
温度升高对长江口芦苇湿地细根形态和生长的影响
2022-05-26 10:02仲启铖张桂莲王开运郑思俊王小玲何晶李德志
生态学报 2022年9期
丁 一,仲启铖,张桂莲,王开运,郑思俊,王小玲,何晶,李德志
1 上海市园林科学规划研究院,城市困难立地生态园林国家林业和草原局重点实验室,上海城市困难立地绿化工程技术研究中心,上海 200232 2 华东师范大学生态与环境科学学院,上海市城市化生态过程与生态恢复重点实验室,上海 200241
据IPCC预测,到21世纪末,全球地表平均温度将较工业革命前上升1.5℃以上(IPCC 2014)。空气、土壤和水体的温度升高将会对人类赖以生存的多种生态系统的结构和功能产生深远影响[1]。滨海湿地位于陆地和海洋生态系统交界处,植被生产力高、有机碳分解速率低、土壤碳积累速率快[2],具有保育水生生物多样性、向近岸水域输送营养物质、过滤降解污染物、保护海岸线[3]以及作为蓝色碳汇[4—6]吸收和固持大气中的CO2等重要生态系统服务。目前已有一些研究采用室内培养、模拟增温和气候梯度等方法研究了升温对滨海湿地植物的影响及其反馈[3,7—9]。这些研究发现升温可能会增强外来入侵植物的扩散[9],改变植物群落结构[8],影响植物的生理和生长指标等。如Charles和Dukes[3]发现升温提高了北美盐沼优势植物互花米草(Spartinaalterniflora)的株高、基径和生物量。Kirwan等[10]预测2—4℃的升温将使互花米草群落的地上生产力提高10%—40%。李沙沙等[11]发现升温显著提高了崇明东滩芦苇(Phragmitesaustralis)和互花米草的单株叶面积、顶叶面积和叶面积指数。但是,与地上部分相比,目前关注全球变暖对滨海湿地植物地下部分影响的研究相对较少,而对地下部分最为活跃、最为敏感的湿地植物细根会对升温条件产生何种响应则更不清楚。
根系是植物最重要的功能器官,超过一半光合作用生产的有机碳会被植物分配到地下部分用于根系生长[12—13],并通过根系周转产生的残体和根系沉积输入根际土壤[14]。尽管直径≤2 mm的细根在总根生物量中仅占不到20%,但由于其处于不断的周转过程中,每年约1/3的全球净初级生产力被消耗用于生产和维持细根[15—16]。细根还是土壤中有机碳的重要来源,植物分配到地下部分的有机碳有15%—25%会通过细根周转输送到土壤碳库中[17—18],是陆地生态系统碳循环的重要环节之一。作为植物根系中活性最强、周转最快的组分,直径≤2 mm的细根对全球气候变化的反应更为敏感,并显著影响生态系统地下部分的碳平衡和养分循环[19],因此成为全球气候变化研究的热点之一。研究表明,升温对细根形态和生长的影响可能与物种[20]、微地形[21]、营养物质有效性[22]、土壤含水率[23]等有关,不同植物细根生产和死亡节律对升温响应的不一致性也会造成不同的细根功能性状变化[23]。Wang等[24]综述了177篇相关文献发现,升温总体上增加了细根生物量和生产量,但对其长度、形态和周转影响不显著,但这一研究缺乏来自滨海湿地的数据。微根管技术是目前研究细根生长动态的主流方法之一。与传统的土钻法[25]、内生长法[26—27]和挖掘法等方法相比,该方法通过在土壤中安装透明微根管,使用小型摄像机或扫描仪对细根进行长期、定点和非破坏性的连续监测[28],能直接捕捉不同土壤层次中细根的生长、死亡和周转等动态变化[29]。但目前这一方法在湿地生态系统中的应用还比较少见[30]。
崇明岛位于长江入海口,其沿河口海岸线分布的大片芦苇湿地为超大城市上海提供了多种重要的生态系统服务。项目组前期在崇明东滩的模拟升温实验发现,气温升高对芦苇湿地的土壤碳排放无显著影响,但却显著增加了其植被地上生物量[31—32];升温显著增加了表层土壤总有机碳含量,但对微生物生物量和相关酶活影响较小[33]。鉴于芦苇细根的活性和周转速率远高于根状茎,且其主要分布在0—40 cm土层[34],为了解升温条件下细根的潜在变化及其对土壤碳平衡的可能影响,本研究利用开顶式生长箱法对芦苇湿地的原状植物—土壤连续体进行模拟升温。2019年5—10月,采用微根管法和根钻法,对0—40 cm土层细根(本研究中指直径≤2 mm芦苇须根)的数量、长度、直径、表面积和生物量等性状特征开展了为期一个生长季的连续观测,并计算细根的净生长速率和周转速率,从而探讨气温升高对芦苇湿地细根形态、生长特征及土壤碳平衡的潜在影响,以期了解细根介导的地下碳循环对全球变暖的响应机理,并为开展滨海芦苇湿地碳汇功能的保护和修复提供科学依据。
1 研究区域与研究方法
1.1 研究区域概况
研究区域位于上海市崇明岛东滩 (31°38′N,121°58′E),该地区分布有长江口规模最大、发育最完善的滨海盐沼湿地之一。该区域属北亚热带海洋性季风气候,年平均气温15.3℃,年平均降水量为1004 mm。土壤类型为滨海盐碱土,以河口沉积物为母质。土壤含盐量在0.1‰—5.0‰之间,pH在8.2—9.2之间,有机质和养分含量较低[33]。崇明东滩也是长江口围垦规模最大的地区之一。样地所处地块在1998年被人工建设的堤坝圈围之前,是发育良好滨海盐沼湿地的高潮滩,以芦苇为原生优势种。该地区被堤坝圈围后,不再受潮汐周期性影响,但由于地势较为低平,在丰水期(如7—9月份)及降水丰富时地表仍不时出现淹水状态,年均水位约为地表以下30 cm。常见优势植物包括芦苇(P.australis)、白茅(I.cylindrica)、拂子茅(Calamagrostisepigeios)和荻(Triarrhenasacchariflora)等。
1.2 研究方法
1.2.1样方建设
开顶式生长箱(open top chamber,OTC)最早建设于2008年。选取地势均匀、植被均一的地块设置样地,OTC由8 mm厚的透明氟化玻璃和不锈钢框架制成,箱体高3.5 m,底部和顶部呈八边形,底部面积为12.5 m2,顶部开口略收紧[31],除必要结构性部件外,箱体均采用透光材料,并且底部留有通风口。将4个建设有OTC的样方作为升温样方(ET),同时紧邻每个OTC,等距离、随机性设置等面积、无OTC的对照样方(CK),具体样方设置如图1所示。2019年的温度监测结果表明,4—11月份空气升温幅度为1.41℃,土壤升温幅度为0.75℃,其中,7、8月份气温升幅最高。
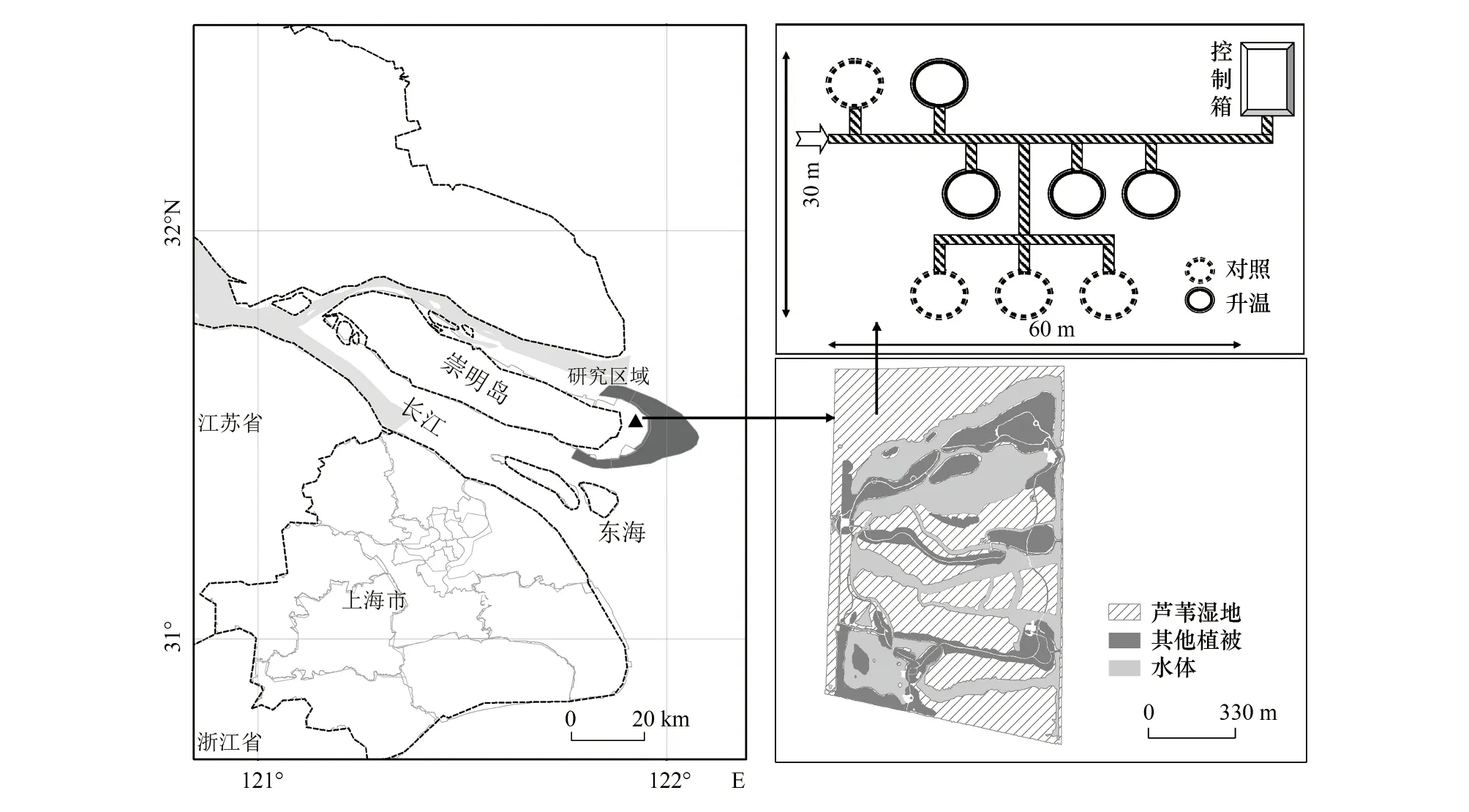
图1 研究区域及试验样地设置Fig.1 The location of the study area and the patterns of plots configurationCK:对照样方Control check;ET:升温样方Elevated temperature plots
目前,该样地的湿地植被群落已经退化,芦苇不再是单一的优势种。为严格模拟气温升高对长江口原生芦苇湿地植被的影响,2018年12月,于崇明东滩鸟类自然保护区内选取以芦苇为单优势种,植株高度、密度均一的湿地斑块(约20 m×20 m大小),间隔2—3 m、相互无干扰地挖掘植物—土壤连续体(长×宽×深:0.6 m×0.6 m×0.6 m),并移栽至升温设施所在样地。在每个样方内布设成对角分布的2个小样方,共16个小样方。在每个小样方内挖设长宽皆略大于原状土块,深度为0.6 m的坑体,并用厚度为5 mm的不锈钢板将四周围隔。将挖掘的原状芦苇植物—土壤连续体随机置放在16个小样方中,然后用移栽现场取回的土壤将空隙填实。ET和CK中土壤的初始理化性质如表1所示。

表1 试验开始前ET和CK中移栽样方各层的土壤理化性质Table 1 Soil physical and chemical properties of the transplanted plots in ET and CK in different soil layer before the warming treatment
在每个样方中,选取其中一侧小样方,首先用特制钢钻,以与地面成45°的夹角,钻一个与微根管直径相近的管洞,然后将有机玻璃质的透明微根管斜插入土壤,共埋设8 根(其中ET和CK样方各4根);另一侧则用于使用土钻进行根系定期取样。微根管外径 7 cm,内径 6.4 cm,长100 cm,埋入土中65 cm,垂直深度超过40 cm。在安装微根管时将密封段插入土壤中,并将管壁紧贴土体。微根管露出地表部分用黑色塑料布裹住,顶端开口处用黑色盖子密封,以避免水分和灰尘进入管中。
1.2.2细根图像的采集与处理
在完成植物移栽和微根管埋设后,用5个月的平衡期使根系生长和土壤恢复稳定。2019年5—10月,使用CI—600根系生长监测系统(CID BioScience Inc.)采集微根窗内的根系图像,每根微根管采集3张图像,每张图像面积为20.8 cm×21.7 cm。将图像按照微根管45°的倾角,划分到0—10 cm,10—20 cm以及20—40 cm土层。每月采集一次图像,整个生长季内共获取6次。采用Winrhizotron MF根系分析软件(Regent Instrument Inc.)进行图像数据的处理和分析,获取和计算微根窗内直径≤2 mm细根的总根长(Total root length TRL)(cm)、总表面积(Total surface area,TSA)(cm2)、平均细根直径(Average root diameter,ARD)(mm)、根长密度(Root length density,RLD)(cm/cm3)以及净生长速率(Net growth rate of root length density,RLDNGR)(cm cm-3d-1)。其中,RLDNGR通过计算一定观测时间段内RLD的变化来计算,公式如下:
RLDNGR=(RLDn+1-RLDn)/T
(1)
式中,RLDn+1与RLDn分别表示第n+1次与第n次观测到的细根根长密度,T为相邻两次观测的间隔天数。
RLD的计算公式如下[35]:
RLD=RL/A×DOF
(2)
A=C根管×D/sin45°
(3)
式中,RL为图片观测到的细根根长;A为微根窗观测区域的面积,C根管为根管图片的边长,即20.8 cm;D为划分的土层深度,即10 cm或20 cm;DOF为田间观测深度,即根管壁与土壤之间的距离,可计算获得,但会破坏样方;也可采用经验值,本中统一取0.2 cm[36]。
1.2.3细根样品的采集与处理
2019年6月期,在每个样方中无根管的一侧小样方选点,用内径5 cm土钻分别钻取 0—10 cm,10—20 cm和20—40 cm 土芯,带回试验室流水冲洗,手拣挑出全部根系并区分出直径≤2 mm细根,同时通过颜色区分死活根。通过LA-S根系扫描仪(万深检测科技有限公司)获取直径≤2 mm的活细根图像,并采用WinRHIZO Tron MF根系分析软件测定根长。将扫描好的根系装入牛皮纸信封内,65℃条件下烘干至恒重后称取生物量。采用如下公式计算比根长(SRL)(m/g)、比表面积(SRA)(cm2/g):
SRL=RL/B
(4)
SRA=RSA/B
(5)
式中,RL为扫描图片观测到的细根根长,RSA为扫描图片观测到的细根表面积,B为用根钻取出、洗净、烘干并称重的细根干物质量(g)。通过RLD和SRL计算单位体积细根生物量密度(RBDV)(g/m3),计算公式如下:
RBDV=RLD/SRL
(6)
单位面积的细根生物量RBDA(g/m2)通过单位体积细根生物量密度乘以取样深度D(m)来换算:
RBDA=RBDV×D
(7)
1.2.4细根周转速率计算
细根周转速率(T(a-1)采用极差法计算[37—38],公式如下:
T=BNPP/Bavg
(8)
BNPP=RBDAmax-RBDAmin
(9)
式中,BNPP为年净细根生产力(g m-2a-1),RBDAmax、RBDAmin分别为一个生长季中单位面积细根的最高生物量和最低生物量(g/m2),Bavg为月现存生物量平均值。
1.3 数据统计分析
采用SPSS 20.0对数据进行统计分析。在整个0—40 cm土层以及0—10、10—20、20—40 cm三个土层分别采用重复测量方差分析(RMANOVA),检验升温和观测时间对TRL、TSA、SRL、SRA、RBDA、ARD和RLDNGR等细根形态和生长指标的影响;对于升温与观测时间存在显著交互效应的指标,进一步采用单因素方差分析(One-way Anova),分别在每个观测时间点上,检验升温对细根形态和生长指标的影响。所有统计结果中,均以P<0.05作为存在显著性差异的标准。
2 结果与分析
2.1 升温在整个生长季对细根的影响
重复资料方差分析表明(表2),在整个生长季内,升温在0—40 cm土层显著增加了细根的总根长、总表面积、总生物量和净生长速率,增加幅度分别为125.8%、82.9%、100.5%和133.3%(表3),且观测时间与升温对这四个指标的影响均存在显著交互作用。升温对细根平均直径、比根长、比表面积无显著影响。
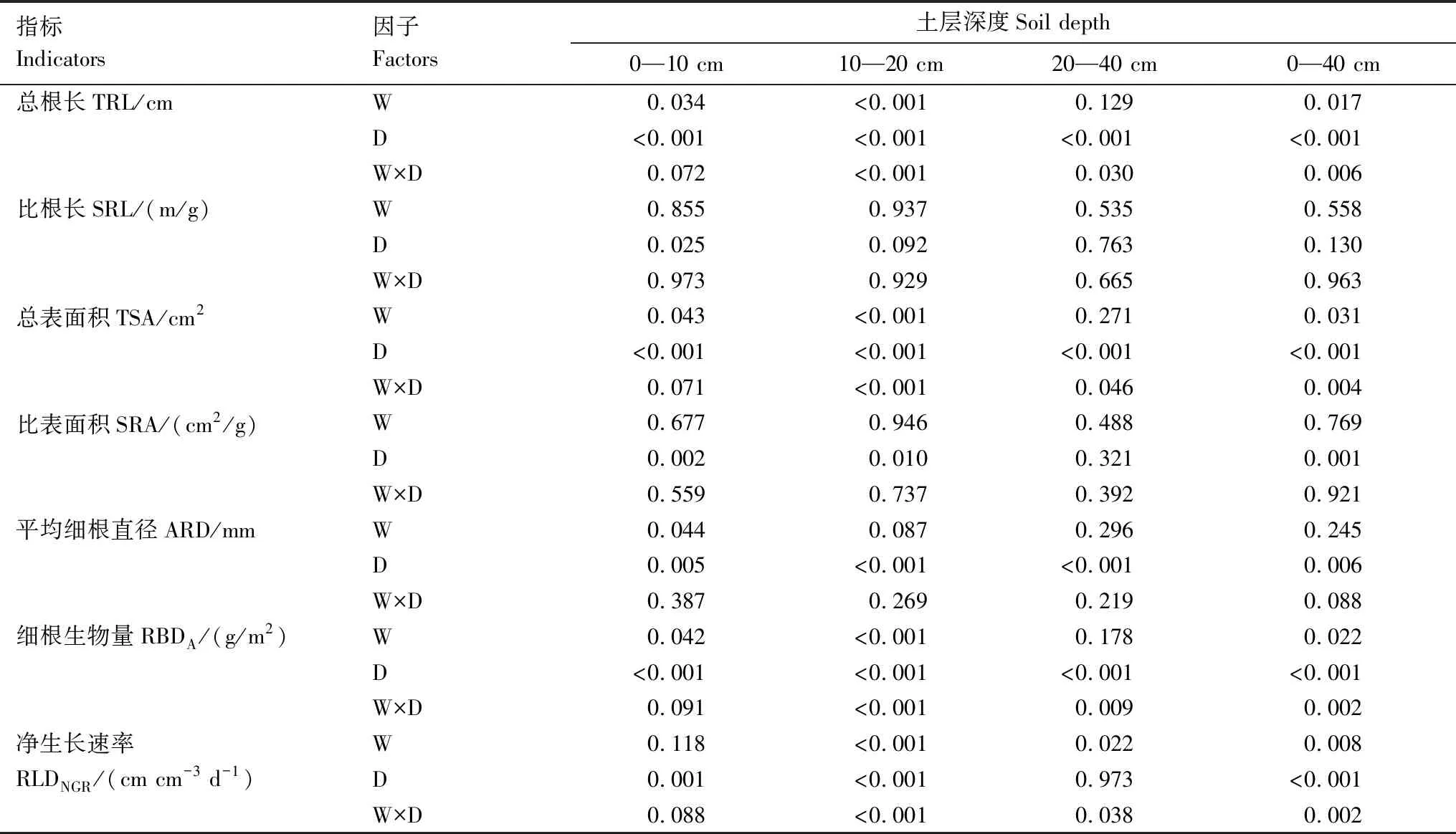
表2 整个生长季不同土层细根形态和生长指标的重复测量方差分析结果Table 2 Repetitive Measure ANOVA (RMANOVA)on the morphological and growth traits of fine roots of P.australis in different soil layer during the growing season

表3 整个生长季不同土层细根形态和生长指标的比较(平均值±标准误)Table 3 Comparisons of the morphological and growth traits of fine roots of P.australis in different soil layer during the entire growing season (mean±SE)
分层来看,升温处理显著增加了0—10 cm和10—20 cm土层细根的总根长、总表面积、细根生物量和净生长速率,且观测时间与升温处理的交互作用显著。升温还使0—10 cm土层细根的平均直径显著降低了13.2%。升温处理对0—10 cm和10—20 cm土层细根的比根长以及比表面积均未产生显著影响。与0—10 cm和10—20 cm土层相比,升温对20—40 cm土层细根的影响较小,仅显著提升了其净生长速率这一项指标。
2.2 升温在不同月份对细根的影响
如图2所示,在整个0—40 cm土层,CK与ET中细根总根长均呈现逐月上升的趋势,10月达到最大值,且前者月平均值始终高于后者。其中,5、8、9、10月,升温分别将细根总根长显著增加了67.1%、117.3%、149.5%和118.3%。在0—10 cm土层,除7月外,ET和CK中细根总根长在另5个月均差异显著,其中8月增幅最大,为166.2%;在10—20 cm土层,升温在除5月外的其他月份显著增加了细根总根长,其中9月增幅最大,达219.4%;在20—40 cm土层,升温仅在10月显著增加了细根总根长,增幅为90.1%。

图2 模拟升温对不同土层、不同观测时间细根总根长的影响Fig.2 Effects of experimental warming on total root length of fine roots of P.australis at different sampling date in different layer不同大写字母表示相同月份不同处理差异显著(P<0.05),不同小写字母表示相同处理不同月份间差异显著(P<0.05)
如图3所示,在整个0—40 cm土层,与总根长类似,CK与ET的细根总表面积同样呈现逐月上升趋势,且ET始终高于CK。其中8、9、10月二者差异显著,ET分别比CK分别高出93.1%、106.7%和96.4%。在0—10 cm土层,6、8、9月差异显著,其中8月增幅最大,为148.3%。在10—20 cm土层,除5月外,升温显著提高了细根总表面积,其中9月增幅最大,达153.9%。在20—40 cm土层,升温对细根总表面积无显著影响。
施药后7、15 d观察各处理的小麦生长情况,1%甲基二磺隆·双氟磺草胺可分散油悬浮剂各剂量对小麦安全性好,无明显药害症状;药剂 30 g/L 甲基二磺隆可分散油悬浮剂、50 g/L双氟磺草胺悬浮剂对小麦也无药害症状。收获时测产结果表明,各药剂处理小区产量要低于人工除草处理,与空白对照相比,均有不同程度的增产作用,增产率为7.84%~11.46%(表4)。

图3 模拟升温对不同土层、不同观测时间细根表面积的影响Fig.3 Effects of experimental warming on total root surface area of fine roots of P.australis at different sampling date in different layer
在整个0—40 cm土层(图4),升温显著提高了8、9、10月的细根生物量,分别达到100.7%、149.4%、122.5%。在0—10 cm、10—20 cm土层,除7月外,其他月ET的细根生物量均显著高于CK。其中,0—10 cm土层的细根生物量增幅8月最高,达143.3%;10—20 cm土层的细根生物量增幅9月最高,为274.4%;在20—40 cm土层,升温仅在10月显著增加了细根生物量。

图4 模拟升温对不同土层、不同观测时间细根生物量的影响Fig.4 Effects of experimental warming on root biomass density per unit area of fine roots of P.australis at different sampling date in different layer
如图5所示,在整个0—40 cm土层,CK与ET细根净生长速率在整个生长季中均呈“M”型曲线:即先升高后降低、再升高再降低。ET中细根净生长速率的季节波动幅度明显高于CK。在生长季初级T1(5—6月)、末期T5(9—10月),CK与ET细根净生长速率无显著差异;但在T2(6—7月)、T3(7—8月)、T4(8—9月),ET的细根净生长速率显著高于CK。其中,T4阶段升温对细根净生长速率的增幅最大(134.1%)。

图5 模拟升温对不同土层、不同观测时间细根净生长速率的影响Fig.5 Effects of experimental warming on net growth rate of root length density of fine roots of P.australis at different sampling date in different layer
同样的“M”曲线趋势出现在每个土层,但其波动幅度随土层加深趋于平缓。在0—10 cm土层,ET细根的净生长速率在T4显著高于CK,在T5显著低于CK;在10—20 cm土层,ET细根的净生长速率在T3和T4显著高于CK;在20—40 cm土层,ET细根的净生长速率仅在T4显著高于CK。
2.3 升温对细根垂直分布的影响
排除体积影响后发现,升温处理显著改变了芦苇细根在不同土层的分布(图6)。在CK中,除20—40 cm土层细根比表面积显著高于另外两个土层外,其他指标在不同土层之间均无显著差异。在ET中,20—40 cm土层细根比表面积与另两个土层无显著差异;0—10 cm、10—20 cm土层细根根长密度、表面积密度、生物量和净生长速率均显著高于20—40 cm土层。总体来看,升温使更高比例的细根分布在0—20 cm土层中。

图6 模拟升温对0—40cm 土层细根垂直分布的影响Fig.6 Effects of experimental warming on vertical distribution of fine roots in the 0—40 cm layer不同小写字母表示相同处理不同土层间差异显著(P<0.05)
2.4 升温对细根周转速率的影响
如图7所示,升温显著提高了10—20 cm以及20—40 cm土层的细根周转速率,增幅分别为32.9%和94.6%。然而,升温对0—10 cm土壤表层的细根周转速率影响并不显著。对比不同土层,在CK中,细根周转速率随土层加深呈下降趋势;但在ET中,不同土层之间的细根周转速率无显著差异。

图7 模拟升温对不同土层细根周转速率的影响Fig.7 Effects of experimental warming on turnover rate of fine roots of P.australis*表示相同土层不同处理间差异显著(P<0.05),不同小写字母表示相同处理不同土层间差异显著(P<0.05)
3 讨论
3.1 升温对芦苇湿地细根形态和生长的总体影响
本研究中,升温在整个生长季中显著提高了长江口芦苇湿地0—40 cm土层细根的生物量、总根长与总表面积,同时也增加了其净生长速率。这与Coldren等[20]通过原位升温试验发现的红树林湿地地下根系生物量升高的结果一致。Sullivan等[21]同样发现升温会导致北极沼泽植物的细根生物量增加。研究表明,如果土壤水分和养分有效性足够,根生物量和生长速率会随着温度升高而增加[39]。项目组前期研究发现,升温会使芦苇湿地土壤微生物氮、磷及相关酶活显著提升,增加微生物矿化作用[40—41],使植物可利用土壤养分增加,从而促进根系生长。同时,升温还会增大芦苇的光合作用[42]和总叶面积[11],光合产物总量增加会使分配给细根的量增加。另一方面,气温升高和叶面积指数同时增加[11]会显著增强芦苇的蒸腾作用,芦苇可能选择采用更长、更高表面积的细根来增加对土壤水分的吸收能力,从而维持其正常的生理代谢活动。
细根的比根长和比表面积反映了细根吸收土壤水分与养分的效率[43]。比值越大,相同的细根生物量会有更高的水分和养分吸收效率。平均细根直径同样可以表征植物细根吸收土壤养分、水分策略,也影响着细根的周转和寿命(周转的倒数)。本研究中,升温未对芦苇湿地细根的比根长、比表面积、平均细根直径产生显著影响。类似的,Björk等[44]在挪威北部进行的模拟升温试验也发现苔原湿地干草甸群落植物的比根长、比表面积未发生显著变化,但干石南灌丛群落细根的比根长与比表面积却显著升高。本研究中,0—40 cm的细根比根长、比表面积和直径未发生显著变化,其可能原因包括:(1)由于湿地水分和养分可用性高,芦苇无需明显改变其细根水分和养分吸收效率以应对升温对其光合作用和蒸腾作用的影响。(2)对根系较细的物种来说,调节比根长的能力相对较弱[45]。上述结果表明,在长江口芦苇湿地中,升温可能会通过促进细根的伸长来增加其总表面积和总生物量,而不会显著改变细根的形态特征,以及水分和养分吸收效率。
细根以周转方式进入到土壤的碳含量约占土壤碳总输入量的14%—50%[12],细根周转深刻地影响着土壤碳循环过程。Fitter等[46]在对英国一个高地草原的土壤升温试验中发现,升温会导致细根周转速率的增加,这与我们的研究结果一致。导致这一现象的原因可能是由于升温条件下细根的生理活性增加、寿命变短,同时微生物矿化作用增强使养分供应增加[47],进而促进细根新陈代谢增强、周转速率加快;另一个可能的原因是升温导致细根总量和密度增加,细根之间竞争加剧导致细根生产、死亡加速[48],周转加快。
3.2 升温对芦苇湿地细根形态和生长时空格局的影响
植物根系的季节动态反映了其物候,生境条件变化如土壤水分、养分的改变通过影响植物细根的水养吸收而改变其季节动态[49],而光合作用强度通过影响光合产物向地下的运输和分配影响细根的生长动态[50]。本研究中,芦苇湿地细根的生物量、总根长、总表面积持续呈逐月升高趋势,即生产大于死亡,现存量持续增加。升温处理提高了湿地细根现存量的增加幅度,使升温与对照的差值逐月扩大。细根生长的季节动态有单峰型曲线、双峰型曲线[51—52]。本研究中,芦苇细根在整个生长季中的净生长速率呈双峰型曲线,且升温导致其季节变异性进一步增大。升温在6—9月持续促进了整个0—40 cm土层的细根净生长速率,其中在8—9月最为显著。7—8月芦苇细根净生长速率相较于6—7月、8—9月相对降低,可能是由于上海2019年梅雨季持续到7月下旬,连续阴雨天气导致芦苇光合作用强度降低,影响了光合作用产物的生产和分配,从而影响植物细根的生长。
细根的垂直分布格局显示了芦苇的营养获取策略,与物种、气候因子以及土壤理化性质密切相关。一般认为,植物细根的分布随土层加深而减少[53—55]。本研究中,对照中不同土层细根根长和生物量的差异不显著。升温使0—20 cm土层细根总根长、总表面积和生物量显著增加,高于20—40 cm土层,这说明升温条件下,细根更加趋向于集中在表层0—20 cm分布[44]。这种垂直梯度上细根分布的变化可能与升温后不同土层中养分的有效性或养分向根的扩散速率的变化有关。Zhong等[33]发现升温使芦苇湿地0—10 cm土层总有机碳、总氮显著提升,而在10—40 cm土层无显著影响。另外,本研究中,升温在不同土层对细根比根长、比表面积、平均细根直径影响均不显著,说明不同土层根系水分和养分吸收效率均未受显著影响。
升温对细根净生长速率的影响在不同土层也存在一定差异。其中,与10—20 cm和20—40 cm土层相比,升温对0—10 cm土层细根净生长速率提升幅度最小,仅在根系生长最快的6—7月、8—9月呈现显著促进作用。这可能是因为该地区土壤为滨海盐碱土,升温条件下蒸发散作用增强会导致表层土壤含盐量显著提高[33],土壤含盐量增加则可能会抑制了细根的正常生长[56—57],限制了升温在某些月份对细根净生长速率的促进作用。
升温对不同土层的细根周转速率产生了不同的影响,升温处理显著提升了10—20 cm、20—40 cm土层的细根周转速率,但对0—10 cm土层无显著影响,导致升温后不同土层的细根周转速率之间差异不显著。与净生长速率一样,0—10 cm土层细根周转速率未产生显著变化,同样可能是由于表层土壤含盐量增加[56—57],从而对细根的代谢和周转产生了潜在抑制。
3.3 升温条件下芦苇细根变化对土壤碳平衡的潜在作用
项目组前期研究发现,升温导致芦苇湿地土壤表层有机碳增加,但对微生物生物量碳无显著影响,我们推测这种现象与升温引起的根源有机碳输入的增加有关。本研究中,结合对根系周转速率及其有机碳含量的测定结果,对根系周转的年度有机碳输入量进行估算。估算结果显示,升温条件下,芦苇湿地通过细根周转进入土壤的有机碳量在0—10 cm、10—20 cm、20—40 cm土层分别增加了55.08、89.27、39.67 g m-3a-1,初步证实了我们前期的推测。Björk等[44]在研究苔原植物对升温的响应时认为,由于升温导致细根生物量的提高和形态的改变,造成更多根源碳输入土壤。值得注意的是,在本研究中,尽管升温未导致0—10 cm土层细根周转速率发生显著变化,但由于参与周转的细根生物量显著增加,该层的根源有机碳输入也同样增加。
根据Wang等[58]的预测,在RCP4.5或RCP8.5气候变化情境下,尽管总面积可能会减少,但由于红树林湿地中温度和降水的增加,以及盐沼湿地中相对海平面的升高,滨海湿地碳的净积累将持续增加。然而,该研究采用的沉积物断代技术(sediment dating techniques)无法确定植物地下部分生产量增加导致的碳积累变化。本研究通过对模拟升温条件下芦苇湿地细根生长和形态指标的连续监测,则为了解气候变化条件下滨海湿地植物根系的变化,及其对滨海湿地碳循环的影响提供了新的依据。本研究结果表明,对于水文条件良好、生态系统健康的滨海芦苇湿地,气候变暖可能会在一定程度上促进表层土壤的根源有机碳输入,从而导致湿地土壤碳储量的增加。另外,除根系周转外,植物根系还能将分配到根部的有机、无机化合物释放到周围土壤形成根际沉积,其中根系分泌物是最重要组分[59]。后续研究将结合根系形态、生长和分泌物指标测定,进一步探讨升温条件下芦苇湿地细根对土壤碳平衡的影响机制。
4 结论
在长江口芦苇湿地中,通过一个生长季的连续监测和分析发现,模拟温度升高显著增加了0—40 cm细根(≤2 mm)的总量(总根长、总表面积、生物量),而对反映细根水分与养分吸收效率的指标(比根长、比表面积、直径)无显著影响。细根总量的增加主要源自0—20 cm土层细根指标的变化,20—40 cm土层的贡献相对较小。升温显著提高了细根净生长速率,增强了其“M”型曲线的生长节律,而这种促进作用随土层加深而逐渐减弱。升温显著增加了10—40 cm土层的细根周转速率,而在0—10 cm土层影响不显著。升温使细根周转速率加快以及参与周转的细根总量增加,导致各土层特别是0—20 cm土层根源有机碳输入显著增加,这可能会深刻影响芦苇湿地的土壤碳平衡。因此,未来在开展滨海湿地土壤碳平衡的模拟和预测时,应当充分考虑升温条件下植物细根相关碳过程的可能变化。
猜你喜欢
草业学报(2022年9期)2022-09-16
物流技术与应用(2022年8期)2022-08-26
东北林业大学学报(2022年6期)2022-07-25
科学家(2022年5期)2022-05-13
小小说月刊·下半月(2021年8期)2021-09-05
绿色科技(2021年5期)2021-04-09
四川农业大学学报(2018年3期)2018-09-04
动漫界·幼教365(大班)(2018年3期)2018-05-14
制造技术与机床(2017年7期)2018-01-19
湖南大学学报·自然科学版(2016年12期)2017-05-12
