
常温和低温条件下不同浓度外源褪黑素对降香黄檀幼苗的生理生态影响
2022-05-13 08:24蒲玉瑾张一璇苗灵凤杨帆
广西植物 2022年4期
蒲玉瑾 张一璇 苗灵凤 杨帆






摘要: 為探究外源褪黑素对不同温度下降香黄檀幼苗的生理生态影响,该研究开展了不同浓度外源褪黑素(300、500、600、700、900、1 200 μmol·L)对常温(白天28 ℃/夜晚25 ℃)和低温(白天8 ℃/夜晚5 ℃)下降香黄檀(Dalbergia odorifera)幼苗的生长发育、光合作用、叶片色素含量、叶片水分状况、膜系统等方面影响研究,并通过隶属函数分析综合评价筛选出具有促进降香黄檀幼苗生长发育和提高低温耐受性的最适外源褪黑素浓度。结果表明:(1)在常温下喷施各个浓度的褪黑素溶液,在生长发育(形态、茎高增长和株高增长),光合作用参数[净光合速率(P)、气孔导度(G)、胞间二氧化碳浓度(C)和蒸腾速率(T)],光合色素[总叶绿素(Total Chl)、叶绿素 a(Chl a)、叶绿素 b(Chl b)和类胡萝卜素(Caro)]和可溶性蛋白含量上均有一定的提高。(2)低温胁迫处理下,喷施各个浓度的褪黑素溶液可在不同程度上缓解低温胁迫对植株形态、茎高增长、株高增长、P、C、T、Total Chl、Chl a、Caro、露点水势(DWP)、相对电导率(RC)、丙二醛(MDA)和可溶性蛋白含量等方面的负面影响。(3)隶属函数分析表明, 600~700 μmol·L外源褪黑素处理促进降香黄檀幼苗生长和缓解低温冷害效果最佳。该研究结果为提高降香黄檀幼苗的低温耐受能力提供了科学依据,有利于降香黄檀在亚热带低温地区的栽培管理。
关键词: 低温胁迫, 光合特性, 降香黄檀, 生理生态, 褪黑素
中图分类号: Q945.3文献标识码: A文章编号: 1000-3142(2022)04-0648-11
Effects of different exogenous melatonin concentrations on eco-physiological characteristics of Dalbergia odorifera seedlings under ambient and low temperatures
PU Yujin ZHANG Yixuan MIAO Lingfeng YANG Fan
( 1. College of Ecology and Environment, Hainan University, Haikou 570228, China;
2. Center for Eco-Environmental Restoration
Engineering of Hainan Province, Haikou 570228, China; 3. Key Laboratory of Agro-Forestry Environmental
Processes and Ecological Regulation of Hainan Province, Haikou 570228, China )
Abstract: The purpose of this study was to explore the eco-physiological effects of different exogenous melatonin concentrations on the Dalbergia odorifera seedlings under different temperatures. The effects of different exogenous melatonin concentrations (300, 500, 600, 700, 900, 1 200 μmol·L) on the eco-physiological traits in term of plant growth, photosynthesis, leaf pigment content, leaf water status, membrane system in D. odorifera seedlings under ambient temperature (28 ℃ day / 25 ℃ night) and low temperature (8 ℃ day /5 ℃ night) were studied. The optimum concentration of melatonin, which could promote the growth and development and enhance the low temperature tolerance, was screened out through the comprehensive evaluation of subordinate function analysis. The results were as follows: (1) Application of all exogenous melatonin concentrations could increase the growth and development of D. odorifera (morphology, stem height and plant height), the photosynthetic parameters (Pn, Gs, Ci, and Tr), the pigment contents (Total Chl, Chl a, Chl b, and Caro), and soluble protein content under ambient temperature. (2) Under low temperature stress, spraying different concentrations of melatonin solution could alleviate the negative impacts caused by low temperature on morphology, stem height increment and plant height increment, Pn, Ci, Tr, Chl, Chl a, Caro, dew point water potential (DWP) and relative conductivity, malonaldehyde and soluble protein contents. (3) The results of subordinate function analysis showed that 600-700 μmol·L melatonin had the best effects on promoting growth and development and alleviating chilling damages. This study provides scientific guidance for coping with chilling stress on D. odorifera seedlings and popularizing its cultivation in low temperature regions of subtropical areas.
Key words: low temperature stress, photosynthetic characters, Dalbergia odorifera, eco-physiology, melatonin
目前,我國红木资源短缺,98%以上的红木木材依赖进口。因此,广泛培育红木森林资源、扩大栽培范围成了国家林业重大发展战略之一。降香黄檀(Dalbergia odorifera)是豆科蝶形花亚科黄檀属常绿半落叶乔木,别名黄花梨、降香、花梨、香红木、香枝木、花梨木、海南檀、花梨母等,是海南省特有的珍贵红木树种,被列为国家二级保护植物,为国家标准5属8类34种红木之一(贾瑞丰等, 2013; 杨冬华等, 2013)。降香黄檀原产于中国海南省,由于引种栽培,因此广西(梁建平等, 2015)、广东(连辉明等, 2014)、福建(叶水西, 2008;王志良等, 2015)等地现也有分布。但是,在这些地区引种栽培的降香黄檀经常受到不同程度的低温伤害。例如,在广东、福建引种栽培的降香黄檀均受到不同程度的寒害和冻害(叶水西, 2008;连辉明等, 2014)。在自然生长条件无法改变的情况下,通过外源物质来增强植物自身对逆境的抗性,可以提高植物适应不利生长环境的能力,平稳渡过极端环境条件。目前,由于利用外源物质来改善降香黄檀对环境胁迫耐受性的研究报道较少。因此,急需开展相关研究来合理科学地指导降香黄檀在亚热带地区的引种栽培和人工林培育。
相关研究表明,外源褪黑素可以通过提高抗氧化机制的效率来增加植物对环境胁迫的抗逆性,其机制主要通过直接对活性氧进行清除;刺激抗氧化酶活性进行抗氧化调节;增强其他抗氧化剂的抗氧化能力;降低电子渗透率,提高线粒体内部氧化磷酸化进度进行抗氧化调节等4种方式来进行调节,外源褪黑素可以缓解叶绿素降解、降低膜脂过氧化程度、调节内源激素及多胺含量来提高植株抗环境胁迫的能力(Zhang et al., 2012;Shi et al., 2014;Turk et al., 2014)。低温下,外源褪黑素可通过提高黄瓜(Cucumis sativus)幼苗、番茄(Solanum lycopersicum)幼苗、小麦(Triticum aestivum)幼苗、狗牙根(Cynodon dactylon)幼苗、菘蓝(Isatis indigotica)种子中抗氧化酶活性;增加渗透保护物质如碳水化合物、脯氨酸含量;大幅降低MDA 和 H2O2含量来保持细胞膜的完整性和功能;减小了低温对植物细胞的损伤,有效地提高了植物抗低温能力(Posmyk et al., 2009; 包宇等, 2013; 潘红艳等, 2013; Turk et al., 2014; 高青海等, 2014; Fan et al., 2015; Liu et al., 2015; Hu et al., 2016)。这些研究表明外源褪黑素对草本植物确实起到提高胁迫适应能力的作用,但对木本植物的研究未见报道。另外,针对不同的草本植物,应用外源褪黑素的浓度也不尽相同,有的甚至差别较大。因此,筛选具有促进降香黄檀幼苗生长发育和缓解低温胁迫作用的最适褪黑素施用浓度,并揭示其对降香黄檀幼苗的生理生态影响机制,对降香黄檀幼苗在亚热带地区的引种栽培具有重要意义。
本文以降香黄檀幼苗为研究对象,从植株生长发育、叶片水分状况、叶片色素含量、光合作用及膜系统参数等角度,探讨不同浓度的外源褪黑素对不同温度下降香黄檀幼苗的生理生态的影响,通过隶属函数分析综合评价,筛选出具有促进降香黄檀幼苗生长发育和缓解低温胁迫作用的最适褪黑素浓度,并探讨其影响机制。
1材料与方法
1.1 实验材料
以从海南尖峰岭购买的2017年的降香黄檀一年实生苗为研究材料,当年12月底将幼苗植于塑料花盆中(口径 21 cm × 19 cm 高)进行培养,每盆1株。培养基质为红壤土和椰糠(V/V, 3∶2),每盆5.0 kg。实验前测得土样基本理化值:最大田间持水量52.99%、全氮1.77 g·kg、全磷0.64 g·kg、有机质58.01 g·kg、有机碳33.65 g·kg、pH 值6.23。实验材料置于海南大学海甸校区大棚温室,生长期间温室平均温度和湿度分别为19.8 ℃和73%。为了获得长势一致的植株,统一在距土壤5 cm处剪去地上部分,等待重新萌发新芽。在2018年3月初,待植株平均株高达到20 cm后,选取长势基本一致的健康幼苗进行褪黑素处理和低温实验。
1.2 实验方法
采用褪黑素和温度两因素进行完全随机实验设计。褪黑素浓度设7个水平(0、300、500、600、700、900、1 200 μmol·L),褪黑素处理为每3 d喷施一次褪黑素溶液,喷施于叶片正反面至滴水为准。温度设2个水平,正常温度设置为白天28 ℃(8:00—20:00),晚上25 ℃(20:00—8:00);低温设置为白天 8 ℃(8:00—20:00),晚上5 ℃(20:00—8:00)。采用人工气候箱(QHX-III, 上海新苗医疗器械医疗器械制造有限公司, 中国)来控制温度。实验处理期间每天采用自来水浇透至饱和,托盘渗液重新浇回盆中的方式来保持土壤水分含量一致。植株从大棚转入培养箱中开始进行褪黑素处理,在常温下适应2 d后,低温处理组的植株在10 ℃适应7 d后,再转入5 ℃的低温处理30 d。因此,褪黑素处理时间为39 d,低温处理持续时间为30 d。该实验设计包括了常温、常温+褪黑素(6个梯度)、低温、低温+褪黑素(6个梯度)共14个处理,每个处理4个重复,每个重复5株幼苗。
1.3 测量指标
1.3.1 生长指标降香黄檀的叶子属于羽状复叶,羽状复叶的生长速度和茎的生长速度有时会出现较大的差异,如图1所示。因此,在统计降香黄檀幼苗生长发育情况时,需要同时测量茎高与株高(直尺测量,精确至0.1 cm),方能较为全面地表示降香黄檀幼苗的生长情况。处理前和结束后各测量一次,计算净增长值。
1.3.2 生理指标参考Yang & Miao(2010)的方法,采用LI-6400光合仪(LI-6400XT, Gene Company Limited, USA)测量植株光合指标,使用2 cm × 3 cm红蓝光源叶室,光源强度为1 200 μmol·m·s-1,测量参数包括净光合速率(Pn, μmol·m·s)、气孔导度(Gs, mol·m·s)、胞间二氧化碳浓度(Ci, μmol·mol)、蒸腾速率(Tr, mmol·m·s)。
光合色素含量的测定参考Yang & Miao(2010)的方法,首先用乙醇法提取,然后用分光光度计(UV-1800PC,上海美谱达仪器有限公司,中国)测量470、649、665 nm 3个波长的吸光度值,最后分别计算出总叶绿素含量(Total Chl, mg·g)、叶绿素a(Chl a, mg·g)、叶绿素b(Chl b, mg·g)及类胡萝卜素含量(Caro, mg·g)。
叶片相对含水量的测量参考Yang & Miao(2010)的方法。从每株幼苗上剪取成熟的功能叶片2~3片,每个重复取1个样。相对含水量(RWC,%)=(F-D)/ F × 100。式中:F为叶片鲜重;D为叶片干重。
叶片水势的测量参考苗灵凤等(2017)的方法。首先在9:00—10:00采下健康成熟叶片后立即置于冰壶内带回实验室,然后将叶片剪成直径3 cm 的整圆形叶片平铺至WP4C露点水势仪样品杯底部,利用WP4C露点水势仪(WP4C, Gene Company Limited, USA)测定叶片水势。
相对电导率测量参考Shi等(2014)的方法。使用电导率仪(FE38,METTLER TOLEDO, USA)测量,相对电导率(RC,%)=C/C × 100。式中:C为鲜叶浸泡2 h的电导率;C为叶片煮沸30 min后的电导率。
丙二醛(MDA)含量和可溶性蛋白的测量参考Han等(2015)的方法,利用分光光度计测定。
1.4 数据分析
采用单因素方差分析(ANOVA)分析同一温度下不同褪黑素浓度之间的显著性差异,采用配对样本t检验分析同一褪黑素浓度下不同温度之间的显著性差异(P<0.05)。
使用隶属函数对降香黄檀幼苗各项指标进行综合分析,综合评价褪黑素浓度对不同温度下降香黄檀幼苗的生理生态影响。参照刘建新等(2018)的方法进行分析。
U(X)=(X-X)/(X-X)。
式中:U(X)代表隶属函数值;X代表幼苗的某个测定指标;X代表该指标的最大值;X代表该指标的最小值。特别需要注意的是,在各个指标中,若该指标的值与降香黄檀的生长以及低温抗性呈现负相关,则公式中X换成X,X换成X,其余的不變。
2结果与分析
2.1 植株形态变化
常温和低温下不同浓度梯度褪黑素溶液对降香黄檀幼苗在生长发育方面均有不同程度的促进效果。在常温下,各个浓度褪黑素处理的长势均比不加褪黑素的对照处理好,其中500、600、700 μmol·L褪黑素处理的植株长势优势明显。低温胁迫阻碍了降香黄檀幼苗的生长发育,使植株生长缓慢,叶片变黄萎蔫。与不加褪黑素的低温对照处理相比,添加外源褪黑素可以缓解低温胁迫作用,其中经过600、700 μmol·L外源褪黑素处理后的降香黄檀苗长势相对较好,缓解低温胁迫作用效果最明显,而其余处理的低温胁迫缓解能力有限(图2)。
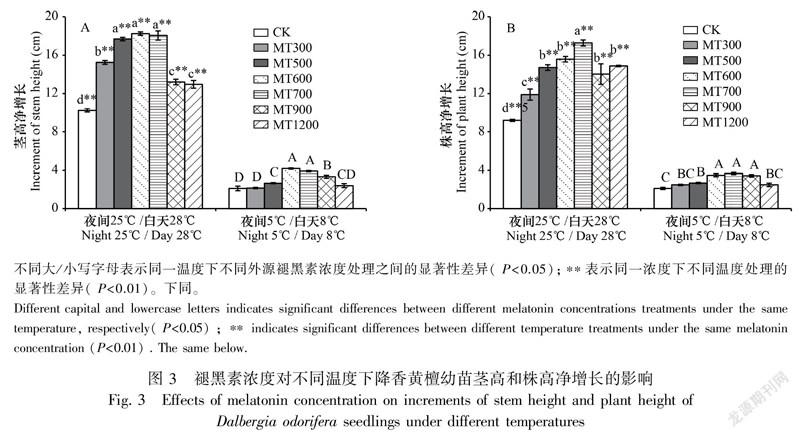
2.2 茎高和株高
常温下外源褪黑素对降香黄檀幼苗茎高、株高增长量的影响趋势为随褪黑素浓度升高呈现先上升后下降的趋势,均显著高于不加褪黑素的对照处理(P<0.05)。500、600、700 μmol·L的外源褪黑素处理对降香黄檀幼苗茎高增长效果最为显著,且这3个浓度间无显著差异(图3: A);700 μmol·L的外源褪黑素处理对株高增长作用最为显著,500、600、900、1 200 μmol·L 4个褪黑素浓度处理的增长作用差异不显著(图3: B)。
在低温胁迫下,降香黄檀幼苗茎高和株高的增长均受到了显著的抑制,施加外源褪黑素可在一定程度上缓解低温对茎、株增长的抑制作用。600、700 μmol·L的外源褪黑素处理植株的茎高增长量显著高于其他褪黑素浓度效果,其中600 μmol·L处理的效果最佳(图3: A);600、700、900 μmol·L的外源褪黑素处理植株的株高增长量显著高于其他浓度的效果,这3个浓度间差异不显著(图3: B)。其余浓度的外源褪黑素处理对缓解低温胁迫对降香黄檀幼苗径高、株高抑制作用的效果并不明显。
2.3 光合作用参数
如图4所示,常温下对降香黄檀幼苗施加各个浓度的外源褪黑素,P、G、C和T随着褪黑素浓度的升高呈现先上升后下降的趋势。P、G和T都在600 μmol·L处理促进作用达到最高值,其余浓度处理的促进作用向600 μmol·L两侧依次减弱;C添加外源褪黑素显著高于不加褪黑素的对照,500、600 μmol·L处理促进作用最为明显,300、900、1 200 μmol·L处理促进作用依次降低。低温对降香黄檀幼苗光合作用造成胁迫危害,施加外源褪黑素可缓解低温对光合作用的抑制,P、G、C和T随褪黑素浓度升高呈现先上升后下降的趋势。除1 200 μmol·L处理外,其他浓度褪黑素处理的P均显著高于不加褪黑素的低温对照, 其中600 μmol·L处理效果最佳;G,T和C在900 μmol·L处理抵御低温胁迫能力最为明显,其余处理与不加褪黑素的低温对照相比,抵御低温胁迫的能力差异不显著。
2.4 叶片色素含量
如图5所示,常温下对降香黄檀幼苗施加各个浓度的外源褪黑素,可促进Total Chl、Chl a、Chl b和Caro的合成,光合色素含量都随着褪黑素浓度升高呈现先上升后下降的趋势。添加外源褪黑素,除300 μmol·L处理外,其他浓度褪黑素处理的Total Chl、Chl a、Chl b和Caro均高于不加褪黑素的对照,并在600 μmol·L处理含量达到最大值,600 μmol·L-1处理促进光合色素合成的效果最为显著(图5);Chl a在1 200 μmol·L处理,Chl b在 700 μmol·L处理,Total Chl 在700、1 200 μmol·L处理均显著高于不添加褪黑素的对照,其他浓度褪黑素处理与不加褪黑素的对照无明显差异(图5:A, B, C)。受低温胁迫的影响,降香黄檀幼苗光合色素含量显著降低,施加外源褪黑素在一定程度上可以缓解低温胁迫对光合色素合成的阻碍。Total Chl、Chl a和Caro随褪黑素浓度升高呈现先上升后下降的趋势,低温胁迫缓解效果较好的为500、600、700 μmol·L处理,其中,Chl a在900、1 200 μmol·L处理与不加褪黑素的对照差异显著,Chl b在300、500、900、1 200 μmol·L处理与不加褪黑素的对照差异显著,Total Chl和Caro在300、900、1 200 μmol·L处理与不加褪黑素的对照差异显著,这些浓度的褪黑素处理对低温胁迫抑制光合色素合成的缓解能力有限(图5)。
2.5 叶片相对含水量和露点水势
如图6所示,常温下对降香黄檀幼苗施加各个浓度的外源褪黑素,随着褪黑素浓度增加,叶片相对含水量表现为先上升后下降的趋势,在600 μmol·L处理达到最高值,但均与不加褪黑素的对照无显著差异(图6: A);露点水势也随着褪黑素浓度增加表现先增大后减小的趋势,在600、700 μmol·L达到较高值,且均显著低于不加褪黑素的对照(图6: B)。低温胁迫使降香黄檀幼苗叶片的相对含水量和露点水势显著降低,施加外源褪黑素可缓解低溫胁迫对相对含水量和露点水势产生的负面影响。相对含水量在600、700 μmol·L处理对低温胁迫的抵御效果稍优于其他处理,但均与不加褪黑素的低温对照不存在显著差异(图6: A);露点水势在1 200 μmol·L处理的抵御低温胁迫效果最显著,600、700、900 μmol·L褪黑素浓度处理抵御能力次之,且均与不加褪黑素的低温对照差异显著(图6: B)。
2.6 叶片抗性生理生化指标
如图7所示,1 200 μmol·L浓度的外源褪黑素处理,相对电导率和MDA水平都显著高于不加褪黑素的对照处理。只有在600 μmol·L的外源褪黑素处理中的相对电导率低于对照处理;其余处理中的相对电导率和MDA水平都与对照处理无显著差异(图7: A, B)。可溶性蛋白含量随着褪黑素浓度的增加呈先上升后下降的趋势,600、700 μmol·L处理的含量显著高于不加褪黑素的对照,且在600 μmol·L处理达到最大值(图7: C)。低温对降香黄檀幼苗相对电导率、MDA和可溶性蛋白含量存在显著的影响,施加外源褪黑素可降低相对电导率和MDA的增幅,且随着褪黑素浓度升高呈现先降低后上升的趋势。在600 μmol·L处理中的相对电导率显著低于其他处理,600 μmol·L处理效果最佳;添加外源褪黑素处理的MDA水平显著低于不加褪黑素的低温对照,600 μmol·L处理低于其他处理组,但各个褪黑素浓度处理之间不存在显著差异;施加外源褪黑素可增加可溶性蛋白的累积,含量上升,随着褪黑素浓度增加呈现先增加后下降的趋势,在600、700 μmol·L处理累积含量显著,其中600 μmol·L处理的抗寒性最强,但两个浓度间不存在显著差异。
2.7 隶属函数分析
通过对各项生长及生理指标的隶属函数计算分析,综合评价褪黑素浓度在常温和低温下对降香黄檀幼苗生长和低温耐受性的影响。如表1所示,常温下褪黑素各个浓度处理的隶属函数值从大到小依次为MT600>MT700>MT500>MT900>MT300>CK>MT1200,常温下促进降香黄檀幼苗生长效果最好的褪黑素处理浓度为600 μmol·L;低温下褪黑素各个浓度处理的隶属函数值从大到小依次为MT600>MT700>MT900>MT500>MT1200>MT300>CK,低温下缓解低温胁迫作用最好的褪黑素浓度仍然是600 μmol·L。
3讨论与结论
褪黑素广泛存在于植物体内,其含量水平与植物生长发育各个阶段具有一定的关系(Shi et al., 2014; Zhang et al., 2015),这说明褪黑素浓度水平会影响植物的生长发育。但是,针对不同植物,最适的褪黑素浓度差异较大。因此,从生理生态适应的角度筛选促进降香黄檀幼苗生长发育的最适浓度,并阐明其生理生态调控机制具有重要意义。比如,有关对黄化的羽扇豆(Lupinus micranthus)的研究发现,胚轴浸入不同浓度梯度的褪黑素溶液中,低浓度的褪黑素均可以对羽扇豆下胚轴的伸长起到有利的作用,而高浓度的褪黑素则抑制生长(Hernandez et al., 2004);当褪黑素浓度小于等于10 μmol·L时对玉米蔗糖合成、水解基因表达、酶活性和蔗糖代谢具有积极的影响,当浓度大于等于100 μmol·L则造成负面影响(Zhao et al., 2015);对黄大豆(Glycine max)施加0~100 μmol·L的褪黑素的研究发现,50 μmol·L的浓度下长势最好,叶面积最大(Wei et al., 2015)。本研究结果表明,常温下对降香黄檀幼苗喷施各个浓度的褪黑素溶液对降香黄檀幼苗虽然均有由一定程度的促进作用,但浓度过低或过高效果都会受到限制;只有在浓度为600~700 μmol·L时才能在植株形态发育、茎高增长和株高增长、光合作用色素和气体交换等多个方面起到最大的生理生态调节作用。另外,通过比较分析发现,外源褪黑素在木本植物应用的最适浓度大大高于在草本植物上应用的最适浓度。
低温是影响热带多年生植物生长发育及其地理分布的主要环境限制因素。当植物遇到低温胁迫时,植物体内会发生一系列的生理生态变化。低温条件下,植物在形态上表现为叶片失水萎蔫、皱褶或出现脱水坏死斑;植株株高、叶片及根系正常生长发育受阻、甚至植株死亡(包宇等, 2013; 高青海等, 2014; Hu et al., 2016)。前人研究表明,施加外源褪黑素可有效缓解低温对草本植物的伤害,尤其是缓解低温对生长发育、光合色素、光合作用及膜稳定性等方面的负面影响(Shi et al., 2014; Turk et al., 2014)。比如,与未加褪黑素处理相比,对低温下的狗牙根添加外源褪黑素后,存活率显著提高,植株长势较好,绿叶数量较多、叶绿素含量较高,且相对电导率和 MDA 含量显著降低(Shi et al., 2014)。本研究结果表明,低温下对降香黄檀施加600~700 μmol·L浓度的外源褪黑素,能够最大限度地缓解低温对降香黄檀幼苗生长的抑制,并增强叶绿素稳定性,提高光合速率。低温下对草本植物施加褪黑素能够改变细胞膜的流动性,同时提高了抗氧化酶的活性,增强清除活性氧簇物质的能力,从而缓解低温对植物造成的伤害(Bajwa et al., 2014; Shi et al., 2014; Zhang et al., 2015; Hu et al., 2016)。本研究中,低温胁迫下喷施600~700 μmol·L浓度的外源褪黑素可使降香黄檀幼苗的相对电导率、MDA和可溶性蛋白含量受低温胁迫危害得到有效缓解,浓度过低或过高缓解效果都会受到影响。另外,通过比较分析发现,外源褪黑素调节草本植物和木本植物对低温胁迫的生理生态适应机制虽有类似之处,但需注意最适浓度的筛选。
在常温下喷施各个浓度的褪黑素溶液,降香黄檀幼苗在生长发育,光合作用参数(Pn、Gs、Ci和Tr),光合色素(Total Chl、Chl a、Chl b和Caro)和可溶性蛋白含量均有一定的提高;低温胁迫下喷施各个浓度的褪黑素可在不同程度上缓解抑制作用,在生长发育、光合作用、光合色素、DWP、RC、MDA和可溶性蛋白含量等方面降香黄檀受低温胁迫的负面影响均能得到有效缓解。但是,褪黑素浓度过低或过高对常温下的促进效果和低温下的缓解负面作用都会受到影响。隶属函数分析表明,无论是在常溫还是低温下,600~700 μmol·L浓度的外源褪黑素处理对降香黄檀的生理生态影响效果最佳。本研究成果为提高降香黄檀的低温耐受能力提供了科学依据,有利于降香黄檀在亚热带低温地区的栽培管理。
参考文献:
BAO Y, LUO QX, HUANG J, et al., 2013. On effect of exogenous melatonin on physiological indexes of tomato seedling under low temperature stress [J]. J SW Chin Univ (Nat Sci Ed) , 38(10): 57-61. [包宇, 罗庆熙, 黄娟, 等, 2013. 外源褪黑素对低温胁迫下番茄幼苗生理指标的影响 [J]. 西南师范大学学报(自然科学版), 38(10): 57-61.]
GAO QH, WANG YK, LU XM, et al., 2014. Effects of exogenous melatonin on growth and antioxidant system of leaves in cucumber seedlings under low temperature and weak light stress [J]. Acta Bot Boreal-Occident Sin, 34(8): 1608-1613. [高青海, 王亚坤, 陆晓民, 等, 2014. 低温弱光下外源褪黑素对黄瓜幼苗生长及抗氧化系统的影响 [J]. 西北植物学报, 34(8): 1608-1613.]
HAN CY, CHAN ZL, YANG F, 2015. Comparative analyses of universal extraction buffers for assay of stress related biochemical and physiological parameters [J]. Prep Biochem Biotechnol, 45(7): 684-695.
HERNÁNDEZ-RUIZ J, CANO A, ARNAO MB, 2004. Melatonin: a growth-stimulating compound present in lupin tissues [J]. Planta, 220(1): 140-144.
HU ZR, FAN JB, XIE Y, et al., 2016. Comparative photosynthetic and metabolic analyses reveal mechanism of improved cold stress tolerance in bermudagrass by exogenous melatonin [J]. Plant Physiol Biochem, 100(1): 94-104.
JIA RF, YANG ZJ, XU DP, et al., 2013. Growth and endogenous hormone content of Dalbergia odorifera seedlings under drought stress [J]. Ecol Environ Sci, 22(7): 1136-1140. [贾瑞丰, 杨曾奖, 徐大平, 等, 2013. 干旱胁迫对降香黄檀幼苗生长及内源激素含量的影响 [J]. 生态环境学报, 22(7): 1136-1140.]
LIAN HM, ZHANG Q, YIN ZY, et al., 2014. Survey and analysis on growth and chilling injury of Dalbergia odorifera plantation in Guangdong Province [J]. J Cent S Univ For Technol, 34(10): 26-31. [連辉明, 张谦, 殷祚云, 等, 2014. 广东降香黄檀人工林生长及寒害调查分析 [J]. 中南林业科技大学学报, 34(10): 26-31.]
LIANG JP, JIANG JL, QIN WM, et al., 2015. Growth rule for a 46-year-old Dalbergia odorifera in Nanning, Guangxi [J]. J Zhejiang A & F Univ, 32(4): 523-528. [梁建平, 蒋军林, 秦武明, 等, 2015. 广西南宁46年生降香黄檀人工林生长规律 [J]. 浙江农林大学学报, 32(4): 523-528.]
LIU JX, OU XB, LIU XL, et al., 2018. Principal component and subordinate function of the alleviating effects of hydrogen peroxide (H2O2) on low-temperature stress in naked oat (Avena nuda) seedlings [J]. Bull Bot Res, 38(5): 748-756. [刘建新, 欧晓彬, 刘秀丽, 等, 2018. 过氧化氢缓解裸燕麦幼苗低温胁迫的主成分和隶属函数分析 [J]. 植物研究, 38(5): 748-756.]
LIU JL, WANG WX, WANG LY, et al., 2015. Exogenous melatonin improves seedling health index and drought tolerance in tomato [J]. Plant Growth Regul, 77(3): 1-10.
MIAO LF, ZHANG LJ, PU YJ, et al., 2017. Factors that affect the measurement using WP4C dewpoint potential meter to determine water potential — illustrated by the case of Dalbergia odorifera [J]. Plant Sci J, 35(1): 93-98. [苗灵凤, 张丽佳, 蒲玉瑾, 等, 2017. WP4C露点水势仪测定植物叶片水势时的影响因素——以降香黄檀为例 [J]. 植物科学学报, 35(1): 93-98.]
PAN HY, ZHANG XQ, LI J, et al., 2013. Effects of exogenous melatonin on antioxidant activities in Isatis indigotica Fort. seedlings after low temperature stress [J]. J NW Univ (Nat Sci Ed), 43(2): 238-242. [潘红艳, 张晓庆, 李婕, 等, 2013. 褪黑素对低温胁迫后菘蓝种子苗抗氧化性影响 [J]. 西北大学学报(自然科学版), 43(2): 238-242.]
POSMYK MM, BAABUSTA M, WIECZOREK M, et al., 2009. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress [J]. J Pineal Res, 46(2): 214-223.
SHI HT, YE TT, ZHONG B, et al., 2014. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (Cynodon dactylon (L.) Pers.) by exogenous calcium [J]. J Integr Plant Biol, 56(11): 1064-1079.
TURK H, ERDAL S, GENISELM, et al., 2014. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings [J]. Plant Growth Regul, 74(2): 139-152.
WANG ZL, 2015. Survey and analysis on growth of Dalbergia odorifera plantation in Fujian Province [J]. Low Carbon World, 21(1): 290-291. [王志良, 2015. 福建降香黃檀人工林生长调查分析 [J]. 低碳世界, 21(1): 290-291.]
WEI W, LI QT, CHU YN, et al., 2015. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants [J]. J Exp Bot, 66(3): 695-707.
WU XX, ZHU ZW, ZHAGN AD, et al., 2017. Effects of exogenous melatonin the growth, photosynthesis and antioxidant system in eggplant (Solanum melongena L.) seedlings under low temperatures [J]. Acta Bot Boreal-Occident Sin, 37(12): 2427-2434. [吴雪霞, 朱宗文, 张爱冬, 等, 2017. 外源褪黑素对低温胁迫下茄子幼苗生长及其光合作用和抗氧化系统的影响 [J]. 西北植物学报, 37(12): 2427-2434.]
YANG F, MIAO LF, 2010. Adaptive responses to progressive drought stress in two poplar species originating from different altitudes [J]. Silv Fenn, 44(1): 23-37.
YANG DH, CHEN F, SONG XQ, et al., 2012. Floristic composition and community characteristics of Dalbergia odorifera T. Chen, an endemic species to Hainan island [J]. Chin J Trop Agric, 32(12): 110-115. [杨冬华, 陈福, 宋希强, 等, 2012. 海南岛降香黄檀区系组成与群落特征分析 [J]. 热带农业科学, 32(12): 110-115.]
YE SX, 2008. Study on preliminary effect of Dalbergia odorifera T. Chen afforestation along different altitude of Minnan Mountain [J]. Strait Sci, 12(1): 95-96. [叶水西, 2008. 闽南丘陵山地降香黄檀不同海拔造林初步效果研究 [J]. 海峡科学, 12(1): 95-96.]
ZHANG N, SUN QQ, ZHANG HJ, et al., 2015. Roles of melatonin in abiotic stress resistance in plants [J]. J Exp Bot, 66(3): 647-656.
ZHAO HB, SU T, HUO LQ, et al., 2015. Unveiling the mechanism of melatonin impacts on maize seedling growth: sugar metabolism as a case [J]. J Pineal Res, 59(2): 255-266.
(责任编辑蒋巧媛)
猜你喜欢
江苏农业科学(2017年1期)2017-02-27
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年7期)2017-01-09
山东农业科学(2016年11期)2016-12-17
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
安徽农学通报(2014年12期)2014-07-28
