
神农架川金丝猴雄性社会结构研究
2022-05-16 10:26杨万吉余辉亮黄天鹏王晓菊杨敬元
野生动物学报 2022年2期
杨万吉 姚 辉 余辉亮 李 强 刘 强 黄天鹏 王晓菊 吴 锋 杨敬元*
(1.神农架金丝猴保育生物学湖北省重点实验室,神农架林区,442421;2.中南林业科技大学生命科学与技术学院,长沙,410004)
金丝猴是仰鼻猴属(Rhinopithecus)动物目前常用的中文名称,该属是旧大陆猴的一个分支,隶属于哺乳纲(Mammalia)灵长目(Primates)猴科(Cercopithecidae)疣猴亚科(Colobinaes)[1]。仰鼻猴属现有5个种,分别是川金丝猴(Rhinopithecusroxellana)、滇金丝猴(Rh.bieti)、黔金丝猴(Rh.brelichi)、怒江金丝猴(Rh.strykeri)和越南金丝猴(Rh.avunculus)。川金丝猴数量较多,分布范围较广,在四川的南坪、松潘、黑水等22个县,陕西秦岭南坡,甘肃岷山与邛崃山和湖北神农架均有分布;滇金丝猴分布于云南西北部和西藏的芒康地区;黔金丝猴主要分布于贵州的梵净山;越南金丝猴分布在越南北部;怒江金丝猴分布在云南省怒江州的高黎贡山和临近国界的缅甸地区[2-4]。前3种以川、黔、滇三省简称命名的金丝猴均是我国特有物种,被列为国家一级重点保护野生动物,同大熊猫(Ailuropodamelanoleuca)一样被誉为“国宝”,也被叫作“第二国宝”[5-7]。神农架川金丝猴仅分布于湖北省神农架地区,处于川金丝猴分布的最东端,也是湖北省唯一的分布区,其种群数量仅1 400多只,为一个典型孤立小种群[8]。通过研究了解其行为习性及社会结构变动,对于人类认识自我、了解自我具有极其重要的科学价值。
神农架川金丝猴成年雌雄具有明显的两性区别,成年雄性体型明显大于雌性,雌性体重约为雄性体重的52%,成年雄性嘴角上方具1对嘴角瘤,雌性无此外显嘴角瘤,成年雌性较雄性毛短且色淡。其次,川金丝猴不同生长阶段毛色变化明显,初生婴猴四肢细长瘦小,颜面部淡蓝色,双眼微睁。出生2个月后毛色开始变化,以后随着时间的推移毛色逐渐变化,至成年时毛色最为鲜艳[4,9]。
群体生活的灵长类动物的社会结构主要有一夫一妻制、一夫多妻制和多夫多妻制3种形式。一夫一妻制的灵长类群体中包括1个成年雄性、1个成年雌性和它们的后代,如白掌长臂猿(Hylobateslar)、西白眉长臂猿(Hoolockhoolock)和西爪哇叶猴(Presbytiscomata)等。一夫多妻制的灵长类单元内有1个成年雄性、多个成年雌性和多个青幼年个体,并由多个此类单元构成一个群体,如狮尾狒(Theropithecusgelada)、印度乌叶猴(Trachypithecusjohnii)和黑白疣猴(Colobuspolykomos)等。亚洲疣猴大多是一夫多妻后宫式的方式组合起来,这种后宫式的单元只有1个成年雄性留在单元内,其余成年和亚成年的雄性被该主雄逐出单元,另外组成全雄集体[10-11]。多夫多妻制的混杂结构中个体间有较明确的等级关系,最高等级的个体往往作为领头主导整个群体的行动,高等级的个体比低等级的个体具有更多的交配机会和食物资源[12],其中存在父系、母系两类模式,例如个别的长尾叶猴(Semnopithecusentellus)、长鼻猴(Nasalislarvatus)是以多夫多妻混杂的父系模式集合成群,橄榄绿疣猴(Procolobusverus)、东黑白疣猴(Colobusguereza)以多夫多妻混杂的母系模式集合成群[10-11]。
金丝猴的社会结构属于疣猴中典型一夫多妻后宫式的婚配方式。任仁眉教授等[13]对神农架川金丝猴社会结构的研究表明川金丝猴具有2个层次的基本社会单元,分别是1个成年雄性和多个成年雌性及多个青幼年个体组成的一雄多雌单元(one-male unit,简称OMU),多个成年或青年雄性组成的全雄单元(all-male unit,简称AMU)。Liu等[14]在对云南塔城的滇金丝猴研究中,也得出其具有同川金丝猴一样的一夫多妻后宫式的社会结构。另外,向左甫教授团队在对贵州焚净山的黔金丝猴[15]和云南高黎贡山的怒江金丝猴[16]社会结构研究中,同样指明它们社会结构的基本模式为一夫多妻制。严格来说,金丝猴群内的OMU中主雄具有绝对优势的交配权,青年雄性在性成熟前离开繁殖群OMU进入AMU中,而AMU中个体挑战主雄是替换OMU中主雄的最直接方式,且占更大的比例[17-18]。本研究在个体识别的基础上,详细记录神农架川金丝猴一个群体内雄性10年间在OMU中、AMU中和迁出群体的变动情况,对雄性迁移方式和迁移月龄进行分析,探索了解神农架川金丝猴雄性的社会结构变化,为神农架国家公园开展该物种的保护提供技术支持。
1 研究方法
1.1 研究对象
以神农架大龙潭区域分布的1个川金丝猴群体为研究对象。2005年始,神农架国家公园的工作人员选择大龙潭区域1个数量为38只的金丝猴群体进行长期跟踪调查,并适度人工补食,2007年成功实现了与该群体的近距离接触,截至2016年该群体数量增长至78只,OMU单元由3个增加至6个,AMU单元在1~2个间不断变动。在近距离观察基础上,对群体内每个个体根据外形特征和单元属性进行识别和命名,详细记录10余年来各个单元间雄性个体的迁移变动情况。
1.2 性别、年龄划分与个体识别
青年猴外形相似,性别只能从外生殖器辨别;成年猴除外生殖器外还有3个第二性特征可区分,雌猴在胸部可见明显的乳房和乳头,雄猴在嘴角可见豆粒状大小的肉瘤,雄猴头顶多具棕红色毛发,背上金黄色毛发明显长于和多于雌猴。
2006年开始观察时,已经在群体内的个体无法得知具体的出生日期,依据体型大小、毛色等,估计这些个体的年龄;2006年以后,所有个体出生均有准确的出生日期。将川金丝猴划分为5个年龄组。(1)婴猴:年龄在1岁以下,刚出生的婴猴外被毛为黑色,至3月大时黑毛脱光换上乳白色毛发,近1岁时头顶与背上长出少许黄色毛发。生长发育过程相对均匀,平均每年体重增加2~3 kg,体长增加7~9 cm。(2)青年猴:雌性1~3岁,雄性1~4岁,近2岁时头顶毛色逐渐变淡,随后发育中背部黄色毛发转浓变棕黑。(3)亚成年猴:雌性4岁,雄性5岁,身体大小接近成年个体,背部逐渐长出黑色背毛和金黄色披肩毛,头顶额两侧毛发逐渐变棕黄。(4)成年猴:雌性5~17岁,雄性6~17岁,成年个体最重可达20 kg,最大体长60 cm,背部金黄色披肩毛较多(成年雌猴在育幼阶段多会掉毛,披肩毛减少)。雌性个体乳房和乳头变得明显,雄性嘴角长出肉瘤。(5)老年猴:17岁以上的个体。
单元内许多川金丝猴具有一些典型的特征,一些有断尾、断指(趾)的情况,一些眼睛内有白色斑点,一些毛色较深,结合这些特征和五官轮廓、不同年龄个体大小差别、毛色差、不同性别等多次观察识别,对猴群中全部个体进行了命名及识别。
1.3 个体变动数据收集
2006—2016年在个体识别基础上,针对川金丝猴个体在群体内单元间变动和出走及进入群体的情况,完成了1 800多d(每年183~200 d)的观察记录。将个体的变动分为:迁出,个体离开研究群或消失;迁入,个体从研究群外进入群体;迁移,个体在研究群内加入或离开OMU或AMU,或组成新的OMU。
把雄性和雌性聚集在一起活动,稳定超过30 d定义为OMU的成立,OMU主雄位置的获得与失去各分2类。OMU主雄获得类型:一是孤雄与原OMU的主雄打斗,OMU中部分或全部雄性跟随孤雄组成新的OMU;二是孤雄与原OMU的主雄未有打斗,雌性主动从原OMU出走跟随孤雄组成新的OMU,雌性出走原因有刚成年避免近亲父本交配的出走,主雄与其他雄猴发生争斗时选择势强者或躲避性出走等。OMU中雌猴数量的增加和减少不列入主雄更替计数。
1.4 数据处理与分析
利用SPSS 19.0统计软件(SPSS Inc.,Chicago,USA)进行Spearman相关性分析和独立样本t检验,均使用双尾显著性检验,显著性水平设为0.05。部分数据仅进行简单的总和、均分统计及Excel表格作图。
2 结果与分析
2.1 OMU主雄的变更
2006—2016年,猴群中OMU单元发生了16次主雄的替换(表1)。在计算主雄替换次数时,另外一些不稳定的OMU单元(形成和消失在30 d以内的)并未统计。

表1 2006—2016年神农架川金丝猴单元主雄替换情况
所有16次主雄替换中,13次发生了新旧主雄的打斗,占全部的81.3%,其中11次替换集中发生在7—10月,为全部的68.8%,1、2、4月均未出现过变更(图1)。
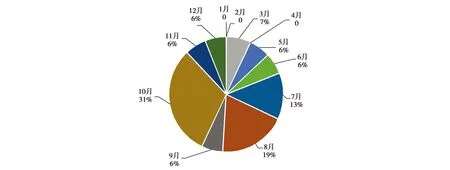
图1 2006—2016年神农架川金丝猴补食群单元主雄替换的月份分布Fig.1 The distribution of dominant male replacements golden snub-nosed monkeys in Dalongtan,Shennongjia from 2006 to 2016
2.2 OMU主雄的任期
统计了研究期间OMU单元主雄的任期(表2),其中研究开始时便在研究群中的主雄都记为2006年1月开始,而在2016年还未结束任期的,其结束时间记做2016年12月。

表2 2006—2016年神农架川金丝猴OMU单元主雄任期和配偶数
雄性任期平均为35.6个月(n=14,SD=27.5),任期最久的雄性为BTm,长达8.5 a;最短任期的是YZMm,仅持续8个月。通过Spearman相关系数分析发现,主雄任期的长短与其同单元最多成年配偶数呈正相关(r=0.644,P<0.05),而与雄性成为OMU主雄时的年龄呈负相关(r=-0.578,P=0.031)。排除DYm相隔3年第2次成为主雄的情况,可得雄性第1次成为OMU主雄的时间为76.2月龄(n=5,SD=3.4)。
2.3 青年雄性个体的迁移和迁出
对于雄性个体在单元间的迁移,主要关注它们从出生群首次迁移时的年龄,以及所迁至的单元类型。在观察到的19次首次迁移中,16次都是去往AMU,占84.2%。另外,雄性第1次迁移发生在36.5月龄时(n=19,SD=12.0),最晚首次迁移发生在52月龄时(表3)。至于迁出,排除掉1岁多随母离群的XWB和2次人为转移,余下16只雄性有9只约在60.6个月大时(n=9,SD=14.6)离开研究群,另外7只留在研究群。将迁出月龄与成为家长的月龄进行独立样本t检验,发现迁出的月龄显著小于成为家长的月龄(t=-2.327,P<0.05)。

表3 神农架川金丝猴青年雄性个体的迁移和迁出情况
3 结论与讨论
本研究发现,OMU主雄地位大部分是通过主动打斗获得的,而且主雄的更替多发生在8月和10月。川金丝猴为季节性繁殖物种,针对该猴群,之前的研究表明,其受孕时间集中在9—10月[19]。雄性替换发生在雌性受孕之前,可以增加新主雄的繁殖成功机会[20-21]。但是,秦岭川金丝猴的研究显示[22],大部分OMU的主雄替换模式中,没有观察到新旧主雄的打斗,或雄性个体身上的外伤,研究者推测是单元内雌性主导了主雄替换,侵入雄性采取不断靠近的策略吸引雌性迁移到自己的身边形成新单元[23]。而本研究中神农架川金丝猴通过打斗的直接替换占OMU主雄替换的80%,其他方式的主雄替换占20%,打斗替换过程不是短暂的一决胜负,从第1次打斗发生,至最后一次打斗,及至OMU原主雄迁出,最短的持续了10余天,最长的持续了6个月。AMU内个体替换OMU主雄的方式多种多样,其中通过引诱雌性的方式建立家庭的可能性不大,最终往往只有短暂的接触或聚集[17],期间OMU主雄对入侵雄性的驱赶也起了较大作用,比如阿拉伯狒狒(Papiohamadryas)和冠叶猴(Presbytispileata)会花费大量精力阻止全雄个体接近繁殖群内的雌性个体[24-25]。与之相比,直接挑战繁殖群主雄更易成功,所占替换方式的比例更高,同时本研究对于少于30 d的雌雄相聚不计入OMU构成事件也是两地金丝猴主雄替换方式产生差异的一个重要原因。但是,在主雄替换的打斗中一方或双方雄性常受重伤,甚至死亡。本研究16次主雄替换中发生13次打斗,冲突中均有雄性受伤,最重的一次为大胆嘴巴左边长6 cm的撕裂伤和右腿内侧贯穿咬伤。有研究报道了长鼻猴主雄替换事件中繁殖群主雄与全雄群冲突后受伤死亡事件[26],这也是一种优胜劣汰的自然选择。
青年雄性均在4岁之前从出生单元迁移出,且大部分是被主雄驱赶而迁移到AMU,迁移年龄明显小于性成熟的年龄。神农架地区及秦岭地区的川金丝猴群体研究均表明,雌猴第1次分娩年龄在5~6岁[27-28],减去川金丝猴妊娠期194~203 d[29-30],雌猴第1次与雄性交配并受孕时间为4~5岁,而亚成年雄性个体在3~4岁时爬跨行为明显增加[31],OMU主雄趋向于选择在亚成年雄性4岁前驱赶其离开,应该是避免近亲繁殖的一种方式。其次,雄性个体总是从出生OMU先迁移至AMU,然后再迁出离开研究群,或者通过打斗成为OMU的主雄。通过比较迁出研究群的月龄,和第1次成为家长的月龄,发现前者显著小于后者。雄性在成为主雄前迁出原群体进入新群体,很大程度减少与亲缘个体的接触,能有更多繁殖机会[32-33]。
个体BTm的任期超过8年,这无疑是不同寻常的,秦岭川金丝猴8年的观测研究也仅只发现1例BZT任期超过5年[33]。研究期间发现,BTm所在OMU中的雌性,常常会同BTm一起攻击AMU中的雄性或其他OMU的主雄,因此对于该主雄的任期,雌性在其中会起到较大作用,主雄的配偶数与主雄的任期长短呈现显著的正相关关系。主雄的年龄与任期也存在显著的负相关,研究也显示出更为年轻力壮的主雄能够打败年老体弱雄性挑战者,而雌性具有选择性帮助雄性的情况。另一方面,观察群雌性的首次受孕年龄大约为5岁,大于雄性的平均3年的任期(35.64个月),主雄在其雌崽性成熟前就被替换,或其雌崽迁出出生单元,这个情况在一些叶猴类[34-35]研究中也有报道,符合近亲交配避免假说[36]。综上所述,本研究中川金丝猴雄性个体迁移出出生单元、迁出出生群体、第1次成为主雄的方式和时间特点等社会结构变化的直接原因是不同雄性间的繁殖竞争,而其中一个重要影响因素是近亲繁殖避免机制。
猜你喜欢
云南画报(2021年10期)2021-11-24
学苑创造·A版(2021年9期)2021-09-16
东方少年·布老虎画刊(2021年1期)2021-06-22
小学阅读指南·低年级版(2020年9期)2020-10-12
人民周刊(2020年15期)2020-08-31
文萃报·周五版(2019年18期)2019-09-10
小天使·六年级语数英综合(2019年2期)2019-01-12
儿童故事画报·自然探秘(2017年7期)2018-03-14
幼儿教育·父母孩子版(2017年3期)2017-06-15
小猕猴智力画刊(2013年1期)2013-03-15
