
千年桐根部黄酮类化合物生物合成对枯萎病菌侵染的响应*
2022-05-11 11:54梁晓洁吴立文赵耘霄汪阳东黄世清张永志傅火勇陈益存
林业科学 2022年2期
王 嘉 梁晓洁 高 暝 吴立文 赵耘霄 汪阳东 黄世清 张永志 傅火勇 陈益存
(1.中国林业科学研究院亚热带林业研究所 杭州 311400; 2.南京林业大学 南京 210037; 3.安吉县龙山林场 湖州 313306)
植物枯萎病是一种土传的真菌病害,已引起100多种植物栽培种发生枯萎病危害(Deanetal., 2012)。有一些植物种类或品种对枯萎病有显著的抗性,可为防治该病提供思路。油桐(Vernicia)是我国传统的重要化工油料树种,已有几千年的栽培历史,由其种子提炼而得的桐油是一种优质干性油,广泛应用于工业、农业、渔业、军事及医药等领域。三年桐(Verniciafordii)是我国油桐的主栽品种,具有生长快、结实早、产量高的特征,但规模化种植面临严重的枯萎病危害。油桐枯萎病又称“桐瘟”,是由尖孢镰刀菌(Fusariumoxysporumf. sp.fordii(Fof-1))引起的致死性土传病害(Chenetal., 2016)。迄今油桐枯萎病已在我国8个省份90多个县市爆发,对现有的近百万公顷油桐林产生了严重威胁(杨素素等, 2014)。相比而言,同属的千年桐(Verniciamontana)具有高抗枯萎病能力,但是生长缓慢、结实晚、产油量和质量低(方嘉兴, 2017)。目前,以千年桐为砧木、三年桐为接穗的嫁接油桐是防治枯萎病的有效方法。
随着系统生物学的不断完善和发展,利用转录组和代谢组学在植物抗病研究取得了重要进展。基于高通量测序的转录组技术已在植物抗病方面得到广泛的应用,如通过比较转录组分析了枯萎病菌侵染不同时期千年桐、三年桐差异表达基因的表达模式(Chenetal., 2016); 利用转录组分析探索了杨树(Populusspp.)受细菌性溃疡病菌(Lonsdaleaquercina)诱导上调表达的关键基因(Houetal., 2016)。代谢组学是继基因组学、转录组学和蛋白组学之后兴起的一门新的组学技术,可以解决许多科学问题(Thévenotetal., 2015)。由于转录组学和代谢组学各有自身的优缺点,通常对它们进行联合分析对生物学问题进行解释。如研究“鲍威尔”脐橙(Citrussinensis)对指状拟青霉(Penicilliumdigitatum)侵染后的防御响应,发现鼠李糖、肌醇等代谢物的积累增强了脐橙对病菌侵染的抵抗力,通过茉莉酸和乙烯途径以及增加ERFs、WRKYS和MYB等转录因子的转录丰度来触发防御反应,并且苯丙烷生物合成途径也在转录水平上被激活(Tangetal., 2018); 通过脐橙抗病突变体的代谢组和转录组联合分析发现突变体中脂肪酸通路重定向,刺激茉莉酸的生物合成和信号传导途径而参与防御反应(Heetal., 2018)。
本研究探讨了三年桐和千年桐根部提取物对Fof-1生长是否有抑制作用,通过代谢组和转录组联合分析明确了千年桐根部抗枯萎病关键代谢物、响应途径及途径中的关键基因,研究结果为阐明千年桐的抗枯萎病机制提供新的思路。
1 材料与方法
1.1 试验材料
植物材料为三年桐、千年桐品种1年生实生苗,种源来自贵州独山县油桐主产区,种植于中国林业科学研究院亚热带林业研究所后山温室大棚。油桐病原菌Fof-1是实验室前期从感枯萎病的三年桐根中分离并鉴定,由本课题组保留(Chenetal., 2016)。病原菌Fof-1采用马铃薯葡萄糖琼脂(PDA)培养基进行培养,孢子制备采用合成低营养琼脂培养基(SNA)。
1.2 三年桐、千年桐根部提取物对病原菌Fof-1的抑制作用差异
取生长良好的三年桐、千年桐幼根烘干后研磨过60目筛。分别称取20 g烘干的样品,用200 mL的95%乙醇超声提取2 h,滤纸过滤后,再用100 mL无水乙醇超声波提取1 h,用滤纸过滤后使用旋转蒸发仪蒸发浓缩,加水稀释至30 mL,并用相同体积的乙酸乙酯萃取,有机样分别蒸发浓缩到膏状,用乙酸乙酯溶解至4 mL。将1 mL等量的油桐根部提取物母液均匀涂抹于PDA平板上,设置:千年桐母液、三年桐母液、乙酸乙酯(萃取剂)对照和纯PDA对照。每组设置3个生物学重复。将直径5 mm圆形油桐枯萎病菌菌块置于PDA平板中央。将平板放置在28 ℃培养箱中暗培养。记录菌落的生长情况和菌落直径。采用十字交叉法测量菌落直径并计算相对抑菌率(相对抑菌率=(对照菌丝生长直径-处理菌丝生长直径)/ 对照菌丝生长直径×100%)。
1.3 病原菌Fof-1接种和样本采集
取生长良好的Fof-1菌块接种于SNA培养基中,28 ℃培养7 天,配置2×106个·mL-1的尖孢镰刀菌孢子悬浮液。挑取生长良好的三年桐、千年桐幼苗,将根表面土清洗干净,然后将整个根系置于75%乙醇1 min,0.5%次氯酸钠处理3 mins,90%酒精溶液浸泡30 s,无菌水冲洗3次。用大头针将主根和侧根轻轻划几个伤口后,在孢子悬浮液中浸泡30 min。将接种后的植株重新种植于无菌土中,放置于28 ℃的人工气候箱中培养,湿度控制在90%左右,光照周期设置为16 h光照,8 h黑暗,光照强度为5 000 lx。三年桐和千年桐各处理30株,10株未感染植株作为对照。
根据枯萎病的症状,分别在第0、1、5和8天收集根系,液氮中速冻后保存在-80 ℃,三年桐和千年桐样本分别被标记为未侵染(F0; M0)、侵染早期(F1; M1)、侵染中期(F2; M2)和侵染晚期(F3; M3)。
1.4 根部次生代谢物提取及测定
选用Fof-1侵染后期的三年桐、千年桐根部样品(F3,M3)及其3个生物重复,经真空冷冻干燥,研磨至粉末状; 称取100 mg的粉末溶解于1 mL提取液(含有0.1 mg·L-1利多卡因(内标)的70%的甲醇水溶液)中; 4 ℃冰箱过夜,期间涡旋3次,提高提取率; 离心(转速10 000 r·min-1,10 mins)后,吸取上清,用微孔滤膜(0.22 μm孔径)过滤样品,并保存于进样瓶中,用于高效液相色谱和串联质谱分析。
样品在完成高效液相色谱和串联质谱代谢物检测后,通过Analyst 1.6.1软件打开“WIFF”格式的原始数据,用于定性和定量分析。过滤和去除多余的原始数据; 根据公共数据库对代谢物进行定性分析,包括MassBank、KNAPSAcK、METLIN(Zhuetal., 2013)、MoTo DB和HMDB(Wishartetal., 2013)等; 为了确保定量分析的准确性,对每个代谢物在三年桐和千年桐样本中检测到的质谱峰进行校正; 主成分分析(PCA)和(正交)偏最小二乘判别分析[(O) PLS-DA]; 基于(O) PLS-DA模型的变量投影重要性(Variable Importance in Project,VIP),可以初步筛选三年桐和千年桐之间显著不同的代谢物,结合单变量分析的差异倍数值来进一步筛选出差异代谢物(VIP ≥ 1,差异倍数 ≥ 2; 差异倍数 ≤ 0.5),并绘制火山图和聚类热图。
1.5 病原菌Fof-1菌侵染后基因表达规律分析
1.5.1 cDNA文库构建和转录组测序 利用北京艾德莱生物公司的植物RNA快速提取试剂盒对侵染前后不同时期的三年桐(F0,F1,F2,F3)、千年桐根部组织(M0,M1,M2,M3)进行RNA提取,通过微量分光光度计Q5000检测RNA浓度及纯度(OD260/OD280、OD260/OD230),并用1%的琼脂糖凝胶电泳对RNA质量进行进一步检测。取质检合格的RNA样品构建cDNA文库,在Illumina HiSeqTM2000上对cDNA文库进行测序,测序策略为双末端测序。
1.5.2 基因注释和差异表达基因分析 将拼接好的转录本序列使用Blast算法(E值 ≤ 1e-5)在以下数据库中进行基因注释,包括Nr(NCBI non-redundant protein sequences)、Nt(NCBI non-redundant nucleotide sequences)、Swiss-Prot(A manually annotated and reviewed protein sequence database)、Pfam(Protein family)、KOG(Clusters of Orthologous Groups of proteins)、GO(Gene Ontology)、KO(KEGG Ortholog database)。利用GENscan软件对基因编码区进行预测,得到编码区的核酸序列和氨基酸序列。使用预测每千碱基的读数/百万位图读数据(Reads per kilobase per million mapped read,RPKM)分析转录本丰度,以评估基因表达水平。根据RPKM值,使用DESeq R软件包查找差异表达基因(差异倍数 ≥ 2,P< 0.05)。为明确Fof-1侵染过程中三年桐、千年桐的不同基因表达模式。根据差异表达基因的log2RPKM值进行了K-均值聚类分析,并对差异表达基因进行KEGG富集分析。
1.5.3 苯丙烷生物合成途径的共表达网络分析 利用R软件包和加权基因共表达网络分析(WGCNA),选取加权截止值 > 0.50(weighted cut-off value)的基因去构建苯丙烷类生物合成途径中的基因网络,并使用Cytoscape软件(https:∥cytoscape.org/download.html)对共表达网络进行可视化。
1.6 实时定量PCR(qRT-PCR)验证
使用Primer Premier 5.0对苯丙烷生物合成途径中的4个关键基因的相应序列的非保守区域设计了特异性引物。每个样品进行3个独立的生物学重复,并进行3次技术重复分析。根据SYBR®Premix Ex TaqTM试剂盒的操作说明,在ABI7300实时定量仪器上以20 μL反应体积进行qRT-PCR分析。扩增条件为95 ℃、30 s,然后在95 ℃ 40循环5 s,然后在60 ℃退火34 s。千年桐转录延伸因子1(TEF1)用作内参以确定基因表达的正常性,并使用2-ΔΔCT方法计算基因的相对表达水平。
2 结果与分析
2.1 千年桐根部化合物对Fof-1的生长具有明显的抑制作用
与三年桐相比,千年桐根部提取物对Fof-1的生长表现出明显的抑制作用,第5天抑制率达到最高,为43.2%; 在第9天,三年桐、千年桐根提取物的抑制率差异最大(图1)。
2.2 千年桐根部黄酮类化合物在Fof-1侵染后含量上升
病原菌Fof-1侵染后的三年桐、千年桐样品(M3,F3)经UPLC和MS/MS代谢物检测,共鉴定了622种已知代谢物,分为33类。在622种代谢产物中,8类代谢产物占据半数以上,包括有机酸(11.25%),氨基酸衍生物(9.16%),核苷酸及其衍生物(8.68%),羟基肉桂酰基衍生物(5.14%),黄酮(4.98%),脂质甘油磷脂(4.98%),黄酮醇(4.66%)和氨基酸(4.50%)。
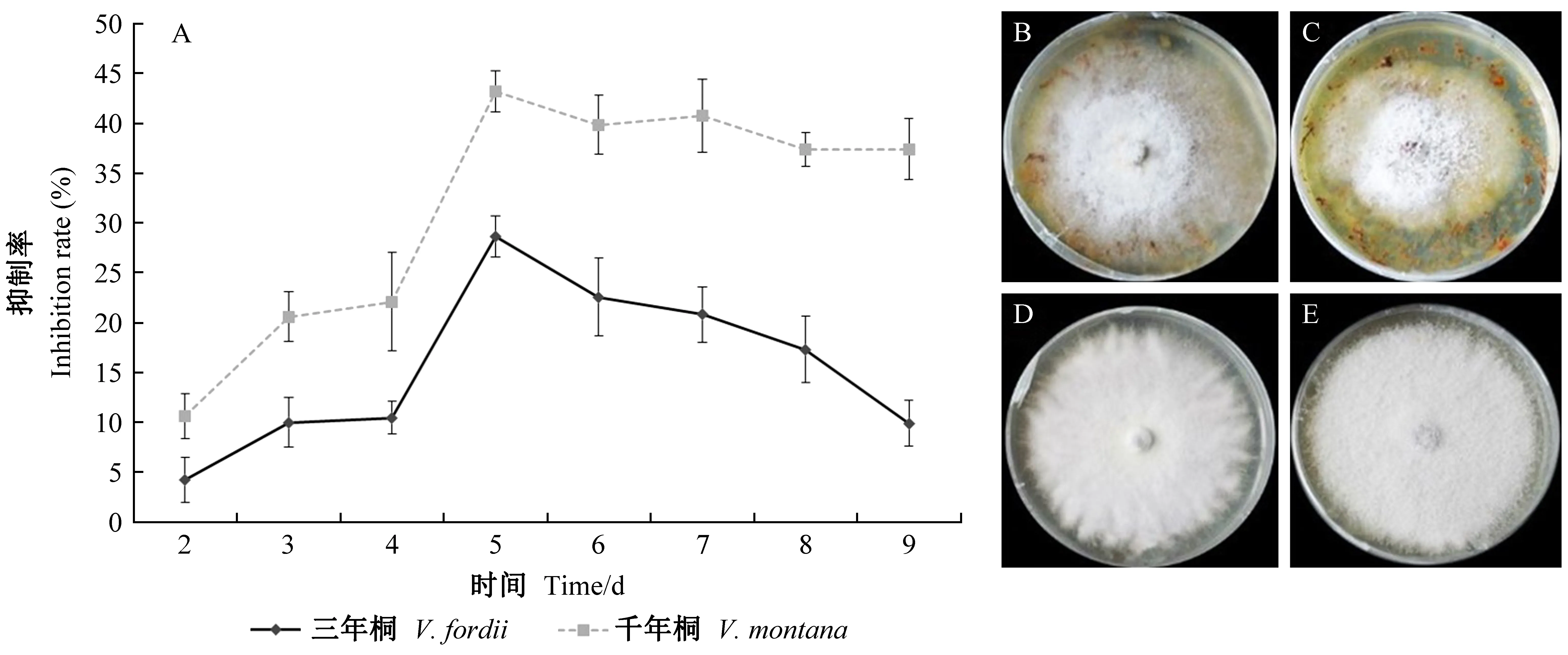
图1 三年桐、千年桐根部化合物对Fof-1的抑制作用差异
为了明确病原菌Fof-1侵染后三年桐、千年桐根部代谢物含量差异和变异度,进行PCA分析和(O)PLS-DA分析(Chenetal., 2009)。PCA分析结果如图2A所示,通过观察质控样品(mix1、mix2)之间的分散程度,可以确定仪器的稳定性和可靠性,PCA结果表明,三年桐、千年桐的根部代谢物的含量差异明显(图2A)。同时,(O)PLS-DA分析可以最大程度地提高组间差异,有助于寻找差异代谢物(Thévenotetal., 2015)。基于PLS-DA模型的得分图,进一步证明了三年桐、千年桐的根部代谢物含量差异明显(图2B)。
与三年桐相比,病原菌Fof-1侵染后千年桐根部有106种代谢产物含量上升(图2C)。这些含量上升代谢物包括黄酮类物质(黄酮、黄烷醇、异黄酮、黄烷酮、花青素、香豆素、香豆素和原花青素等32种化合物,占30.19%),氨基酸衍生物(9种化合物,占8.49%),核苷酸及其衍生物(8种化合物,7.55%),奎宁酸盐及其衍生物(7种化合物,6.60%),羟基肉桂酰基衍生物(7种化合物,6.60%)等29类化合物。结果可见,黄酮类物质占据最大比例,其中芒柄花苷(属于异黄酮)和橙皮苷(属于黄烷酮)的倍数变化分别达到2 026.26和1 185.52倍(表1)。进一步通过热图可以明显看出,106种物质在病原菌Fof-1侵染后的三年桐、千年桐根部含量有明显差异(图2D)。

图2 枯萎病菌Fof-1侵染后三年桐、千年桐根部代谢组学分析
2.3 千年桐根部苯丙烷类合成途径在Fof-1侵染后显著富集
利用Blast算法(E-value≤1e-5)在6个公共数据库(Nr,Pfam,KOG,Swiss-Prot,GO,KEGG)中,三年桐、千年桐共注释了258 435和245 240个转录本。根据RPKM值进行差异表达分析,三年桐、千年桐分别有20 031和6 366个差异表达基因。根据K均值聚类分析将这些差异表达基因分别划分为20个表达模式。
当植物受到病害胁迫发生生理和生化反应之前,首先是基因表达的启动。因此,病原菌侵染早期的基因应答特别重要。对病原菌侵染早期的三年桐和千年桐(F1,M1)上调差异表达的基因进行分析,发现在三年桐和千年桐根部分别有7 015和1 986个(图3A)。进一步KEGG通路富集分析表明,千年桐的1 986个差异表达基因显著富集在以下7个途径: 苯丙烷类生物合成,核糖体,其他聚糖降解,缬氨酸、亮氨酸和异亮氨酸降解,乙醛酸盐和二羧酸盐代谢,烟酸和烟酰胺代谢,半乳糖代谢(P<0.05)(图3B)。对于三年桐,7 015个差异表达基因显著富集在以下9个途径: 氧化磷酸化,代谢途径,碳代谢,精氨酸生物合成,蛋白酶体,柠檬酸盐循环,丙氨酸、天冬氨酸和谷氨酸代谢,乙醛酸和二羧酸代谢,氨基酸的生物合成(P<0.05)(图3B)。由以上结果分析,与抗病相关的苯丙烷类生物合成途径只在病原菌Fof-1侵染早期的千年桐根部发生了响应,在三年桐根部没有发生响应。

表1 千年桐根部Fof-1侵染后32种上调黄酮类化合物的信息

图3 枯萎病菌Fof-1侵染后三年桐和千年桐根部基因表达分析及通路富集分析
2.4 千年桐根部苯丙烷类生物合成途径在Fof-1侵染后上调响应
代谢组学分析表明,在Fof-1侵染后期,千年桐的106种上调代谢产物中,黄酮类物质占最大比例(32种化合物,占30.19%)。在大多数情况下,基因转录过程先于代谢过程发生。因此,在三年桐、千年桐的转录组学分析中,将研究重点放在侵染早期(F1,M1)。与代谢组学结果一致,在Fof-1侵染早期,千年桐根部差异表达基因也富集到黄酮类生物合成途径(P= 0.213 168)(图3B)。苯丙烷类生物合成是黄酮类生物合成途径的上游途径,并由此产生各种化合物,包括黄烷酮、花青素、异黄酮等等(https:∥www.kegg.jp/kegg/pathway.html),差异表达基因分析结果表明在侵染早期千年桐差异根部表达基因显著富集到苯丙烷生物合成途径中(P= 0.019 170 4)(图3B),但三年桐在侵染早期并未富集到上述途径。
在侵染早期,千年桐差异表达基因共注释到苯丙烷生物合成途径中的差异表达基因有30个,包括4-香豆酸: CoA连接酶(4-coumarate: CoA ligase,4CL),β-葡萄糖苷酶(β-glucosidase,BGL),咖啡酰-CoA O-甲基转移酶(caffeoyl-CoA O-methyltransferase,CCoAOMT),过氧化物酶(peroxidase,POX),反式肉桂酸4-单加氧酶(trans-cinnamate 4-monooxygenase)等编码基因(表2),千年桐中注释到黄酮类生物合成途径的差异表达基因有7个,包括黄酮类3-单加氢酶、咖啡酰-CoA O-甲基转移酶(caffeoyl-CoA O-methyltransferase,CCoAOMT)、反式肉桂酸4-单加氧酶、黄酮醇合酶(flavonol synthase,FLS)、柚皮素3-双加氧酶等,其中2个咖啡酰-CoA O-甲基转移酶基因、1个反式肉桂酸4-单加氧酶基因(comp158232_c0;comp158238_c0;comp162378_c0)也是上述苯丙烷生物合成途径中的基因。
2.5 千年桐根部苯丙烷生物合成途径在Fof-1侵染后的基因互作网络
利用WGCNA分析(加权截止值> 0.50),发现与苯丙烷合成途径相关的30个相关基因中,有4个是病原菌侵染后早期应答的中心基因,包括4-香豆酸: CoA连接酶(4CL-like 7)编码基因(comp164220_c0),β-D-木糖苷酶(Xyl,β-D-xylosidase)编码基因(comp163132_c0),β-葡萄糖苷酶41(BGL 41)编码基因(comp165691_c0)和过氧化物酶N1(POX N1)编码基因(comp115360_c0)。这4个中心基因与其他1 625个基因具有极高的相关性(图4A)。根据KEGG通路注释,将1 625个基因分为5类,包括代谢、遗传信息处理、环境信息处理、细胞处理和生物系统。其中,代谢途径占最大比例(图4B)。

表2 千年桐根部苯丙烷类生物合成途径和黄酮类生物合成途径的差异表达基因信息

图4 枯萎病菌Fof-1后千年桐根部苯丙烷类生物合成途径中的关键基因响应其他基因的网络分析
为探讨在枯萎病菌Fof-1侵染后,千年桐根部显著富集的苯丙烷类合成途径中4个中心基因的表达变化,进行了qRT-PCR分析验证。结果表明,千年桐根部4个中心基因CoA连接酶(4CL-like 7)编码基因(comp164220_c0)、β-D-木糖苷酶(Xyl)编码基因 (comp163132_c0)、β-葡萄糖苷酶41(BGL 41)编码基因 (comp165691_c0)和过氧化物酶N1(POX N1)编码基因 (comp115360_c0)均在病原菌Fof-1侵染早期表达量显著升高(图5)。这与转录组测序结果相一致(图5)。
3 讨论
3.1 黄酮类化合物与植物抗病的关系
由尖孢镰刀菌引起的土传真菌病害不仅对作物,对规模化种植的经济林木也造成了巨大的损失。油桐枯萎病对现有规模化种植的油桐林也造成了毁灭性破坏。三年桐是我国油桐主栽培品种,产量高、桐油品质好,但易感枯萎病。相对于三年桐,千年桐产量低、桐油品质次,但高抗枯萎病。千年桐抗枯萎病机制的研究对三年桐抗性育种有重要的指导意义。
植物自身产生的化合物对病原菌的防御起到了重要作用。苯丙烷及其衍生物是由维管植物产生的次生代谢产物,参与植物的许多生理活动,如生长、发育、繁殖和防御等,包括黄酮类化合物、简单酚类、醌类等。其中,黄酮类物质种类多、分布广,具有丰富的生物学活性,对多种病原菌具有防御作用,黄酮类生物合成途径基因的表达通过增加黄酮类产物从而增强植物对病原菌的抗性。本研究采用高效液相色谱和串联质谱检测方法对Fof-1侵染后的三年桐、千年桐根部样本进行代谢组学分析,并筛选出32个黄酮类化合物,其含量在千年桐中显著高于三年桐,初步推测这些黄酮类化合物可能在千年桐抗病中发挥作用。大量的研究表明,杨树(Populusalba)中黄酮类途径基因的表达可以增强杨树对病原菌的抗性(Baietal., 2020),黄酮类代谢相关基因的表达增强了大麦对禾谷镰刀菌(Fusariumgraminearum)的抗性(Karreetal., 2019),黄酮类途径代谢流的增强可以增加苜蓿(Medicagosativa)对病原菌镰刀菌(Fusariumoxysporumf. sp.medicaginis)的抗性(Gilletal., 2018)。炭疽菌(Colletotrichumsublineolum)侵染后高梁(Sorghum)会产生一种黄酮类物质发挥拮抗作用(Duetal., 2010); 从马铃薯(Solanumtuberosum)—洋葱中提取的黄酮类化合物对枯萎病菌萌发及其生长也有明显的抑制作用(Yangetal., 2019); 苜蓿通过产生一种异黄酮物质可以抵御锈病病原菌的侵染(Lietal., 2018)。

图5 千年桐根部苯丙烷合成途径4个中心基因响应Fof-1侵染的qRT-PCR验证
3.2 苯丙烷类和黄酮类生物合成途径相关基因在抗病中的重要作用
在病原菌侵染早期,千年桐中共注释到苯丙烷类和黄酮类生物合成途径中的差异上调基因共有34个,其中许多基因已与植物抗病相关,如白云杉(Piceaglauca)β-葡萄糖苷酶(BGL)已被证实可以增强植物对云杉芽虫(Choristoneurafumiferana)的天然抗性(Mageroyetal., 2015); 咖啡酰-CoA O-甲基转移酶(CCoAOMT)是植物中木质素生物合成途径的关键酶,在马尾松(Pinusradiata)中CCoAOMT可以调节木质素的组成和含量(Wagneretal., 2011),木质素含量的增加具有抗生物胁迫和非生物胁迫的作用。植物过氧化物酶(POX)或其同工酶可以通过催化合成杀菌物质、促进木质素合成、参与乳突形成和颗粒状沉积物的积累而构成植物的抗病性,如马铃薯中POX可以增强对疫霉(Phytophthorainfestans)的抗性(Yangetal., 2020)、第3类过氧化物酶PRX34在拟南芥中表达增强了对细菌和真菌病原体的抗性(Zhaoetal., 2019)。丝氨酸羧肽酶(SCPs)具有酰基转移酶功能,玉米(Zeamays)丝氨酸羧肽酶基因ZmSCP可以增强对水稻纹枯病菌(Rhizoctoniasolani)的抗性(Liuetal., 2013)。
千年桐根部在枯萎病病原菌Fof-1侵染后,黄酮类化合物含量增加,响应的苯丙烷类生物合成途径和黄酮类类生物合成途径也显著富集,上述途径中4个中心基因在病原菌Fof-1侵染后也显著上调。这4个中心基因中,4-香豆酸: CoA连接酶-7(4CL)是苯丙烷类生物合成途径的限速酶,位于黄酮类化合物代谢途径的重要分支或生物合成途径的下游(Lietal., 2014),与黄酮和木质素的合成密切相关(Chenetal., 2019)。据文献报道,香鳞毛蕨(Dryopterisfragrans)4CL基因的表达可以提高烟草(Nicotianatabacum)中黄酮类化合物的含量(Lietal., 2020); 水曲柳(Fraxinusmandshurica)4CL基因可以通过增加松柏醇含量提高烟草的耐旱性和渗透胁迫能力(Chenetal., 2020)。中心基因编码蛋白β-D-木糖苷酶(Xyl)是1个多功能蛋白,对根尖细胞的细胞壁结构有重要调控作用(Xiongetal., 2007)。β-葡萄糖苷酶(BGL)编码基因的表达直接影响银杏(Ginkgobiloba)中黄酮类化合物的含量(Fangetal., 2019)。由此推测,在病原菌Fof-1侵染后,千年桐根部通过4-香豆酸: CoA连接酶-7(4CL)等中心基因的启动,激发苯丙烷类生物合成途径及黄酮类生物合成途径,响应病原菌入侵,产生更多的黄酮类物质,包括芒柄花苷、橙皮苷等异黄酮和黄烷酮化合物,起到对枯萎病病原菌Fof-1的抵御作用。
3.3 展望
本研究着重讨论病原菌Fof-1侵染后千年桐根部黄酮类化合物及其所在的苯丙烷类合成途径在抗枯萎病中的作用,而病原菌Fof-1侵染后千年桐根部其他上调化合物,如儿茶素、茉莉酸等也可能在千年桐抗枯萎病中发挥作用。儿茶素合成相关基因DFR受杨树(Populusspp.)细菌性溃疡病菌(Lonsdaleaquercina)的诱导而表达上调并发挥抗病作用(Houetal., 2016); 苹果(Malussieversii)褐斑病菌(Diplocarponmali)的入侵会引起表儿茶素含量的显著升高,且在抗病苹果中的上升程度要高于感病苹果,说明表儿茶素可能与苹果抗褐斑病密切相关(Yinetal., 2013a)。此外,病菌入侵后茉莉酸途径关键基因COII和PLD在抗病苹果中呈现上调表达,外源施加茉莉酸可以提高感病苹果对褐斑病菌的抗性(Yinetal., 2013b); 水杨酸、茉莉酸和脱落酸信号转导途径参与了鹰嘴豆(Cicerarietinum)对枯萎病菌(Fusariumoxysporumf. sp.ciceri)的抗性反应,并可能激发其他信号通路(Gayatridevietal., 2012)。在番茄(Solanumlycopersicum)中茉莉酸甲酯通过水杨酸和黄酮醇的积累诱导了种子对半活体营养番茄专化型尖孢镰刀菌(hemi-biotrophFusariumoxysporumf.sp.lycopersici)的抗性(Krletal., 2015)。
因此,在后续的研究中,应重点关注儿茶素、茉莉酸等化合物在千年桐抗枯萎病中的作用以及这些物质和黄酮类化合物之间是否有协同作用,进一步解析千年桐的抗枯萎病机制。
4 结论
千年桐根部提取物与三年桐根部提取物相比较,对油桐枯萎病病原菌Fof-1的生长具有明显的抑制作用。通过代谢组分析发现,与三年桐相比,千年桐根部黄酮类化合物含量增高,其中芒柄花苷、橙皮苷等异黄酮和黄烷酮化合物是三年桐的1 000倍以上。进一步通过比较转录组分析,千年桐根部负责黄酮类化合物合成的上游关键信号通路“苯丙烷类生物合成途径”在病原菌Fof-1侵染后比三年桐显著富集。“苯丙烷类生物合成途径”中4-香豆酸CoA连接酶(4CL-like 7)、β-D-木糖苷酶(Xyl)、β-葡萄糖苷酶41(BGL 41)和过氧化物酶N1(POX N1)编码的4个中心基因经qRT-PCR验证在枯萎病病原菌Fof-1侵染早期上调表达,与转录组结果一致。基于代谢组和转录组的联合分析,表明千年桐在枯萎病病原菌侵染后,苯丙烷生物合成途径发生应答响应,在千年桐根部产生异黄酮和黄烷酮等黄酮类化合物,发挥抵御病原菌入侵的作用。
猜你喜欢
热带农业科学(2022年4期)2022-11-23
食品工业(2022年5期)2022-06-13
天津农业科学(2022年5期)2022-05-31
中国药房(2022年10期)2022-05-30
中国瓜菜(2022年3期)2022-05-05
科学导报·学术(2020年55期)2020-02-22
分析化学(2018年4期)2018-11-02
成长·读写月刊(2017年7期)2017-07-13
中国医药科学(2016年5期)2016-10-08
现代农业科技(2009年9期)2009-02-21
