符音高之比越简单, 诱发的N2波幅就越小, 极度符合简单频率比学说的预测, 说明大脑皮层的神经活动对音程中的音高关系非常敏感, 表征这些音程信息的方式与标准音乐实践的协和性分类原则一致。由音高线索引发的瞬态特异性神经信号——音高起始反应(pitch onset response)的波幅变化也与行为结果及音乐协和性的等级顺序一致, 再次证明音乐协和性知觉建立在低水平的神经反应上, 而且早期编码形式就已经采用了高水平的音乐组织原则(Bidelman & Grall, 2014)。言语加工过程的语音编码(phonetic code)模式也与之相似, 初级听觉皮层及其邻近区域对言语信息的早期加工同样遵循语音信息的高级抽象规则而非简单的声学特征(Scharinger et al., 2011), 提示音乐和语言可能共享特定的神经基础。
在皮层下水平, 研究者以频率跟随响应(frequency-following response, FFR)为指标发现音乐协和性的认知通路可能起源于脑干(Bidelman, 2013; Bidelman & Heinz, 2011; Bidelman & Krishnan, 2009; Bones et al., 2014)。FFR是周期性声音刺激诱发产生的听觉脑干稳态响应, 反映脑干神经元群对周期性声音刺激时间细节结构(temporal fine structure)的同步锁相活动, 因此, 利用FFR能够清晰地记录和比较脑干对不同协和程度音乐刺激的同步锁相能力差异(Bidelman, 2013; Bidelman & Krishnan, 2009; Bones et al., 2014; Coffey et al., 2016)。研究发现, 协和音程比不协和音程诱发更稳健且同步性水平更高的相位锁定信号(协和音程在基频和倍频处诱发的频谱能量幅度更大且更集中), 信号幅值与被试的评定结果及音乐协和性的等级程度显著相关, 说明脑干神经元群的锁相活动中储存了关于音乐协和性音高等级关系的感觉信息, 并以此作为最初的编码模式(Bidelman & Krishnan, 2009)。动物研究发现, 脑干(Tramo et al., 2001)和下丘(McKinney et al., 2001)等区域对协和音程表现出更稳健的同步锁相活动, 并且能够有效地预测动物的音程偏好行为。所以, 人类和动物的研究都表明, 协和音程在听觉系统中存在编码优势。这也说明, 相较于周期性不规则的不协和音程, 皮层下神经元群对协和音程声学特征的同步锁相能力更强, 这在很大程度上降低了大脑的计算负荷。换言之, 音乐协和性知觉偏好差异反映了听觉系统对音高层级信息表征能力的神经生物学差异。总之, 发生在皮层水平的更高级的音乐愉悦感体验建立在皮层下水平的神经反应基础上。
大脑的高级认知区域也参与音乐协和性的知觉加工。Minati等人(2009)表明, 协和和弦比不协和和弦在额叶中下回(inferior and middle frontal gyri)、前运动皮层 (premotor cortex)和顶下叶(inferior parietal lobule)等区域诱发更强的血氧动力学信号响应, 非音乐家被试的激活水平表现出明显的右侧化优势, 音乐家未表现出明显的偏侧化。但是, Foss等人(2007)的研究却发现相反的音乐协和性效应, 不协和音程比协和音程在非音乐家的右侧额下回以及音乐家的前扣带回(anterior cingulate cortex)、颞上回(superior temporal gyrus)、左侧额叶中下回和顶下叶等区域的激活水平更强。导致这种不一致的结果可能是实验任务和刺激材料不同, 前者采用四音和弦为刺激, 并要求被试判断每个和弦的协和性(Minati et al., 2009), 后者使用双音和声, 被试只需要被动地听(Foss et al., 2007)。此外, 在一项系统平衡了音符数量、音色、节奏及旋律轮廓的研究中, 不协和音程比协和音程在内侧前额叶(medial prefrontal cortex)和前扣带回喙部等区域的激活水平更高(Bravo et al., 2020)。有趣的是, 这些脑区是负责冲突检测及评估负性情绪和恐惧相关信息的脑区, 而且在不协和条件下, 双侧初级听觉皮层与扣带回之间的功能连接更强, 说明大脑在处理不协和音乐时需要更多的信息资源参与, 也证实了不协和音程具有明显的负性效价信息特征。与之相反, 协和音程引发的正性愉悦体验有极高的奖赏价值(Gold et al., 2019; Zatorre & Salimpoor, 2013)。一项采用预测奖赏错误(reward prediction errors)范式的研究发现, 以协和音程结尾的音乐令被试产生动机性学习行为, 并且像美食和金钱一样激活中脑多巴胺奖赏系统的核心——伏隔核(nucleus accumbens)(Gold et al., 2019)。因此, 大脑高级皮层对协和音程与不协和音程的加工可能存在着不同的认知通路。
以上的研究表明, 音乐协和性信息既能够诱发皮层下前注意水平的基础神经反应, 也能够调动高级水平的认知资源参与, 甚至能够激活奖赏系统和影响执行功能。两种认知水平的表征均受音乐训练影响(Andermann et al., 2020; Bidelman & Heinz, 2011; Foss et al., 2007; Itoh et al., 2010; Minati et al., 2009; Pagès-Portabella & Toro, 2020; Proverbio et al., 2016), 这说明, 音乐经验对音乐协和性的知觉加工具有塑造作用。未来研究应该探讨不同音乐文化背景如何自上而下的调节音乐协和性的神经机制。
6 研究展望
音乐具有传情达意的作用, 能够引起听者强烈的情绪体验。音乐协和性是连接音乐与情绪的关键元素。相关文献已经在理论观点和研究方法上有了较大的突破, 但仍然存在着许多尚需解决的问题:(1)什么样的声学特征导致和声音程被认为是协和的?(2)人类如何进化出加工这些特征的能力?(3)为什么人类把这些声学特征赋予了相应的审美属性?
针对问题一, 最值得关注的是泛音和谐性与拍频效应的理论之争。然而, 二者并非不可调和, 近期的研究结果都相继支持了多因素混合理论的假设, 认为二者共同决定音乐协和性知觉(Friedman et al., 2021; Harrison & Pearce, 2020)。但这也引发了诸多疑问。譬如, 两种认知活动对音乐协和性的影响哪一个作用更大?二者引起的协和性知觉效果是否截然相反, 即前者主要负责引发协和感, 后者主要导致不协和感?最初的信息编码形式是发生在神经系统还是发生在基底膜?未来研究应该寻找新的范式分离这两个因素。针对问题二, 语音相似性假设和比较心理学证据都提供了较好的回答, 对协和音程偏好反映了听觉系统对自然和谐泛音特征以及同类嗓音相似性特征的神经生物学倾向, 但对不协和音程排斥的生物学原因还并不清楚。此前的研究表明, 粗糙感极为明显的尖叫声音能够直击大脑的恐惧中枢——杏仁核(Arnal et al., 2015), 人类很可能把厌恶情绪泛化至所有具有明显粗糙感的声音特征。未来研究可以分析不协和音程在诱发负性情绪方面是否遵循相似的演化规律和认知模式。问题三要比前两个问题更加复杂, 因为音乐协和性知觉是否在人类社会中普遍存在尚有争议, 除了受先天因素约束外, 经验塑造及文化熏陶同样扮演着重要角色, 甚至有人认为, 人类根本就没有进化出任何音乐审美能力, 对音程的喜爱或排斥只是长期受西方现代和声音乐规范影响的结果。一些未接触过西方音乐文化的群体就未表现出协和性的审美偏好(McDermott et al., 2016), 一些地区甚至盛行不协和的拍频复调音乐(Ambrazevičius, 2017; Vassilakis, 2005)。所以, 人类为何将一些声学特征赋予相应的审美属性仍然值得进一步思考。此外, 中国传统民族音乐具有偏重旋律变化而轻视和声作用的特点, 在乐器样式、音乐形式和文化内涵等方面都与西方音乐存在较大差异。研究者可以基于音乐库分析法, 考察中国民族音乐在未接触西方音乐前具有怎样的和声知觉观念, 这或许有助于理解人类为何会形成当前的和声音乐审美特征。而且, 中国是一个民族众多的国家, 蕴含着大量多声部音乐形态的少数民族音乐文化, 如侗族大歌和壮族山歌等, 这些多声部音乐在音程协和性的表现手法上是否与西方和声音乐存在着异曲同工之妙?相应的, 民族文化又如何塑造这些群体的音乐审美特征?近期, 由意大利和尼泊尔政府联合开展的喜马拉雅山脉干城章嘉峰科考项目发现, 影响音乐协和性的关键因素是音乐文化熏陶, 与海拔高度、人口特征及民族身份无关(Prete et al., 2020)。因此, 深入探究一些音乐文化较为浓厚的多声部少数民族音乐, 有利于阐明文化对音乐协和性审美观念的塑造作用。
国内对音乐协和性的研究主要集中在史实描述、作品分析和审美思辨等方面, 具有较强的主观性和个人风格, 几乎很难见到实证性的文章, 主要原因与国内音乐学与其它学科的交流度不高有关。相较之下, 国外对音乐协和性的研究成果颇丰富, 涉及声学分析、毕生发展、音乐训练、文化经验以及动物行为等多方面的实证性考察, 并结合认知神经科学探索音乐协和性的脑与认知神经机制, 为理解音乐的基本原理提供了极丰富的资料。事实上, 中国早在春秋时期就建立了与古希腊相似的简单整数比的音乐思想(陈思, 陈其射, 2019; 薛冬艳, 2018)。甚至在律学方面, 明代的朱载堉率先利用自制的八十一档双排大算盘计算出“十二平均律”, 这比欧洲的同样成果早了近半个世纪。令人遗憾的是, 朱载堉的成果在当时并未引起重视, 而十二平均律在西方一经提出就迅速得到了推广, 并革新了乐器制造工艺技术和相应的作曲理论(Chow, 2020; 孙铿亮, 2017)。所以, 现代西方乐器具有律制规整, 泛音协调, 音响统一的特点。中国民族乐器却在音响上个性鲜明, 声学复杂, 且缺少低音乐器, 因而不适合进行大规模的交响合奏。直至今日, 中国民乐也未能够形成完整的音乐协和性理论。我们应当正视这种差距。尽管民乐从未停止过交响化的尝试与探索, 但在乐器改革的热潮下却出现了一些“全盘西化”的极端做法(唐朴林, 2010), 当然也存在反对的声音, 认为中国音乐具有独特的审美价值, 不应该以西方的标准来评价, 民乐追求的就应当是曲高和寡、高山流水的意境。但是, 科学不分东西, 西方交响音乐发展也是改革的结果, 但改革的方向却是受实证科学的牵引, 通过大量的心理声学和生理声学的实证研究建立了相应的音乐协和性理论, 并且广泛地应用于实际创作(Bennett, 2018; Chan et al., 2019)。需要强调的是, 本文只从音乐协和性这一基本听觉音乐现象角度指出科学研究的必要性, 不对音乐形式和艺术手法提出任何看法。中西方音乐各具特色, 但多线程的和声音乐无疑是未来音乐发展的必经之路。
陈思, 陈其射. (2019). 《老子》“一二三”的音乐阐微.(6), 125–134.
孙铿亮. (2017). “十二平均律”:从理论提出到键盘实践——j.s.巴赫对钢琴艺术的历史贡献.,(6), 255–256.
唐朴林. (2010). “乐改”何从?(2), 1–6.
王小盾. (2017). 上古中国人的用耳之道——兼论若干音乐学概念和哲学概念的起源.(4), 149–183.
薛冬艳. (2018). 声生于日, 律生于辰——阐发先秦、两汉二分、三分生律思维.(2), 64–72.
Ambrazevičius, R. (2017). Dissonance/roughness and tonality perception in Lithuanian traditional Schwebungs diaphonie.(1&2), 39–53.
Andermann, M., Patterson, R. D., & Rupp, A. (2020). Transient and sustained processing of musical consonance in auditory cortex and the effect of musicality.(4), 1320–1331.
Arnal, L. H., Flinker, A., Kleinschmidt, A., Giraud, A.-L., & Poeppel, D. (2015). Human screams occupy a privileged niche in the communication soundscape.(15), 2051–2056.
Ball, P. (2008). Facing the music.(7192), 160–162.
Belfi, A. M., Moreno, G. L., Gugliano, M., & Neill, C. (2021). Musical reward across the lifespan., 1–8.
Bennett, W. R., & Holland, C. K. (Eds). (2018).Switzerland, Cham: Springer Nature Switzerland AG.
Bernini, A., & Talamucci, F. (2014). Consonance of complex tones with harmonics of different intensity.(2), 78–89.
Bidelman, G. M. (2013). The role of the auditory brainstem in processing musically relevant pitch., 264.
Bidelman, G. M., & Grall, J. (2014). Functional organization for musical consonance and tonal pitch hierarchy in human auditory cortex., 204–214.
Bidelman, G. M., & Heinz, M. G. (2011). Auditory–nerve responses predict pitch attributes related to musical consonance–dissonance for normal and impaired hearing.(3), 1488–1502.
Bidelman, G. M., & Krishnan, A. (2009). Neural correlates of consonance, dissonance, and the hierarchy of musical pitch in the human brainstem.(42), 13165–13171.
Bones, O., Hopkins, K., Krishnan, A., & Plack, C. J. (2014). Phase locked neural activity in the human brainstem predicts preference for musical consonance., 23–32.
Bonin, T., & Smilek, D. (2016). Inharmonic music elicits more negative affect and interferes more with a concurrent cognitive task than does harmonic music.(3), 946–959.
Bowling, D. L., Hoeschele, M., Gill, K. Z., & Fitch, W. T. (2017). The nature and nurture of musical consonance.(1), 118–121.
Bowling, D. L., & Purves, D. (2015). A biological rationale for musical consonance.(36), 11155–11160.
Bowling, D. L., Purves, D., & Gill, K. Z. (2018). Vocal similarity predicts the relative attraction of musical chords.(1), 216–221.
Brattico, E., Pallesen, K. J., Varyagina, O., Bailey, C., Anourova, I., Järvenpää, M., ... Tervaniemi, M. (2009). Neural discrimination of nonprototypical chords in music experts and laymen: An MEG study.(11), 2230–2244.
Bravo, F., Cross, I., Hopkins, C., Gonzalez, N., Docampo, J., Bruno, C., & Stamatakis, E. A. (2020). Anterior cingulate and medial prefrontal cortex response to systematically controlled tonal dissonance during passive music listening.(1), 46–66.
Brown, S., & Jordania, J. (2013). Universals in the world’s musics.(2), 229–248.
Butler, J. W., & Daston, P. G. (1968). Musical consonance as musical preference: A cross-cultural study.(1), 129–142.
Chan, P. Y., Dong, M., & Li, H. (2019). The science of harmony: A psychophysical basis for perceptual tensions and resolutions in music.,, 2369041.
Chiandetti, C., & Vallortigara, G. (2011). Chicks like consonant music.(10), 1270– 1273.
Chow, S. (2020). A localised boundary object: Seventeenth- century western music theory in China., 75–113.
Christensen, T. (2004). Rameau and musical thought in the Enlightenment. In I. Bent, (general ed.),Cambridge: Cambridge University Press.
Coffey, E. B. J., Herholz, S. C., Chepesiuk, A. M. P., Baillet, S., & Zatorre, R. J. (2016). Cortical contributions to the auditory frequency–following response revealed by MEG., 11070.
Conard, N. J., Malina, M., & Münzel, S. C. (2009). New flutes document the earliest musical tradition in southwestern Germany.(7256), 737–740.
Cousineau, M., McDermott, J. H., & Peretz, I. (2012). The basis of musical consonance as revealed by congenital amusia.(48), 19858–19863.
Crespo-Bojorque, P., & Toro, J. M. (2015). The use of interval ratios in consonance perception by rats (rattus norvegicus) and humans (homo sapiens).(1), 42–51.
Crocker, R. L. (1963). Pythagorean mathematics and music.(2), 189–198.
Crowder, R. G., Reznick, J. S., & Rosenkrantz, S. L. (1991). Perception of the major/minor distinction: V. Preferences among infants.(3), 187–188.
de Cheveigné, A. (2005). Pitch perception models. In C. J. Plack & A. J. Oxenham (Eds.),(pp. 169–233). New York: Springer.
Dellacherie, D., Roy, M., Hugueville, L., Peretz, I., & Samson, S. (2011). The effect of musical experience on emotional self-reports and psychophysiological responses to dissonance.(3), 337–349.
di Stefano, N., Focaroli, V., Giuliani, A., Formica, D., Taffoni, F., & Keller, F. (2017). A new research method to test auditory preferences in young listeners: Results from a consonance versus dissonance perception study.(5), 699–712.
Dostrovsky, S. (1975). Early vibration theory: Physics and music in the seventeenth century.(3), 169–218.
Feng, L., & Wang, X. (2017). Harmonic template neurons in primate auditory cortex underlying complex sound processing.(5), E840–E848.
Fishman, Y. I., Volkov, I. O., Noh, M. D., Garell, P. C., Bakken, H., Arezzo, J. C., ... Steinschneider, M. (2001). Consonance and dissonance of musical chords: Neural correlates in auditory cortex of monkeys and humans.(6), 2761–2788.
Foss, A. H., Altschuler, E. L., & James, K. H. (2007). Neural correlates of the Pythagorean ratio rules.(15), 1521–1525.
Friedman, R. S., Kowalewski, D. A., Vuvan, D. T., & Neill, W. T. (2021). Consonance preferences within an unconventional tuning system.(3), 313–330.
Gill, K. Z., & Purves, D. (2009). A biological rationale for musical scales.(12), e8144.
Gold, B. P., Mas–Herrero, E., Zeighami, Y., Benovoy, M., Dagher, A., & Zatorre, R. J. (2019). Musical reward prediction errors engage the nucleus accumbens and motivate learning.(8), 3310–3315.
Greenwood, D. D. (1961). Auditory masking and the critical band.(4), 484–502.
Greenwood, D. D. (1991). Critical bandwidth and consonance in relation to cochlear frequency–position coordinates.(2), 164–208.
Guernsey, M. (1928). The role of consonance and dissonance in music.,(2), 173–204.
Harrison, P., & Pearce, M. T. (2020). Simultaneous consonance in music perception and composition.(2), 216–244.
Harrison, P. M. (2021). Three questions concerning consonance perception.(3), 337–339.
Helmholtz, H. (1885).. 2nd ed (A. J. Ellis, Trans). New York: Dover
Hoeschele, M., Cook, R. G., Guillette, L. M., Brooks, D. I., & Sturdy, C. B. (2012). Black-capped chickadee (Poecile atricapillus) and human (Homo sapiens) chord discrimination.(1), 57–67.
Hulse, S. H., Bernard, D. J., & Braaten, R. F. (1995). Auditory discrimination of chord-based spectral structures by European starlings (Sturnus vulgaris)., 409–423.
Itoh, K., Suwazono, S., & Nakada, T. (2010). Central auditory processing of noncontextual consonance in music: An evoked potential study.(6), 3781–3787.
Izumi, A. (2000). Japanese monkeys perceive sensory consonance of chords.(6), 3073–3078.
Kennedy, D., & Norman, C. (2005). What don’t we know.(5731), 75.
Kieffer, A. (2016). Riemann in France: Jean marnold and the “modern” music-theoretical ear.,(1), 1–15.
Killin, A. (2018). The origins of music: Evidence, theory, and prospects., 2059204317751971.
Kim, S. G., Lepsien, J., Fritz, T. H., Mildner, T., & Mueller, K. (2017). Dissonance encoding in human inferior colliculus covaries with individual differences in dislike of dissonant music.(1), 5726.
Koda, H., Basile, M., Olivier, M., Remeuf, K., Nagumo, S., Blois–Heulin, C., & Lemasson, A. (2013). Validation of an auditory sensory reinforcement paradigm: Campbell’s monkeys (Cercopithecus campbelli) do not prefer consonant over dissonant sounds.(3), 265–271.
Kohlrausch, A., Fassel, R., & Dau, T. (2000). The influence of carrier level and frequency on modulation and beat–detection thresholds for sinusoidal carriers.(2), 723–734.
Konoval, B. (2018). Pythagorean pipe dreams? Vincenzo galilei, marin mersenne, and the pneumatic mysteries of the pipe organ.(1), 1–51.
Masataka, N. (2006). Preference for consonance over dissonance by hearing newborns of deaf parents and of hearing parents.(1), 46–50.
Masataka, N., & Perlovsky, L. (2013). Cognitive interference can be mitigated by consonant music and facilitated by dissonant music., 2028.
McDermott, J. H., Lehr, A. J., & Oxenham, A. J. (2010). Individual differences reveal the basis of consonance.(11), 1035–1041.
McDermott, J. H., & Oxenham, A. J. (2008). Music perception, pitch, and the auditory system.(4), 452–463.
McDermott, J. H., Schultz, A. F., Undurraga, E. A., & Godoy, R. A. (2016). Indifference to dissonance in native Amazonians reveals cultural variation in music perception.(7613), 547–550.
McKinney, M., Tramo, M., & Delgutte, B. (2001). Neural correlates of musical dissonance in the inferior colliculus. In D. J. Breebaart, A. J. M. Houtsma, A. Kohlrausch, V. F. Prijs, & R. Schoonhoven, (Eds.),(pp.83–89). Maastricht, the Netherlands: Shaker Publishing.
McLachlan, N., Marco, D., Light, M., & Wilson, S. (2013). Consonance and pitch.(4), 1142–1158.
McPherson, M. J., Dolan, S. E., Durango, A., Ossandon, T., Valdés, J., Undurraga, E. A., ... McDermott, J. H. (2020). Perceptual fusion of musical notes by native Amazonians suggests universal representations of musical intervals.(1), 2786.
Mehr, S. A., Singh, M., Knox, D., Ketter, D. M., Pickens–Jones, D., Atwood, S., ... Glowacki, L. (2019). Universality and diversity in human song.(6468).
Meyer, M. (1903). Experimental studies in the psychology of music.(3/4), 192–214.
Milne, A. J., Laney, R., & Sharp, D. B. (2016). Testing a spectral model of tonal affinity with microtonal melodies and inharmonic spectra.(4), 465–494.
Minati, L., Rosazza, C., D'Incerti, L., Pietrocini, E., Valentini, L., Scaioli, V., ... Bruzzone, M. G. (2009). Functional MRI/event-related potential study of sensory consonance and dissonance in musicians and nonmusicians.(1), 87–92.
Montoya, R. M., Horton, R. S., Vevea, J. L., Citkowicz, M., & Lauber, E. A. (2017). A re-examination of the mere exposure effect: The influence of repeated exposure on recognition, familiarity, and liking.(5), 459–498
Norman-Haignere, S. V., Kanwisher, N., McDermott, J. H., & Conway, B. R. (2019). Divergence in the functional organization of human and macaque auditory cortex revealed by fMRI responses to harmonic tones.(7), 1057–1060.
Oxenham, A. J. (2018). How we hear: The perception and neural coding of sound., 27–50.
Pagès‐Portabella, C., & Toro, J. M. (2020). Dissonant endings of chord progressions elicit a larger ERAN than ambiguous endings in musicians.(2), e13476.
Palisca, C. V. (1961). Scientific empiricism in musical thought. In H. H. Rhys (Eds.),(Vol. 2361, pp. 91–137). Princeton University Press.
Parncutt, R., & Hair, G. (2011). Consonance and dissonance in music theory and psychology: Disentangling dissonant dichotomies.(2), 119–166.
Parncutt, R., Reisinger, D., Fuchs, A., & Kaiser, F. (2019). Consonance and prevalence of sonorities in Western polyphony: Roughness, harmonicity, familiarity, evenness, diatonicity.(1), 1–20.
Pear, T. H. (1911). The experimental examination of some differences between the major and the minor chord.(1), 56–88.
Perani, D., Saccuman, M. C., Scifo, P., Spada, D., Andreolli, G., Rovelli, R., ... Koelsch, S. (2010). Functional specializations for music processing in the human newborn brain.(10), 4758–4763.
Pesic, P. (2014).. Cambridge, Massachusetts: MIT Press.
Pisanski, K., & Feinberg, D. R. (2019). Vocal attractiveness. In S. Frühholz & P. Belin (Eds.),(pp. 607–625). New York: Oxford University Press.
Plack, C. J. (2010). Musical consonance: The importance of harmonicity.(11), R476–R478.
Plantinga, J., & Trehub, S. E. (2014). Revisiting the Innate Preference for Consonance.(1), 40–49.
Plomp, R., & Levelt, W. J. M. (1965). Tonal consonance and critical bandwidth.(4), 548–560.
Popescu, T., Neuser, M. P., Neuwirth, M., Bravo, F., Mende, W., Boneh, O., ... Rohrmeier, M. (2019). The pleasantness of sensory dissonance is mediated by musical style and expertise.(1), 1–11.
Popham, S., Boebinger, D., Ellis, D. P. W., Kawahara, H., & McDermott, J. H. (2018). Inharmonic speech reveals the role of harmonicity in the cocktail party problem.(1), 2122
Postal, O., Dupont, T., Bakay, W., Dominique, N., Petit, C., Michalski, N., & Gourévitch, B. (2020). Spontaneous mouse behavior in presence of dissonance and acoustic roughness.588834.
Prete, G., Bondi, D., Verratti, V., Aloisi, A. M., Rai, P., & Tommasi, L. (2020). Universality vs experience: A cross-cultural pilot study on the consonance effect in music at different altitudes., e9344.
Primavesi, O. (2016). Empedocles’ cosmic cycle and the pythagorean tetractys.(1), 5–29.
Proverbio, A. M., Orlandi, A., & Pisanu, F. (2016). Brain processing of consonance/dissonance in musicians and controls: A hemispheric asymmetry revisited.(6), 2340–2356.
Purves, D. (2017).Cambridge: Harvard University Press.
Scharinger, M., Idsardi, W. J., & Poe, S. (2011). A comprehensive three-dimensional cortical map of vowel space.(12), 3972– 3982.
Schneider, A. (2018). Pitch and pitch perception. In R. Bader (Eds.),(pp. 605–685). Berlin, Heidelberg: Springer.
Schwartz, D. A., Howe, C. Q., & Purves, D. (2003). The statistical structure of human speech sounds predicts musical universals.(18), 7160–7168.
Seror, G. A., & Neill, W. T. (2015). Context dependent pitch perception in consonant and dissonant harmonic intervals.(5), 460–469.
Shapira, L. I., & Stone, L. (2008). Perception of musical consonance and dissonance: An outcome of neural synchronization.(29), 1429–1434.
Stolzenburg, F. (2015). Harmony perception by periodicity detection.(3), 215– 238.
Stumpf, C. (1989). Konsonanz and dissonanz., 91–107.
Sugimoto, T., Kobayashi, H., Nobuyoshi, N., Kiriyama, Y., Takeshita, H., Nakamura, T., & Hashiya, K. (2010). Preference for consonant music over dissonant music by an infant chimpanzee.(1), 7–12.
Sukljan, N. (2020). Renaissance music between science and art.(2), 183–206.
Tabas, A., Andermann, M., Schuberth, V., Riedel, H., Balaguer-Ballester, E., & Rupp, A. (2019). Modeling and MEG evidence of early consonance processing in auditory cortex.(2), e1006820.
Tenney, J. (1988).. New York: Excelsior Music Publishing.
Toro, J. M., & Crespo-Bojorque, P. (2017). Consonance processing in the absence of relevant experience: Evidence from nonhuman animals., 33–44.
rainor, L. J., & Heinmiller, B. M. (1998). The development of evaluative responses to music:: Infants prefer to listen to consonance over dissonance.(1), 77–88.
Trainor, L. J., Tsang, C. D., & Cheung, V. H. (2002). Preference for sensory consonance in 2–and 4–month–old infants.(2), 187–194.
Tramo, M. J., Cariani, P. A., Delgutte, B., & Braida, L. D. (2001). Neurobiological foundations for the theory of harmony in western tonal music.(1), 92–116.
Valentine, C. W. (1914). The method of comparison in experiments with musical intervals and the effect of practice on the appreciation of discords.(1), 118–135.
Vassilakis, P. N. (2005). Auditory roughness as a means of musical expression., 119–144.
Vencovskỳ, V. (2016). Roughness prediction based on a model of cochlear hydrodynamics.(2), 189–201.
Virtala, P., Huotilainen, M., Partanen, E., Fellman, V., & Tervaniemi, M. (2013). Newborn infants' auditory system is sensitive to Western music chord categories., 492.
Virtala, P., & Tervaniemi, M. (2017). Neurocognition of major–minor and consonance–dissonance.(4), 387–404.
von Békésy, G. (1960).. New York: McGraw Hill.
Watanabe, S., Uozumi, M., & Tanaka, N. (2005). Discrimination of consonance and dissonance in Java sparrows.(2), 203–208.
Weiss, M. W., Cirelli, L. K., McDermott, J. H., & Trehub, S. E. (2020). Development of consonance preferences in Western listeners.(4), 634–649.
Werker, J. F., & Hensch, T. K. (2015). Critical periods in speech perception: New directions., 173–196.
Zatorre, R. J., & Salimpoor, V. N. (2013). From perception to pleasure: Music and its neural substrates.(Suppl. 2), 10430– 10437.
Zentner, M. R., & Kagan, J. (1996). Perception of music by infants.(6595), 29–29.
Zhou, L., Liu, F., Jiang, J., & Jiang, C. (2019). Impaired emotional processing of chords in congenital amusia: Electrophysiological and behavioral evidence., 103577.
Why musical emotion can be induced by harmony? The effect and cognitive mechanism of musical consonance
ZHANG Hang1, MENG Le1, ZHANG Jijia2
(1Department of Psychology, Renmin University of China, Beijing 110872, China)(2Faculty of Education, Guangxi Normal University, Guilin 541004, China)
Music is an advanced activity of human cognition and one of the universal ways of emotional expression in life. As the core element connecting music and emotion, the rationale for simultaneous consonance is still unsolved. The core of this problem is how our brain possesses musical chords composed of several tones and why some tone combinations sound relatively pleasant (consonance) while others sound unpleasant (dissonance). This question has fascinated scholars since the ancient Greeks. Physicists have been trying to find answers to the differences between acoustic features of consonance and dissonance harmony. Biologists argue that consonance perception is the basic emotional experience evoked by sound events in the auditory system. Psychologists are more inclined to examine whether musical consonance perception is nature or nurture. Although musical consonance has been researched mainly using western theoretical perspectives, studying musical consonance in Chinese traditional music culture is urgently needed.
musical consonance, harmonicity, beating, musical culture, cognitive mechanism
2021-04-17
孟乐为共同第一作者。
张积家, E-mail: Zhangjj1955@163.com
B842, B845
猜你喜欢
乐府新声(2022年1期)2022-03-30
儿童时代·快乐苗苗(2021年11期)2022-01-22
音乐研究(2019年5期)2019-11-22
黄河之声(2018年20期)2018-01-24
黄梅戏艺术(2017年1期)2017-07-04
海峡姐妹(2017年5期)2017-06-05
琴童(2017年3期)2017-04-05
戏剧之家(2016年18期)2016-10-27
戏剧之家(2016年17期)2016-10-17
中华老年多器官疾病杂志(2016年8期)2016-05-14
