
千金子叶绿体基因组的结构及特征分析
2022-05-30 10:48田祥瑞徐梦彬王飞叶凌凤
杂草学报 2022年2期
田祥瑞 徐梦彬 王飞 叶凌凤
摘要:千金子是稻田一年生禾本科恶性杂草,已成为威胁水稻生产的优势种群。采用Illumina HiSeq测序及生物信息分析方法对千金子叶绿体全基因组序列进行测定、组装、注释和解析。结果表明,千金子叶绿体基因组全长为135 656 bp,GC含量为38.26%,呈现典型的禾本科植物叶绿体基因组的4段闭合环状结构;其中,大单拷贝区80 843 bp,小单拷贝区12 768 bp,反向重复区42 045 bp;通过功能富集,共获得注释基因131个,包括84个蛋白编码基因、39个tRNA基因和8个rRNA基因;另外,还检测到49条简单重复序列(SSR)和46条长度大于20 bp的重复序列。利用IQTREE软件,基于叶绿体基因组以粳稻为外群构建了25种禾本科杂草的系统进化树,显示出千金子与真穗草属、虎尾草属和狗牙根属杂草具有相对较近的亲缘关系,而3种稗属杂草则与水稻的亲缘关系更近。本研究结果可为千金子的精准识别和分子进化解析提供理论借鉴。
关键词:千金子;叶绿体基因组;结构特征;功能基因注释;SSR;系统发育进化树
中图分类号:S451 文献标志码:A 文章编号:1003-935X(2022)02-0006-09
Structure and Characterization Analysis of Chloroplast Genome of Leptochloa chinensis
TIAN Xiang-rui,XU Meng-bin,WANG Fei,YE Ling-feng
(Institete of Mordern Agriculture,Jiangsu Provincial Agricultural Reclamation and Development Corporation,Nanjing 210031)
Abstract:Leptochloa chinensis (L.) Nees is an annual gramineousmalignant weed in rice field,which has become a dominant weed population threatening rice production.In this study,Illumina HiSeq sequencing and bioinformatics analysis methods were used to determine,assemble,annotate and analyze the whole chloroplast genome sequence of L.chinensis. The results indicated that the full length of L. chinensis chloroplast genome was 135 656 bp and the content of GC was 38.26%,showing a typical four segment closed ring structure of chloroplast genome of gramineous plants;among them,the large single copy region was 80 843 bp,the small single copy region was 12 768 bp,and the inverted repeat region was 42 045 bp;through functional enrichment,131 annotated genes were obtained,including 84 protein encoding genes,39 tRNA genes and 8 rRNA genes;in addition,49 simple sequence repeats (SSRs) and 46 repeats longer than 20 bp were detected. Using IQTREE software,the phylogenetic tree of 25 weeds was constructed based on chloroplast genomes and japonica rice as outgroup;it showed that there were relatively close relationships among L. chinensis,Eustachys,Chloris and Cynodon,while 3 Echinochloa weeds were more closely relatedtorice.ThisstudyprovidesatheoreticalreferencefortheaccurateidentificationandmolecularevolutionanalysisofL.
收稿日期:2022-03-28
基金項目:江苏省重点研发计划(编号:BE2019396);江苏省“双创博士”及南京市留学人员科技创新计划。
作者简介:田祥瑞(1989—),男,河南新乡人,博士,农艺师,主要从事作物有害生物综合治理技术研究。E-mail:635369028@qq.com。
chinensis.
Key words:Leptochloa chinensis;chloroplast genome;characterization;gene function annotation;SSR;phylogenetic tree
千金子[Leptochloa chinensis (L.) Nees]为稻田一年生禾本科恶性杂草,21世纪以来,伴随少免耕栽培模式及直播水稻的大面积推广,千金子在我国南方稻田迅速蔓延[1]。加之生产上长期使用氰氟草酯进行茎叶处理,缺乏合理的药剂轮换,国内已发现多个千金子田间种群对氰氟草酯产生了不同水平的抗药性[2]。生产上氰氟草酯的用药量逐年递增,防除难度越来越大,已演变成影响水稻生产的优势杂草之一,严重威胁粮食安全。当前,对千金子的研究集中于发生规律、化除技术、抗性机制等方面,在利用高通量测序技术获得的225个植物基因组中,有32个属于禾本科植物,其中包括4种禾本科杂草[3],已报道转录组测序的杂草有菵草[4]、稗草[5]、牛筋草[6]、杂草稻[7]等。由于千金子的全基因组和转录组序列尚未公布,其遗传背景和系统进化等方面的研究仍需加强。
叶绿体基因组(cpDNA)为双链环状分子,是母系单亲遗传细胞器DNA,其序列和结构保持相对稳定保守,极少发生基因重复,易于提取、净化、测序。叶绿体rbcL、matK、trnH-psbA基因与核基因ITS片段相结合,被推荐作为高等植物的DNA条形码[8]。叶绿体基因组简单重复序列(simple sequence repeat,SSR)特异性、重现性较高,成为分析遗传多样性和物种起源和演化的重要标记[9];叶绿体基因组单核苷酸多态性(single nucleotide polymorphism,SNP)突变较为丰富,测序结果可直接用于生物统计和遗传分型。基于多样化的分子标记技术,叶绿体基因组序列已被广泛应用于植物遗传进化、亲缘关系及分类鉴定研究,如大田作物[10-12]、水果花卉[13-15]、茶叶[16]、药用植物[17-18]、濒危树种[19-20]等。
禾本科植物的叶绿体基因序列长度在115~150 kb之间,水稻、小大麦、高粱等主要作物的叶绿体基因组序列陆续在NCBI公布[21]。在国内的杂草叶绿体研究中,利用稗草叶绿体基因组trna-b1及psbA标记可将稗属材料分为4个大类[22];杂草稻的叶绿体基因组序列具有显著多态性[23];在杂草稻叶绿体基因组中发现了1 020个SNP位点,新开发trnG-trnfM、psbM-trnC及trnT-trnL等3对叶绿体分子标记[24];另外,山羊草[25]、鼠尾草[26]的叶绿体基因组也被测序组装和注释分析。本研究采用Illumina HiSeq测序及生物信息分析方法,对千金子的叶绿体基因组序列及结构特征进行分析,以期为千金子的精准识别鉴定提供理论依据。
1 材料与方法
1.1 叶绿体基因组的提取与测序
千金子整株杂草于2020年7月采集于江苏省黄海农场水直播稻田(34°15′3″N,119°58′15″E),在运输过程中用湿润的脱脂棉裹住根系以保证杂草活力;到達实验室后将样品洗净晾干,剪取叶片作为高通量测序样品置于液氮中冷冻保存。采用MagMAX植物 DNA 试剂盒(Thermo Fisher)对千金子叶片进行基因组DNA的提取与纯化,样品送苏州金唯智生物科技有限公司进行高通量测序。
采用Illumina HiSeq测序仪进行基因组De Novo测序,采用VAHTS Universal DNA Library Prep Kit for Illumina 试剂盒进行文库构建。具体过程如下:200 ng gDNA用超声波破碎仪(Covaris S220)随机打断成小于500 bp的片段,然后通过End Prep Enzyme Mix进行末端修复,两端加测序接头。再使用VAHTSTM DNA Clean Beads纯化470 bp左右的片段,最后用P5和P7引物进行扩增。使用Qsep100 (Bioptic)仪器检测文库质量,并且通过Qubit 3.0检测文库浓度。DNA文库混合后,按 Illumina NovaSeq仪器使用说明书进行2×150 bp双端测序,由NovaSeq自带的NovaSeq Control Software (NCS) + OLB + GAPipeline-1.6软件读取序列信息。
1.2 叶绿体基因组的组装与注释
测序的Reads数据通过质量控制后,采用velvet软件进行组装和拼接,使用SSPACE及GapFiller软件进行缺口补洞和修正。采用GoSeq软件寻找编码基因,使用tRNAscan-SE程序和检测转移RNA(tRNAs)、RNAmmer程序鉴定核糖体RNA(rRNA)。利用NCBI中的BLAST程序将千金子叶绿体基因组序列与核酸数据库进行比对注释,采用GO数据库进行基因功能富集分析、KEGG数据库进行基因通路富集分析。将组装注释完成的千金子叶绿体全基因组序列通过BankIt路径上传至GenBank数据库。
1.3 SSR与重复序列的预测分析
利用在线软件MISA(http://webblast.ipk-gatersleben.de/misa/index.php)分析鉴定千金子叶绿体全基因组的简单重复序列,设定最小重复次数:单核苷酸≥10,二核苷酸≥6,三核苷酸≥5,四核苷酸≥3,五核苷酸≥3,六核苷酸≥3;使用在线程序REPuter(http://bibiserv.cebitec.uni-bielefeld.de/reputer)对千金子叶绿体基因组中的正向重复(forward repeats)、反向重复(reverse repeats)、互补重复(complement repeats)及回文重复(palindromic repeats)等4种重复序列进行分析,设置最小重复(minimal repeat size)值为20、海明距离(hamming distance)值为3。
1.4 禾本科杂草的系统进化分析
在NCBI数据库中选取25种禾本科杂草的叶绿体基因组序列进行系统进化分析。通过MAFFT软件将序列矩阵对齐;经过检验和校正后,将对齐后的序列导入IQTREE软件,以粳稻叶绿体基因组全序列作为外群,构建系统发育树,使用嵌入式超快引导方法(UFBoot)估计节点支持值。
2 结果与分析
2.1 千金子叶绿体基因组的结构特征
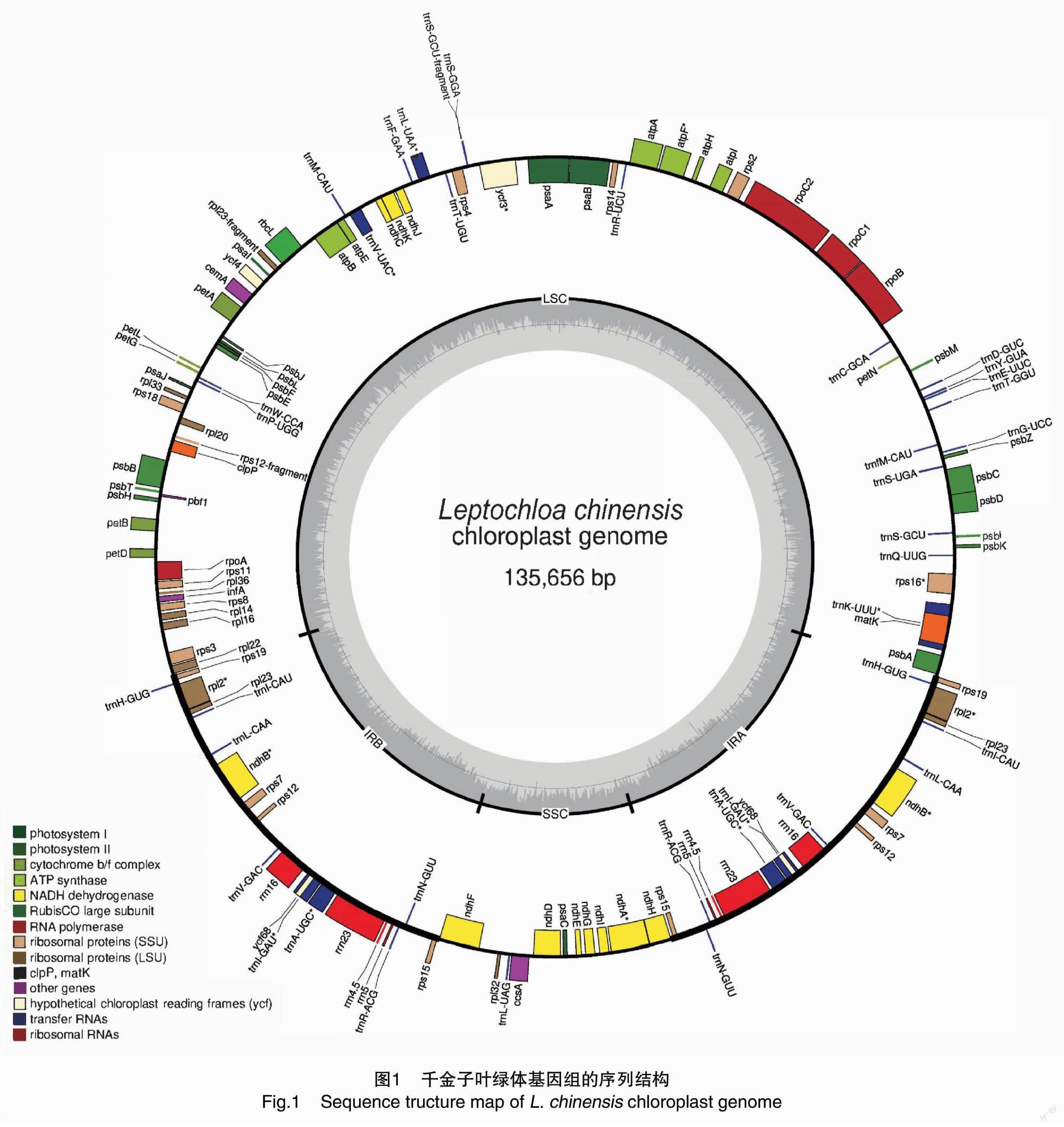
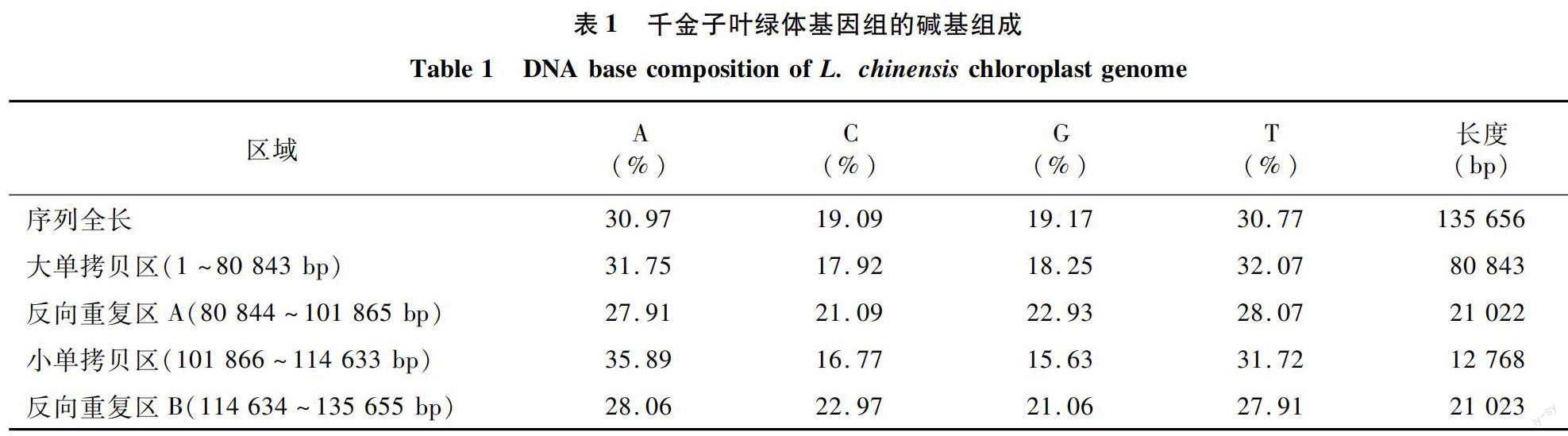
千金子叶绿体基因组序列全长为135 656 bp,GC含量为38.26%,将序列上传至GenBank,获得登录号MZ901215。该基因组呈现典型的4段闭合环状DNA结构(图1),其中,大单拷贝区(LSC)长度为80 843 bp,GC含量为36.17%;小单拷贝区(SSC)长度为12 768 bp,GC含量为32.40%;反向重复区A(IRA)长度为21 022 bp,GC含量为44.02%;反向重复区B(IRB)长度为21 023 bp,GC含量为44.03%(表1);2个反向重复区结构对称、方向相反,被大、小单拷贝区隔开。
2.2 千金子叶绿体基因组的功能基因注释
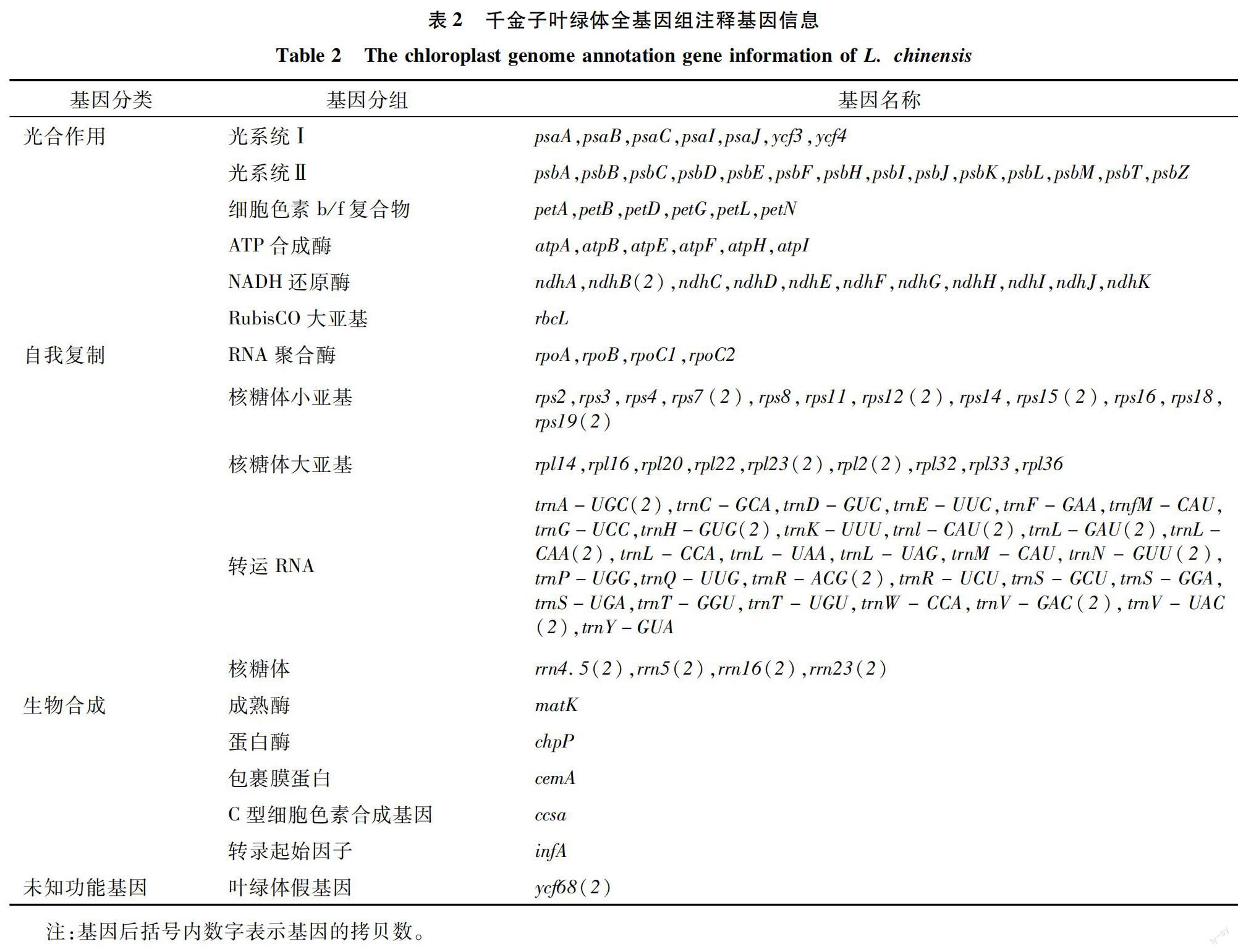
千金子叶绿体基因组共有131个基因,包括89个单拷贝基因和21个双拷贝基因。其中,蛋白编码基因(PCG)总计84个,编码区碱基总长度 55 851 bp,最短基因长度91 bp,最长基因长度 4 538 bp,编码基因的平均序列长度725 bp,GC比例为38.91%。非编码ncRNA总计47个,包括核糖体RNA(rRNA)8个,转运RNA(tRNA)39个;LSC区域有82个基因,包括58个PCG和24个tRNA;SSC区域有38个基因,包括37个PCG和1个tRNA;IRA及IRB区域的基因序列相同、对称分布,共有19个双拷贝基因,包括8个双拷贝PCG(rps19、rpl2、rpl23、ndhB、rps7、rps12、ycf68、rps15),7个双拷贝tRNA(trnL-CAU、trnL-CAA、trnV-GAC、trnL-GAU、trnA-UGC、trnR-ACG、trnN-GUU)和全部4个双拷贝rRNA(rrn16、rrn4.5、rrn23、rrn5)。
利用GO、KEGG数据库进行基因功能注释(表2),光合作用有关基因(光系统、细胞色素b/f复合物、ATP合成酶、NADH还原酶、RubisCO大亚基)富集到46个,自我复制有关基因(RNA聚合酶、核糖体亚基、rRNA、tRNA)富集到78个,生物合成有关基因(成熟酶、蛋白酶、包裹膜蛋白、C型细胞色素合成基因、转录起始因子)富集到5个(matK、chpP、cemA、ccsa、infA),未知功能基因是双拷贝的叶绿体假基因ycf68。
2.3 千金子叶绿体基因组的SSR及重复序列分析
通过设置不同长度基序的最小重复次数,由表3可知,在千金子叶绿体基因组中发现简单重复序列49个。其中,单核苷酸重复为37个,占比75.51%,基序A(n)、C(n)、G(n)、T(n)的SSR重复分别为17、2、1、17个;二核苷酸重复为2个,AT(6)和TA(6)重复1次;无三核苷酸重复;四核苷酸重复为8个,占比16.33%,基序AACG(3)、AATA(3)、AGAA(3)、ATAC(4)、GTAG(5)、TCGT(3)、TTCT(3)、TTTA(3)重复1次;五核苷酸重复为2个,AAAGT(3)、TTTCTA(3)重复1次。
通过设置序列最小长度,在千金子叶绿体基因组中发现46条长度大于20 bp的重复序列,其中有正向重复25个,回文重复21个,无正向重复和互补重复(表4);最长的重复序列大小为52 bp,分别位于trnN-GUU-ycf1、rps15-ycf1和rps18;最短重复序列大小为29 bp,位于trnS-UGA和trnS-GCU;有5条重复序列分别位于ycf3*、ndhB*的内含子中。
2.4 基于叶绿体基因组的禾本科杂草系统发育分析
基于25种禾本科杂草的叶绿体基因组全序列,以粳稻叶绿体基因组作为外群构建进化树(图2),其中22种禾本科杂草聚为一个大支,稗(E. crusgalli)、稗原变种(E. crusgalli var. crusgalli)以及细叶旱稗(E.crusgalli var. praticola)等3种稗属杂草单独聚为一小支。22种禾本科杂草的大支分為2个分支,第1分支由千金子属(L. chinensis、L. virgata、L. pluriflora)、真穗草属(E. glauca)、虎尾草属(C. virgata、C.barbata)、狗牙根属(C. dactylon)聚成的分支,三齿稃草属(T. basedowii、T. scintillans、T. lanigera、T. vanleeuwenii、T. concinna)、固沙草属(O. kokonoricus)、锋芒草属(T. australianus)聚成的分支,以及结缕草属(Z. japonic、Z. tenuifolia、Z. matrella)的分支组成;第2分支由香根草属(H. macra)和芦苇属(P. australis)的小支、鹧鸪草属(E. major、E. obtuse)的小支以及小丽草属(C. africana)聚成。从亲缘关系来看,千金子与真穗草属、虎尾草属和狗牙根属杂草具有相对较近的亲缘关系,而稗属杂草则与水稻的亲缘关系更近。
3 讨论与结论
禾本科植物的叶绿体基因组一般呈现闭合环状DNA结构,已公布的禾本科作物的叶绿体基因组大小在115~150 kb之间,GC含量均在37.1%~38.9%之间,大单拷贝区长度约80 kb,小单拷贝区长度约13 kb,每个反向重复区长度约20 kb,蛋白编码基因与tRNA的种类和数量存在物种间差异,而rRNA的种类和数量在物种间保持相对稳定[21]。水稻叶绿体基因组为134 525 bp(NC_008155),注释基因162个;稻田恶性杂草水田稗的叶绿体基因为139 891 bp,大单拷贝区长 82 108 bp,小单拷贝区长13 205 bp,反向重复区长44 578 bp,注释基因131个,包含双拷贝基因19个[22]。本研究测序组装的千金子叶绿体基因组序列长度为135 656 bp,大单拷贝区长 80 843 bp,小单拷贝区长12 768 bp,反向重复区长42 045 bp,共有基因131个,包括84个蛋白编码基因、39个tRNA基因和8个rRNA基因;其双链闭环结构与经典的叶绿体结构相吻合,大、小单拷贝区,反向重复区的序列长度符合禾本科植物相关特点,注释基因数量、双拷贝基因数与稗草相似,补充了我国稻田重要恶性杂草的关键基因信息。
对于叶绿体基因组的编码基因及功能方面,禾本科植物之间具有较小差异。与光合作用有关的ycf1、ycf2被发现在部分禾本科植物中逐渐退化[27],本研究同样没有在千金子中发现。与乙酰辅酶A羧化酶合成有关的accD基因在水稻中存在,而在小大麦、玉米、高粱及黑麦中发生退化缺失[28],本研究也发现accD的缺失现象;千金子叶绿体4个rRNA的种类与主要禾本科作物、稗草完全一致,也显示出rRNA在禾本科植物中的高度保守性。本研究检测的49个SSR基序和46条重复序列,提供了独特的千金子DNA识别信息,将为开发叶绿体分子标记提供依据。
叶绿体基因组多为直系同源,对揭示植物进化及亲缘关系具有重要作用。冯克伟利用禾本科叶绿体全基因组序列揭示了不同亚科、不同物种间的亲缘关系,证实竹亚科与早熟禾亚科相对稻亚科更近[29]。林张翔利用稗属叶绿体基因组序列的进化分析推算稗属与稷属的分化约在21.6百万年前,水田稗与稗在3.3百万年前发生分化[22]。朱国忠等对13份栽培稻和野生稻的叶绿体基因组数据进行系统发育分析,发现不同籼粳类型的栽培稻和野生稻分别被聚类到相应类群的杂草稻中[24]。本研究首次基于叶绿体基因组对25种禾本科杂草的系统进化及亲缘关系进行分析,发现稗属杂草与水稻亲缘关系较近,千金子属、真穗草属、虎尾草、狗牙根属亲缘关系较近,三齿稃草属、固沙草属、锋芒草属、结缕草属亲缘关系较近,香根草属、鹧鸪草属、小丽草属亲缘关系较近,该结果与传统的禾本科植物形态分类学相对应,成为研究禾本科杂草系统发育和演化的科学参考。
千金子作为重要的稻田恶性杂草,在水稻生长前期进行土壤封闭和苗后早期进行茎叶处理可以控制其危害[30]。因此,早期精准识别是提高药剂化学除草效果的重要前提,而目前生产上对千金子的早期鉴定仅依赖于形态学识别,存在一定的局限性,本研究解析的千金子叶绿体基因组结构与特征,将为千金子的精准鉴定和分子进化解析提供理论借鉴。
致谢:感谢江苏省农垦农业发展股份有限公司黄海分公司孙文忠书记、苏垦农发黄海分公司董梦雅主任对千金子田间样品采集的帮助。
参考文献:
[1]杨浩娜,王立峰,邬腊梅,等. 稻田恶性杂草千金子的抗药性研究进展[J]. 农药学学报,2019,21(增刊1):772-779.
[2]武向文,王法国,曹 青. 华东部分稻区水稻田千金子对氰氟草酯的抗性[J]. 农药学学报,2019,21(3):285-290.
[3]Gaines T. Part Ⅱ Grass genomics[M]//Kraehmer H. Grasses:crops,competitors,and ornamentals. New Jersey:John Wiley & Sons,Ltd.,2019:14-15.
[4]潘 浪. 麦田菵草(Beckmannia syzigachne)对精唑禾草灵抗药性及其机理研究[D]. 南京:南京农业大学,2018.
[5]贾新平,叶晓青,梁丽建,等. 基于高通量测序的海滨雀稗转录组学研究[J]. 草业学报,2014,23(6):242-252.
[6]An J,Shen X F,Ma Q B,et al. Transcriptome profiling to discover putative genes associated with paraquat resistance in goosegrass (Eleusine indica L.) [J]. PLoS One,2014,9(6):e99940.
[7]Han B,Ma X D,Cui D,et al. Comprehensive evaluation and analysis of the mechanism of cold tolerance based on the transcriptome of weedy rice seedlings[J]. Rice,2020,13(1):12.
[8]Kress W J,Erickson D L. A two-locus global DNA barcode for land plants:the coding rbcL gene complements the non-coding trnH-psbA spacer region[J]. PLoS One,2007,2(6):e508.
[9]張韵洁,李德铢. 叶绿体系统发育基因组学的研究进展[J]. 植物分类与资源学报,2011,33(4):365-375.
[10]王荣升,魏 鑫,曹立荣,等. 基于叶绿体基因多样性的中国水稻起源进化研究[J]. 植物遗传资源学报,2011,12(5):686-693.
[11]邱 涛,刘孝伟,唐 津,等. 玉米CMS-C同质异核、同核异质系叶绿体基因组比较分析[J]. 四川农业大学学报,2019,37(1):1-7.
[12]杨新泉,宋 星,杜金昆,等. 六倍体小麦(AABBDD)及其近缘种属野生二粒小麦和粗山羊草叶绿体SSR遗传差异研究[J]. 中国农业科学,2007,40(7):1324-1330.
[13]程 慧,葛春峰,张 红,等. 果树叶绿体基因组测序及系统发育研究进展[J]. 核农学报,2018,32(1):58-69.
[14]李泳潭,张 军,黄亚丽,等. 杜梨叶绿体基因组分析[J]. 园艺学报,2020,47(6):1021-1032.
[15]杨 芳. 七里香蔷薇叶绿体基因组测序及结构分析[J]. 基因组学与应用生物学,2019,38(8):3586-3594.
[16]叶晓倩,赵忠辉,朱全武,等. 茶树 “龙井43” 叶绿体基因组测序及其系统进化[J]. 浙江大学学报(农业与生命科学版),2014,40(4):404-412.
[17]吴丽新,龚 洵,潘跃芝. 姜状三七遗传多样性和遗传分化研究[J]. 植物科学学报,2020,38(4):525-535.
[18]乔永刚,贺嘉欣,王勇飞,等. 药用植物苦参的叶绿体基因组及其特征分析[J]. 药学学报,2019,54(11):2106-2112.
[19]杨 斌,孟庆瑶,张 凯,等. 孑遗濒危植物矮扁桃叶绿体全基因组特征分析及亲缘關系鉴定[J]. 植物研究,2020,40(5):686-695.
[20]于 涛,张宇阳,高 健,等. 极小种群濒危植物盐桦叶绿体基因组特征分析[J]. 林业科学,2019,55(2):41-49.
[21]李裕华,任永康,赵兴华,等. 禾本科主要农作物叶绿体基因组研究进展[J]. 生物技术通报,2020,36(11):112-121.
[22]林张翔. 稗草叶绿体基因组测序及其分子标记开发与利用[D]. 杭州:浙江大学,2015.
[23]姚 南. 基于叶绿体基因组序列多态性的杂草稻起源以及杂草稻的落粒性研究[D]. 重庆:重庆大学,2015.
[24]朱国忠,戴伟民,陈晓锋,等. 基于基因组数据库的杂草稻叶绿体分子标记开发[J]. 南京农业大学学报,2015,38(2):240-247.
[25]王梦醒,詹豪爽,吕萌荔,等. 粗山羊草叶绿体基因RNA 编辑位点的鉴定与分析[J]. 麦类作物学报,2014,34 (10):1341-1349.
[26]何懿菡,韩立敏,刘玉萍,等. 鼠尾草叶绿体基因组序列分析[J]. 植物研究,2017,37(4):572-578.
[27]Downie S R,Katz-Downie D S,Wolfe K H,et al. Structure and evolution of the largest chloroplast gene (ORF2280):internal plasticity and multiple gene loss during angiosperm evolution[J]. Current Genetics,1994,25(4):367-378.
[28]唐 萍,阮秋燕,彭 程. 禾本科植物叶绿体基因组结构的系统进化研究[J]. 中国农学通报,2011,27(30):171-176.
[29]冯克伟. 禾本科叶绿体基因组的比较与系统发育分析[D]. 杨凌:西北农林科技大学,2014.
[30]王红春,徐 蓬,孙钰晨,等. 江苏省稻田杂草的发生现状与防控建议[J]. 杂草学报,2019,37(4):1-5.
猜你喜欢
天中学刊(2022年4期)2022-11-08
化学与生物工程(2022年3期)2022-03-26
作物研究(2021年4期)2021-01-03
药学研究(2020年3期)2020-12-17
智慧健康(2018年26期)2018-02-09
首都食品与医药(2017年23期)2017-10-20
中国塑料(2016年2期)2016-06-15
兽医导刊(2016年12期)2016-05-17
应用海洋学学报(2015年3期)2015-11-22
中国煤层气(2015年4期)2015-08-22
