
江西稻瘟病自然病圃菌群毒性相关基因的鉴定与分析
2022-05-07 00:56霍光华李湘民杨迎青
江西农业大学学报 2022年2期
兰 波,余 建,霍光华,李湘民,杨迎青*
(1.江西省农业科学院 植物保护研究所,江西 南昌 330200;2.江西农业大学 菌物资源保护与利用江西省重点实验室,江西 南昌 330045)
【研究意义】由稻瘟病菌Magnaporthe oryzae(无性态:Pyricularia oryzae)引起的稻瘟病是世界水稻生产中三大毁灭性病害之一,对水稻的产量和品质均造成严重影响[1],已成为水稻高产、稳产的主要限制因子。江西是水稻种植大省,稻米产量占中国的第三位,然而江西地处温暖湿润的亚热带区也是稻瘟病常发、重发区,一般早稻发病较重,中稻次之,而以晚稻发病较轻。如2006 年发生面积5.7×105hm2,其中早稻发生面积达4.5×105hm2[2]。实践证明,利用品种自身的抗性控制稻瘟病是最经济有效的方法。但是,稻瘟菌毒性相关基因高频率变异使得病原菌小种极不稳定,常导致新培育出的抗稻瘟病品种在推广3~5年后抗性逐渐减弱或者丧失[3]。【前人研究进展】由于水稻寄主与稻瘟病菌之间的互作符合Flor 的“基因对基因”学说,二者存在典型的特异性识别和互作规律[4]。因此,开展稻瘟病菌群体结构与毒性基因研究、掌握病原菌群体变化规律及毒性信息,对于品种选育和抗病品种合理布局都具有重要的意义[5-6]。目前已克隆和分析了近40 个与稻瘟病菌生活史中各个阶段相关的功能基因[7]。其中MPG1,MPS1,MagB,CPKA,MNH1,MgYCA1,MgS11和MgATG5 为稻瘟病菌重要的致病相关基因。如CPKA编码蛋白激酶A的催化亚基PKA-c是稻瘟病菌致病性所必需的基因[8],MPG1功能作用于稻瘟病菌对外界信号的识别[9];MPS1与附着胞的侵入、稻瘟病菌的营养生长、产孢和子囊形成密切相关[10]。稻瘟病菌无毒基因研究相对较少,已成功克隆的无毒基因仅有几个[11],如Avr-pia,Avr-pit,Avr-pik,Avr-pita,Avr-pizt,Avrco39,PRE1和ACE1在寄主与病原菌互作发生抗病反应过程中发挥重要的作用。其中无毒基因Avr-pita编码223个氨基酸的金属蛋白酶,能与抗性基因Pi-ta直接互作,激活特异性免疫反应[12];AVR1-CO39可以特异的激发有抗病基因Pi-Co39(t)水稻的防卫反应[13]。近年来,国内学者也开展了部分地区的稻瘟病菌无毒基因分子检测,朱名海等检测了Ace1、Avr-Pia、Avr-Pik、Avr-Pita、Avr-Pizt和PWL2等6 个无毒基因在南繁核心区与非核心区稻瘟病菌中的分布频率,结果表明不同区域稻瘟病菌中无毒基因存在一定差异[14]。郭敬玮等(2021)利用特异性引物标记测定了Avr-Pia等7 个无毒基因在云南罗平120 个田间菌株中的存在情况[15]。邓云等(2021)通过测定24 个无毒基因在福建省稻瘟病菌中的出现频率,明确了Avr-z5、Avr-9(t)这两个无毒基因出现频率最高而且稳定,对应的抗病基因在今后的抗性育种中可以重点应用[16]。而江西在利用分子鉴定方法测定稻瘟病菌中致病基因和无毒基因却未见报道。【本研究切入点】本研究于2020年在江西5个稻瘟病自然诱发病圃采集分离的189个稻瘟病菌单孢菌株,利用16个已克隆或鉴定的具有重要功能的稻瘟病菌毒性相关基因的特异性引物进行PCR 检测,明确相关基因在菌群中的分布情况。【拟解决的关键问题】通过监测江西省不同地区稻瘟病菌的毒性相关基因的分布,掌握菌群遗传结构变化可为江西水稻抗病育种、抗病品种合理布局以及延长使用周期、稻瘟病综合防控提供科学依据。
1 材料与方法
1.1 供试菌株
2020 年从江西5 个稻瘟病自然诱发病圃种植的高感品种丽江新团黑谷(LTH)病穗颈标本分离纯化的189个稻瘟病菌单胞菌株,其中都昌稻瘟病病圃42株(编号DC01~DC42),井冈山稻瘟病病圃44株(编号JGS01~JGS44),丰城稻瘟病病圃39株(编号FC01~FC39),万安稻瘟病病圃32株(编号WA01~WA32),婺源稻瘟病病圃32株(编号WY01~WY32)。
1.2 稻瘟病菌菌丝体培养
将供试菌株活化后再纯化一次,挑取适量菌丝在PDA 液体培养基中于转速120 r/min摇床上28 ℃培养3~5 d,待菌丝体生长至一定量后用灭菌纱布过滤收集、烘干,-20 ℃保存备用。
1.3 稻瘟病菌基因组DNA提取
将收集的菌丝体分装于2 mL 的Eppendorf 管中,加2 颗5 mm 灭菌碳钢珠后迅速加入65 ℃预热的CTAB 提取液,应用上海净信科技生产的全自动样品快速研磨仪进行磨样,使用CTAB[18]法进行基因组DNA提取,最后用RNAase进行处理后抽提一次,-20 ℃保存备用。
1.4 供试菌株致病基因与无毒基因测定
采用公开报道的8 个致病基因MNH1、MgATG5、MPG1、MgYCA1、MgS11、MagB、MPS1、CPKA,8 个无毒基因Avr-pia、Avr-pizt、ACE1、Avr-pit、Avr-pita、Avr-co39、Avr-pik,PRE1共16 对特异性SSR 标记引物(表1)[17-19],经PCR 扩增对供试稻瘟病菌的毒性相关基因的分子检测。PCR 反应体系(20µL):TaqDNA聚合酶0.2µL,10×Buffer(MgCl2)2µL,正、反向引物各1µL(0.2µmol/L),dNTP 0.5µL(2.5 mmol/L),DNA模板1µL(≤200 ng),ddH2O补足至20µL。PCR反应程序:95 ℃预变性5 min,94 ℃变性30 s,50~62 ℃(根据不同引物退火温度)退火30 s,72 ℃延伸30~60 s,具体依据目的基因片段长度而定,35个循环,72 ℃延伸10 min。琼脂糖凝胶电泳检测:取PCR 产物5µL与适量的6×Loading buffer 混合液混合,在15 g/L琼脂糖凝胶中电泳(电压100 V,时间20 min),染色后在凝胶成像系统下拍照。

表1 稻瘟病菌致病基因与无毒基因分子检测特异性引物标记序列Tab.1 Molecular detection of pathogencity genes and avirulence genes of Magnaporthe oryzae using specific primer-marking sequences
1.5 数据统计与分析
根据PCR扩增结果,建立基于稻瘟病菌毒性相关基因指纹的“0~1”数据库。根据每个样品电泳条带的有无,有则赋值为“1”,无则赋值为“0”。统计各引物的扩增频率,并应用Ntsys 数据处理系统的Nei&Li方法[20]计算遗传距离,用最长距离法对16对引物的扩增情况进行聚类,分析稻瘟病菌群体遗传宗谱。
2 结果与分析
2.1 稻瘟病菌无毒基因在江西菌群的PCR测定结果
8对稻瘟病菌无毒基因特异性引物扩增结果显示均能扩增出符合目的片段大小的指纹条带,部分菌株扩增电泳条带结果如图1所示。无毒基因在5个地区病圃的稻瘟病菌中检测频率差异较大。其中,都昌病圃菌株中8 个无毒基因的检测频率为0~83.33%,Avr-Pik最高为83.33%,Avr-Pit与PRE1未检测到。井冈山病圃菌株中8 个无毒基因的检测频率为0~88.64%,Avr-Pizt最高为88.64%,Avr-Pit未检测到。丰城病圃菌株中8个无毒基因的检测频率为2.56%~100%,Avr-Pizt最高为100%,Avr-Pit最低为2.56%。万安病圃菌株中8 个无毒基因的检测频率为3.13%~100%,Avr-Pizt与Avr-Pia最高均为100%,Avr-Pit最低为3.13%。婺源病圃菌株中8 个无毒基因的检测频率为0~100%,Avr-Pik、Avr-co39与PRE1最高均为100%,Avr-Pit最低为0,未检测到(表2)。

图1 部分稻瘟病菌无毒基因Avr-Pik PCR检测结果Fig.1 The PCR detection results of avirulence gene Avr-Pik from partial strains of Magnaporthe oryzae

表2 稻瘟病菌无毒基因在江西菌株中的分布检测结果Tab.2 Distribution of avirulence genes of Magnaporthe oryzae in strains isolated from Jiangxi
2.2 稻瘟病菌致病基因在江西菌群的PCR测定结果
8 对稻瘟病致病基因的特异性引物除MNH1均能扩增出符合目的基因条带,部分PCR 检测结果如图2 所示。供试的稻瘟病菌致病基因在5 个地区病圃的稻瘟病菌中检测频率差异较大。其中,都昌病圃菌株中8 个致病基因的检测频率为0~90.48%,MPG1最高为90.48%,MNH1未检测到。井冈山病圃菌株中8 个致病基因的检测频率为0~93.18%,MPG1与MagB最高均为93.18%,MNH1未检测到为0。丰城病圃菌株中8 个致病基因的检测频率为0~100%,MPG1最高为100%,MNH1未检测到为0。万安病圃菌株中8 个致病基因的检测频率为0~100%,MPG1、MPS1与MgS11最高均为100%,MNH1未检测到为0。婺源病圃菌株中8 个致病基因的检测频率为0~100%,MPG1最高为100%,MNH1最低为0,未检测到(表3)。
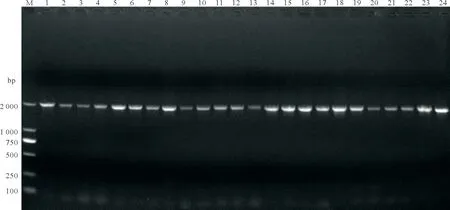
图2 部分稻瘟病菌致病基因MagB检测结果Fig.2 The PCR detection results of pathogencity gene MagB from partial strains of Magnaporthe oryzae

表3 稻瘟病菌致病基因在江西菌株中的分布检测结果Tab.3 Distribution of pathogencity genes of Magnaporthe oryzae in strains isolated from Jiangxi
2.3 江西稻瘟病菌群体遗传宗谱分析
将5个地区病圃稻瘟病菌中检测的16个毒性相关基因结果按顺序排列,用专业软件进行聚类分析,聚类结果见图3。在本研究中以0.72遗传相似水平下分析稻瘟病菌群体结构特征层次比较丰富(表4),遗传宗谱划分较为合理。在0.72 遗传相似水平下,江西5 个地区病圃稻瘟病菌群可以划分为11 个遗传宗谱。其中宗谱JX1 占总菌株数的28.57%,宗谱JX2 占总菌株数的36.51%,宗谱JX3 占总菌株数的10.58%,宗谱JX4 占总菌株数的2.65%,宗谱JX5 占总菌株数的1.59%,宗谱JX6 占总菌株数的1.06%,宗谱JX7 占总菌株数的7.41%,宗谱JX8 占总菌株数的5.29%,宗谱JX9 占总菌株数的1.59%,宗谱JX10 占总菌株数的0.53%,宗谱JX11占总菌株数的4.23%。宗谱JX1与JX2为优势宗谱,JX3为婺源病圃菌株特有宗谱,其他宗谱类型为小宗谱或稀有宗谱,具有地域特有性特征。

表4 在遗传系数0.72水平下稻瘟病菌群体的遗传宗谱Tab.4 Genetic lineages of Magnaporthe oryzae strains at the genetic similarity coefficient level of 0.72

图3 稻瘟病菌毒性基因检测结果聚类分析Fig.3 Cluster map based on virulence genes detection of Magnaporthe oryzae
3 结论与讨论
稻瘟病菌在长期的进化过程中,其群体遗传结构随品种、地理环境、栽培方式等因素变化而发生变异,病原菌毒性相关基因及其致病型的组成和变异直接关系到稻瘟病的发生和流行[21]。监测区域稻瘟病毒性基因特别是无毒基因,对地区的病害防控具有科学的指导意义。传统方法是利用含有单基因抗瘟品种接种稻瘟病菌,根据抗感反应推导菌株含有与抗瘟基因相对应的无毒基因,该方法由于受操作人员、环境等因素影响结果准确率,且工作量大,劳动强度高。随着越来越多的稻瘟病菌无毒基因被鉴定与克隆,利用特异性引物来检测无毒基因被认为是目前最有效的手段。
本研究利用已公开报道克隆的8 个致病基因和8 个无毒基因特异性引物测定了2020 年从江西5 个稻瘟病自然病圃中分离的189 个单孢菌株毒性相关基因的分布情况。结果表明:8 个无毒基因Avr-pia、Avr-pizt、ACE1、Avr-pit、Avr-pita、Avr-co39、Avr-pik、PRE1在江西都昌、井冈山、丰城、万安、婺源5 个稻瘟病自然病圃稻瘟病菌的检测频率存在较大差异。其中都昌菌株中Avr-pik检测频率最高,井冈山、丰城、万安菌株中Avr-pizt检测频率均最高,婺源菌株中Avr-pik、Avr-co39、PRE1检测频率均最高为100%,而Avr-pit在所有地区菌株中检出频率都是最低。因此,与检出频率高相对应的稻瘟病抗病基因在江西具有重要的应用潜力,而与检出频率低相对应的稻瘟病抗病基因应当慎用。8 个致病基因MNH1、MgATG5、MPG1、MgYCA1、MgS11、MagB、MPS1、CPKA在江西5 个地区稻瘟病自然病圃菌株中只检测到7 个,MNH1未检测到,且存在较大的分布差异。MPG1在5 个地区病圃稻瘟病菌的检测频率均为最高在90%以上,可见江西稻瘟病菌遗传多样性较丰富。
基于稻瘟病菌毒性相关基因的遗传宗谱测定更符合病菌本身的遗传特性。本研究中江西5 个稻瘟病病圃菌株在16个稻瘟病菌毒性相关基因测定结果聚类被划分为11个遗传宗谱,其中宗谱的菌株比例存在很大的差异,3 个优势宗谱JX1、JX2、JX3 占总菌株数的75.66%,其他8 个小宗谱或稀有宗谱只占总菌株数的24.34%,结果表明江西稻瘟病菌株遗传结构丰富,但又存在优势种群。此外,在不同地区的稻瘟病菌群存在着独特的遗传宗谱类型,比如宗谱JX3 只存在婺源稻瘟病圃菌株中,宗谱JX5 只存在都昌稻瘟病圃菌株中,宗谱JX6 与JX10 只存在井冈山稻瘟病圃菌株中。不同地区的宗谱多样性使得稻瘟病菌的毒性也不同,这也与笔者多年多点的水稻品种稻瘟病抗性鉴定结果一致,相同的品种在不同地区的稻瘟病病圃中的抗性表现存在差异。
由于田间稻瘟病菌具有易变性的特点,尤其是无毒基因的变异是导致水稻抗瘟品种抗性丧失的主要原因,抗瘟品种的持久抗性与其种植地区稻瘟菌致病基因和无毒基因的变化密切相关[22]。因此,对不同地区稻瘟病菌致病基因和无毒基因分布测定和动态变化监测,指导当地水稻抗瘟品种的合理布局,是有效防控稻瘟病流行发生的重要手段,也是一项长期性的基础工作。本研究结果显示江西省稻瘟病菌无毒基因与致病基因组成复杂多样,且遗传宗谱具有地域特有性特点。从稻瘟病菌无毒基因检测结果来看,在江西水稻品种安全布局中,含有抗性基因Pia、Pizt和Pik的品种具有较好的推广价值。
致谢:江西现代农业科研协同创新专项(JXXTCX201901,JXXTCXBSJJ202119)同时对本研究给予了资助,谨致谢意!
猜你喜欢
食品安全导刊(2022年11期)2022-11-17
新疆农业科学(2022年6期)2022-07-13
现代园艺(2022年9期)2022-06-07
中国农学通报(2022年12期)2022-06-01
作物学报(2022年6期)2022-04-08
草地学报(2022年3期)2022-03-28
作物学报(2022年4期)2022-02-10
农民致富之友(2020年15期)2020-05-25
中学生物学(2019年7期)2019-10-17
江苏农业学报(2019年1期)2019-09-10
