
木尔坦棉花曲叶病毒在新疆的发生分布及烟粉虱带毒检测
2022-06-07 11:26高国龙张兴旺姜子健黄家风都业娟
植物保护 2022年3期
关键词:棉花
高国龙 张兴旺 姜子健 黄家风 都业娟

摘要 為了明确木尔坦棉花曲叶病毒cotton leaf curl Multan virus (CLCuMuV)在新疆的发生分布,对采自新疆13个地(州)的1 166份蜀葵、苘麻、棉花疑似病株进行CLCuMuV分子鉴定和带毒检测,并对3种寄主上同期采集的1 588头烟粉虱进行隐种鉴定和带毒检测。结果显示,CLCuMuV DNA-A组分及其伴随的DNAβ卫星分子均可侵染蜀葵、苘麻、棉花;基于DNA-A及DNAβ组分的进化分析表明,新疆3种寄主上发生的CLCuMuV与国内已报道的CLCuMuV 进化关系一致;蜀葵、苘麻、棉花上CLCuMuV带毒率存在差异,分别为27.87%、33.33%和14.95%;MEAM1和MED隐种为新疆烟粉虱主要种群,分别占26.70%和66.44%,带毒率分别为21.46%和17.63%,说明MED隐种为新疆烟粉虱优势种群,2个隐种均可携带CLCuMuV且带毒率相近;MEAM1隐种在蜀葵、苘麻、棉花上带毒率分别为21.39%、0和28.21%,MED隐种分别为19.61%,8.47%和11.18%,表明2个隐种在不同寄主上的带毒率存在差异。本研究明确了新疆地区侵染蜀葵、苘麻和棉花的CLCuMuV发生及分布情况,并明确新疆烟粉虱主要种群MEAM1和MED 隐种在3种寄主上的分布及带毒情况,为新疆棉区防控CLCuMuV发生严重危害提供科学依据。
关键词 CLCuMuV;棉花;蜀葵;苘麻;烟粉虱
中图分类号: S435.61
文献标识码: A
DOI: 10.16688/j.zwbh.2021154
Abstract In order to determine the distribution of cotton leaf curl Multan virus (CLCuMuV) in Xinjiang, 1 166 suspected plants of Alcea rosea, Abutilon theophrasti and cotton collected from 13 prefectures (autonomous prefectures) in Xinjiang were investigated by molecular identification and CLCuMuV detection, and the total of 1 588 Bemisia tabaci were also collected from the three hosts to determine the cryptic species of B.tabaci and their viruliferous rate of CLCuMuV by PCR. The results showed that A.rosea, A.theophrasti and cotton could be infected by CLCuMuV DNA-A and its beta satellite DNAβ. CLCuMuV DNA-A and its DNAβ from the three hosts in Xinjiang were closely related to the CLCuMuV isolates from other provinces in China based on phylogenetic tree analysis. The rate of CLCuMuV infection in A.rosea, A.theophrasti and cotton was 27.87%, 33.33% and 14.95%, respectively. The population of B.tabaci in Xinjiang was mainly comprised of MEAM1 and MED cryptic species. MEAM1 accounted for 26.70% with 21.46% viruliferous rate, and MED accounted for 66.44% with 17.63% viruliferous rate, indicating that MED cryptic species is the dominant population of B.tabaci in Xinjiang. Both MEAM1 and MED could carry CLCuMuV with similar viruliferous rate, the viruliferous rate of MEAM1 in A.rosea, A.theophrasti and cotton was 21.39%, 0 and 28.21%, respectively, and that of MED was 19.61%, 8.47% and 11.18%, respectively. The results showed that the viruliferous rates of MEAM1 and MED on different hosts were different. In conclusion, the occurrence and distribution of CLCuMuV in A.rosea, A.theophrasti and cotton in Xinjiang were determined, and the distribution and viruliferous rates of MEAM1 and MED on the three hosts were also determined, which provides scientific basis for the prevention and control of CLCuMuV in Xinjiang.69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
Key words CLCuMuV;cotton;Alcea rosea;Abutilon theophrasti;Bemisia tabaci
棉花曲叶病cotton leaf curl disease(CLCuD)是棉花生长期的毁灭性病害,不仅造成棉纤维长度、细度等品质下降,还严重影响产量,造成的产量损失可达10%~90%[1]。该病害是由烟粉虱Bemisia tabaci(Gennadius)传播的双生病毒科Geminiviridae菜豆金色花叶病毒属Begomovirus的多种病毒引起,通常统称为棉花曲叶病毒cotton leaf curl virus(CLCuV)。CLCuV可侵染其他寄主,并通过烟粉虱传播到棉花上[1],因此烟粉虱和其他寄主的分布和带毒情况对该病害的传播及流行具有重要作用。
烟粉虱是由多个隐种组成的物种复合体,也是传播CLCuV的唯一昆虫介体[2]。其中Middle East-Asia Minor 1(MEAM1)和Mediterranean(MED)2个隐种入侵性强、分布广泛,在我国最为常见且为害严重[3-4]。新疆烟粉虱隐种鉴定显示2010年前均为MEAM1隐种[4-5],自2011年首次报道MED隐种入侵新疆后[6],不同区域烟粉虱种群组成呈动态变化:南疆和田地区烟粉虱种群由MEAM1隐种发展为MED隐种和MEAM1隐种混发,以MED隐种为优势种群[7];东疆鄯善县和北疆克拉玛依市两区域由MED隐种发展为2个隐种混发,但以MED隐种为优势种群[7-8];石河子市检测结果表明,由MEAM1隐种转变为2个隐种混发,无明显优势种群[7-9];乌鲁木齐市烟粉虱仅检测到MED隐种[7-9];库尔勒市检测到Asia II 7隐种[9]。由于不同隐种烟粉虱对CLCuV的传播能力差异显著,因此及时监控烟粉虱隐种动态变化趋势及带毒情况是预防该病害暴发流行的有力措施。
CLCuV严重危害棉花的同时,还能以多种观赏花卉和杂草作为寄主。苏丹已报道锦葵科Malvaceae的棉属Gossypium、木槿属Hibiscus、悬铃花属Malvaviscus、蜀葵属Alcea、苘麻属Abutilon等9个属植物可受到CLCuV危害[10];巴基斯坦在茄科Solanaceae、葫芦科Cucurbitaceae、旋花科Convolvulaceae、豆科Fabaceae及锦葵科的蜀葵Alcea rosea (L.) Cavan.、苘麻Abutilon theophrasti Medicus.、木槿Hibiscus syriacus L.等寄主上观察到了CLCuV危害的曲叶症状[11];印度则报道了通过传毒试验和ELISA检测,明确菜豆、辣椒、番茄和烟草中存在CLCuV[12]。上述报道明确了CLCuV除了危害棉花之外,还存在广泛的寄主,尤其是锦葵科中多个属均可受到侵染,但受限于早期试验条件的限制,并未明确病毒的具体种名。
木尔坦棉花曲叶病毒cotton leaf curl Multan virus (CLCuMuV)是CLCuV中危害最重、分布最廣的种类[13],由DNA-A组分及其伴随的DNAβ卫星分子组成。DNAβ只能依赖辅助病毒进行复制,对病毒侵入寄主后的显症具有重要作用[14]。CLCuMuV是导致20世纪90年代巴基斯坦CLCuD大流行的主要病原[14],并在印度[15]、菲律宾[16]等国家都有危害。我国继2006年在广东省首次发现CLCuMuV侵染朱槿H.rosa-sinensis L.后[17],广西、福建、江苏、海南、云南、内蒙古和新疆等地均报道该病毒的入侵[9]。寄主除棉花外[18-19],还包括秋葵属Abelmoschus的黄秋葵A.esculentus (L.) Moench[20]、悬铃花属的垂花悬铃花M.arboreus var. penduliflorus [21]、木槿属的朱槿[22]和红麻H.cannabinus L.[23]等锦葵科寄主,但无侵染蜀葵属和苘麻属寄主的报道。蜀葵是锦葵科蜀葵属多年生观赏性花卉植物,在新疆城市和农村的房前屋后及田间地头都有成片的分布;苘麻为锦葵科蜀葵属草本植物,在世界范围广泛分布[24],也是棉田中最为常见的杂草,CLCuMuV能否侵染二者,与其能否在新疆棉田扩散危害直接相关。
新疆作为我国重要棉区,2019年棉花产量占全国84.90%,伴随携毒朱槿苗木和烟粉虱的跨区调运,CLCuMuV向新疆棉区传播的潜在风险急剧增大;CLCuMuV潜在寄主蜀葵和苘麻等在新疆的广泛分布为病毒的扩散和持续危害提供了有利条件;而烟粉虱隐种组成动态变化又为病害的有效监控增加了难度。因此,本研究对全疆主要棉区开展棉花和其他寄主CLCuMuV侵染情况的普查,并对烟粉虱隐种的发生分布及带毒情况进行检测,明确该病毒和带毒烟粉虱隐种的发生情况,以期为新疆棉花曲叶病的早期监测防控提供科学依据。
1 材料与方法
1.1 材料
供试蜀葵、苘麻、棉花样品共1 166份,为2017 年至2020年采自新疆东疆、南疆和北疆13个地(州):其中778份为表现植株矮缩、叶片卷曲皱缩等症状的蜀葵、苘麻疑似病株叶片;388份棉花样品无典型症状,但均采自表现曲叶症状的蜀葵、苘麻附近。供试烟粉虱1 588头为同期采集自蜀葵、苘麻和棉花。
1.2 方法
1.2.1 DNA提取
植物样品DNA的提取使用天根生化科技(北京)有限公司的新型植物基因组DNA提取试剂盒(DP305-03),并将DNA于-20℃保存。烟粉虱总DNA的提取[25],将单头烟粉虱置于1.5 mL的离心管中,吸取20 μL裂解液[10 mmol/L Tris (pH 8.4),50 mmol/L氯化钾,0.2% 明胶,0.45% 吐温-20,0.45% NP-40,60 mg/mL蛋白酶K]并用研棒将其充分研磨匀浆,吸取20 μL裂解液清洗研棒并将清洗液与匀浆混合,65℃水浴1 h,接着沸水浴10 min,经短暂离心后即可用于PCR反应,DNA保存于-20℃。69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
1.2.2 CLCuMuV检测及烟粉虱隐种鉴定
分别利用2对特异引物 AF/AR[19]和βF/DNA-βR对CLCuMuV DNA-A及伴随的DNAβ卫星分子进行PCR检测;利用烟粉虱 mtCOⅠ 基因通用引物 C1-J-2195/R-BQ-2819[26]进行烟粉虱鉴定,再通过B-F/B-R、Q-F/Q-R[27]对烟粉虱MEAM1和MED隐种进行鉴定(表1)。
1.2.3 CLCuMuV DNA-A及DNAβ组分的克隆测序及序列分析
分别在蜀葵、苘麻和棉花样品中选取代表性病株,以其基因组DNA为模板,利用CLCuMuV特异性引物AF/AR和βF/DNA-βR对CLCuMuV DNA-A及伴随的DNAβ卫星分子进行PCR扩增(表1),克隆并测序。序列测定由生工生物工程(上海)股份有限公司完成,测序结果使用DNAMAN软件进行序列拼接及相似性分析,利用MEGA-X软件的邻接法(neighbor-joining)完成聚类分析及进化树构建。
2 结果与分析
2.1 侵染蜀葵、苘麻和棉花的CLCuMuV的分子鉴定
首先对蜀葵、苘麻疑似病株中的CLCuMuV进行分子鉴定,结果显示,4株蜀葵样品和3株苘麻样品均能扩增到CLCuMuV DNA-A 832~834 bp的目标片段,7个片段之间的核苷酸序列相似性为99.16%~100.00%;BLAST检索结果显示,7个序列与CLCuMuV分离物DNA-A序列的相似性为89.95%~99.88%。表明疑似感病蜀葵和苘麻均被CLCuMuV侵染。再对2株蜀葵和2株苘麻病株中CLCuMuV伴随的DNAβ分子进行鉴定,均能扩增到432~433 bp的目标DNAβ片段,4个片段之间的核苷酸序列相似性为99.08%~100.00%,与CLCuMuV DNAβ序列的相似性为85.19%~99.77%,表明侵染蜀葵和苘麻的CLCuMuV伴随有DNAβ分子。
对20份采集自感病蜀葵和苘麻附近的棉花样品进行CLCuMuV鉴定,结果从8株棉花中扩增到CLCuMuV DNA-A目标片段,序列大小均为832 bp,且核苷酸序列相似性为99.16%~100.00%;8个序列与CLCuMuV分离物DNA-A序列相似性为91.95%~100.00%,表明8株棉花均被CLCuMuV侵染;对其中3份样品中CLCuMuV伴随的DNAβ分子进行鉴定,均可获得约400 bp 的预期目标片段,测序结果表明其均为433 bp,3个片段之间的序列相似性为99.54%~99.77%,与CLCuMuV卫星分子DNAβ的序列的相似性为85.19%~99.77%,表明侵染棉花的CLCuMuV伴隨有DNAβ卫星分子。进一步比对分析发现,棉花中的CLCuMuV与蜀葵和苘麻病株中DNA-A序列间的相似性均超过98.56%,其伴随的DNAβ卫星分子序列间的相似性也均超过99.62%。
2.2 侵染蜀葵、苘麻和棉花的CLCuMuV DNA-A及DNAβ组分进化分析
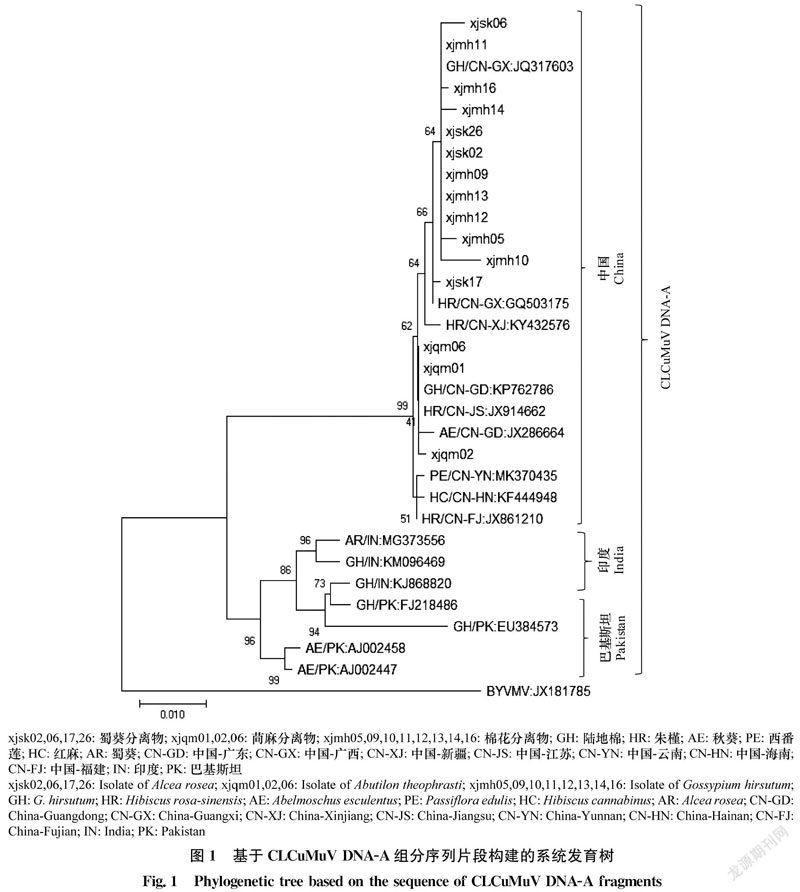
基于蜀葵、苘麻和棉花中获得的CLCuMuV DNA-A 序列构建系统发育树,结果如图1所示,所有CLCuMuV聚类在一个大的进化分支上,中国来源的CLCuMuV聚在一个亚分支上,其中新疆蜀葵3个分离物xjsk02、xjsk06和xjsk26及8个棉花分离物xjmh05、xjmh09、xjmh10、xjmh11、xjmh12、xjmh13、xjmh14和xjmh16与侵染广西棉花的CLCuMuV(登录号:JQ317603)进化关系最近;蜀葵分离物xjsk17与侵染广西朱槿的CLCuMuV进化关系最近;苘麻分离物xjqm01、xjqm02和xjqm06与侵染广东棉花、秋葵及江苏朱槿的CLCuMuV进化关系最近。而来自巴基斯坦、印度的CLCuMuV则聚类在另外1个亚分支上。以上结果表明,新疆蜀葵、棉花上发生的CLCuMuV与广西棉花上的CLCuMuV具有相同的进化关系;新疆苘麻上发生的CLCuMuV与广东和江苏锦葵科植物上的CLCuMuV具有相同的进化关系。
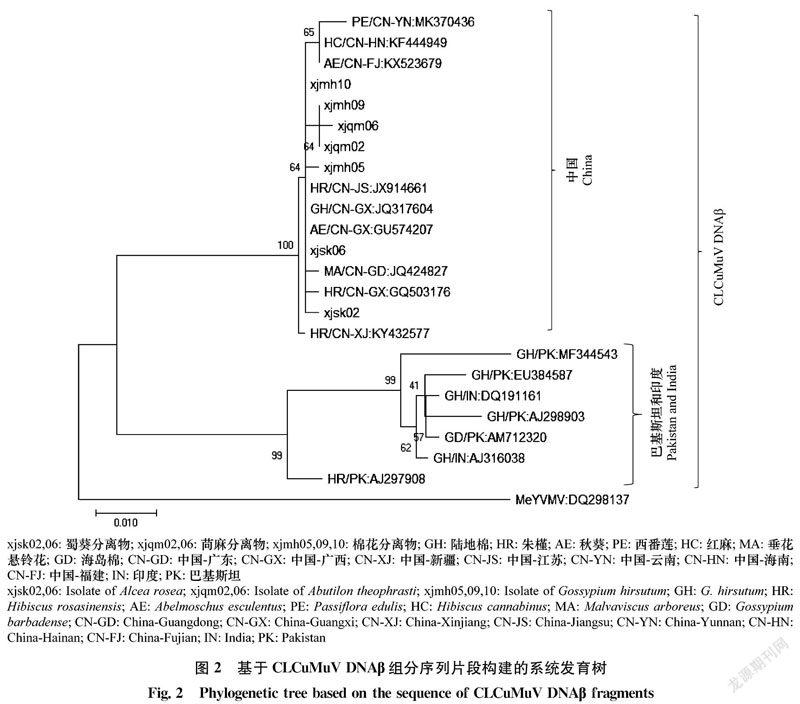
基于蜀葵、苘麻和棉花中获得的CLCuMuV DNAβ 序列构建系统发育树,结果如图2所示,所有CLCuMuV伴随的DNAβ聚类在一个大的进化分支上,中国来源的CLCuMuV伴随的DNAβ聚在一个亚分支上,其中新疆蜀葵2个分离物xjsk02、xjsk06伴随的DNAβ与侵染广西朱槿和侵染广东垂花悬铃花的CLCuMuV伴随的DNAβ进化关系最近;苘麻分离物xjqm02、xjqm06伴随的DNAβ和棉花分离物xjmh05、xjmh09及xjmh10与侵染江苏朱槿和侵染广西棉花的CLCuMuV伴随的DNAβ进化关系最近。而来自巴基斯坦、印度的CLCuMuV伴随的DNAβ则聚类在另一个亚分支上。以上结果表明,新疆蜀葵、苘麻和棉花上发生的DNAβ分子在进化上与其辅助病毒CLCuMuV具有一致性,即新疆发生的CLCuMuV及其卫星分子DNAβ与来源于广东、广西及江苏的锦葵科植物上的CLCuMuV及其卫星DNAβ具有相同的进化关系。
2.3 蜀葵、苘麻及棉花携带CLCuMuV的检测结果
为了明确CLCuMuV在全疆的发生分布,对采集自新疆13个地州的1 166份样品进行PCR带毒检测,结果显示(表2),276份样品感染CLCuMuV,且均伴随DNAβ卫星分子,带毒率为23.67%。对3种寄主的带毒情况进行分析,757份蜀葵样品中211份样品检测到病毒,带毒率为27.87%,其中东疆18份样品中,15份检测到病毒,检出率为83.33%;北疆214份样品中,105份检测到病毒,检出率为49.07%;南疆525份样品中,91份检测到病毒,检出率为17.33%。21份苘麻样品中7份检测到病毒,检出率为33.33%,其中南疆13份样品中,4份检测到病毒,带毒率为30.77%;北疆8份样品中,3份检测到病毒,检出率为37.50%。388份棉花样品中58份检测到CLCuMuV,检出率为14.95%,其中东疆16份样品中,10份检测到病毒,检出率为62.50%;南疆22份样品中,5份检测到病毒,检出率为22.73%;北疆350份样品中,43份检测到病毒,检出率为12.29%。上述结果表明,蜀葵和苘麻带毒率分别为27.87%和33.33%,均高于棉花14.95%的带毒率;蜀葵带毒率表现为东疆>北疆>南疆,苘麻带毒率表现为北疆>南疆,棉花带毒率表现为东疆>南疆>北疆。69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
2.4 蜀葵、苘麻及棉花上烟粉虱隐种及CLCuMuV带毒率的检测
利用烟粉虱 mtCOⅠ 基因片段通用引物对病株上的粉虱进行种类鉴定,明确病株上采集的1 588头粉虱均为烟粉虱;进而利用隐种特异性引物对烟粉虱进行隐种鉴定,结果显示MEAM1隐种为424头,占26.70%,MED隐种1 055头,占66.44%,另有109头未确定隐种,占6.86%(表3)。对烟粉虱带毒情况进行检测,结果显示,MEAM1隐种带毒率为21.46%,东疆、南疆和北疆的带毒率分别为18.75%、7.98%和40.59%;MED隐种带毒率为17.63%,东疆、南疆和北疆的带毒率分别为13.45%、18.32%和16.83%;未确定种烟粉虱均不带毒。对3种寄主上的烟粉虱隐种及其带毒率进行分析,蜀葵上1 303头烟粉虱,检测到242头携带CLCuMuV,带毒率为18.57%,其中MEAM1隐种占28.70%,带毒率为21.39%,MED隐种占63.39%,带毒率为19.61%;苘麻上73头烟粉虱中,MED隐种占80.82%,带毒率为8.47%;MEAM1隐种占15.07%,未检测到携带CLCuMuV;棉花上212头烟粉虱中,30头检测到病毒,带毒率为14.15%,其中MEAM1隐种占18.40%,带毒率为28.21%,MED隐种占80.19%,带毒率为11.18%。上述结果表明,烟粉虱MEAM1和MED隐种均可携带CLCuMuV,平均带毒率差异不大;但从区域分布来看,MEAM1隐种的带毒率在东疆(18.75%)、南疆(7.98%)和北疆(40.59%)差异较大,而MED隐种在上述三个区域带毒率差异不大;从烟粉虱寄主来看,蜀葵上的烟粉虱带毒率(18.57%)高于苘麻(6.85%)和棉花(14.15%)的带毒率。
3 结论与讨论
本研究针对CLCuMuV入侵新疆的寄主开展了研究,明确CLCuMuV可侵染蜀葵和苘麻,并伴随DNAβ卫星分子,这是我国首次报道。分子检测结果表明,蜀葵带毒率为27.87%,苘麻带毒率为33.33%,说明CLCuMuV通过花卉携带传入新疆后,已经在东疆、南疆和北疆借助多年生花卉及棉田杂草定殖下来,成为田间扩散的初侵染源。
对感病寄主附近的棉花进行CLCuMuV分子鉴定,明确CLCuMuV能够侵染棉花,并伴随卫星分子DNAβ,感病寄主附近的棉花带毒率为14.95%。因目前检测的感病棉花均无明显的受害症状,表明新疆棉花隐症带毒。对于隐症带毒是否与棉花品种有关,或是与病毒侵入时期过晚有关,还有待深入研究。
烟粉虱是CLCuMuV的唯一昆虫传播介体。本研究对感病蜀葵、苘麻、棉花上采集的烟粉虱进行隐种鉴定及带毒检测,结果显示,新疆烟粉虱主要由MEAM1和MED隐种组成,MED隐种为优势种群;MEAM1隐种带毒率为21.46%,MED隐种带毒率17.63%。虽然在不同的地理区域烟粉虱隐种的分布及带毒率存在差异,但二者均具有潜在的传毒风险。林林等的研究表明MEAM1可将CLCuMuV传播到棉花上[28],Pan等的研究表明MED隐种具有将CLCuMuV传到烟草上的能力[29]。鉴于本研究在田间棉花植株上已检测到CLCuMuV的侵染,关于MEAM1隐种和MED隐种传毒能力还有待进一步深入研究。
作者前期的调查发现,在新疆多个地区的花卉植物朱槿携带CLCuMuV,因气候寒冷,朱槿在新疆主要以室内盆栽模式进行种植,不利于烟粉虱传播CLCuMuV,而蜀葵属于多年生花卉,因其花色艳丽,易于种植,是新疆各地主要的庭院花卉;苘麻作为棉田主要杂草,在新疆亦有广泛分布。已有的研究表明,田間杂草就像双生病毒Begomovirus的储存库,可以作为其生存和传播的寄主或替代寄主,能同时被多种该属病毒及卫星复合侵染从而增加病毒重组的几率[30-31]。本研究从田间症状和分子检测水平进一步明确了CLCuV中的CLCuMuV可侵染蜀葵和苘麻。棉花为一年生喜温草本植物,在寒冷的冬季不能生长,而中间寄主不仅为病毒提供越冬场所,并为次年棉花生长期烟粉虱传播病毒提供毒源,还可借远距离调运将病毒及烟粉虱传播到新的区域,因此中间寄主在棉花曲叶病的传播中起重要作用,务必引起重视。
参考文献
[1] MAHARSHI A,YADAV N K,KUMAR P,et al. An overview on cotton leaf curl disease: An emerging potential threat to cotton [J]. International Journal of Current Microbiology Applied Sciences,2017,6(10): 2154-2162.
[2] 刘银泉,刘树生. 烟粉虱的分类地位及在中国的分布[J]. 生物安全学报,2012,21(4): 247-255.
[3] LIU Shusheng,COLVIN J,DE BARRO P. Species concepts as applied to the whitefly Bemisia tabaci systematics: how many species are there? [J]. Journal of Integrative Agriculture,2012,11(2): 176-186.
[4] TENG Xi,WAN Fanghao,CHU Dong. Bemisia tabaci biotype Q dominates other biotypes across China [J]. Florida Entomologist,2010,93(3): 363-368.
[5] 罗晨,姚远,王戎疆,等. 利用mtDNA COⅠ基因序列鉴定我国烟粉虱的生物型[J]. 昆虫学报,2003,45(6): 759-763.69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
[6] 段晓东,马丽娟,姚正培,等. 新疆地区烟粉虱类群mtDNA COI基因序列分析[J]. 生物安全学报,2011,20(1): 50-55.
[7] 韩畅,张兴旺,高国龙,等. 新疆番茄黄化曲叶病毒与烟粉虱隐种的区域分布检测[J]. 石河子大学学报(自然科学版),2020,38(2): 160-165.
[8] 曹骞,李晶,买热木古丽·克依木,等. 新疆地区烟粉虱生物型的区域分布及其携带的番茄黄化曲叶病毒检测[J]. 昆虫学报,2013,56(6): 652-664.
[9] 陈婷,汤亚飞,何自福,等. 我国朱槿曲叶病毒病及其传播介体烟粉虱分布调查[J]. 南方农业学报,2020, 51(11): 2697-2705.
[10]NUR E E,SALIH H S A. Cotton leaf curl virus disease [J]. International Journal of Pest Management,1970,16(1): 121-131.
[11]SINGH J,SOHI A S,MANN H S,et al. Studies on whitefly Bemisia tabaci (Genn.) transmitted cotton leaf curl disease in Punjab [J]. Journal of Insect Science,1994,7: 194-198.
[12]NATESHAN H M,MUNIYAPPA V,SWANSON M M,et al. Host range,vector relations and serological relationships of cotton leaf curl virus from southern India [J]. Annals of Applied Biology,1996, 128(2): 233-244.
[13]BRIDDON R W,MARKHAM P G. Cotton leaf curl virus disease [J]. Virus Research,2000,71(1/2): 151-159.
[14]ZHOU Xueping,LIU Yule,ROBINSON D J,et al. Four DNA-A variants among Pakistani isolates of cotton leaf curl virus and their affinities to DNA-A of geminivirus isolates from Okra [J]. The Journal of General Virology,1998,79(4): 915-923.
[15]ZAFFALON V,MUKHERJEE S,REDDY V,et al. A survey of geminiviruses and associated satellite DNAs in the cotton-growing areas of northwestern India [J]. Archives of Virology,2012,157(3): 483-495.
[16]SHE Xiaoman,TANG Yafei,HE Zifu,et al. Molecular characterization of cotton leaf curl Multan virus and its associated betasatellite infecting Hibiscus rosa-sinensis in the Philippines [J]. Journal of Plant Pathology,2017, 99(3): 765-768.
[17]毛明杰,何自福,虞皓,等. 侵染朱槿的木爾坦棉花曲叶病毒及其卫星DNA全基因组结构特征[J]. 病毒学报, 2008,24(1): 64-68.
[18]CAI Jianhe,XIE Ke,LIN Lin,et al. Cotton leaf curl Multan virus newly reported to be associated with cotton leaf curl disease in China [J]. Plant Pathology,2010,59(4): 794-795.
[19]汤亚飞,何自福,杜振国,等. 木尔坦棉花曲叶病毒及其伴随的β卫星分子复合侵染引起广东棉花曲叶病[J]. 中国农业科学,2015,48(16): 3166-3175.
[20]董迪,何自福,柴兆祥. 广东黄秋葵黄脉曲叶病样中检测到烟粉虱传双生病毒[J]. 植物保护,2010,36(1): 65-68.
[21]汤亚飞,何自福,杜振国,等. 侵染垂花悬铃花的木尔坦棉花曲叶病毒分子特征研究[J]. 植物病理学报,2013, 43(2): 120-127.
[22]章松柏,夏宣喜,张洁,等. 福州市发生由木尔坦棉花曲叶病毒引起的朱槿曲叶病[J]. 植物保护,2013,39(2): 196-200.
[23]汤亚飞,何自福,杜振国,等. 海南红麻曲叶病的病原检测与鉴定[J]. 植物病理学报,2015,45(6): 561-568.
[24]YETURUA S,JENTZSCHAB P V,CIOBOT V,et al. Handheld Raman spectroscopy for the early detection of plant diseases: Abutilon mosaic virus infecting Abutilon sp. [J]. Analytical Methods,2016,8(17): 3450-3457.69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
[25]秦丽,王佳,邴孝利,等. 利用mtCOI PCR-RFLP技术鉴定中国境内九个烟粉虱隐种[J]. 昆虫学报,2013,56(2): 186-194.
[26]刘国霞,高长生,付海滨,等. 烟粉虱Q与B隐种mtCOI基因的遗传变异及其对利用CAPS标记进行隐种鉴别的影响[J]. 昆虫学报,2014,57(10): 1238-1244.
[27]SHATTERS J,ROBERT G,POWELL C A,et al. Improved DNA barcoding method for Bemisia tabaci and related Aleyrodidae: Development of universal and Bemisia tabaci biotype-specific mitochondrial cytochrome c oxidase I polymerase chain reaction primers [J]. Journal of Economic Entomology,2009,102(2): 750-758.
[28]林林,蔡健和,羅恩波,等. 南宁市朱槿曲叶病毒病病原分子鉴定和寄主范围研究[J]. 植物保护,2011,37(4): 44-47.
[29]PAN Lilong, CUI Xiyun, CHEN Qunfang, et al. Cotton leaf curl disease: which whitefly is the vector?[J]. Phytopathology, 2018, 108(10): 1172-1183.
[30]RODRGUEZ-PARDINA P E,HANADA K,LAGUNA I G,et al. Molecular characterisation and relative incidence of bean- and soybean-infecting begomoviruses in northwestern Argentina [J]. Annals of Applied Biology,2011, 158(1): 69-78.
[31]SOUZA T A,MACEDO M A,ALBUQUERQUE L C,et al. Host range and natural infection of tomato chlorosis virus in weeds collected in Central Brazil [J]. Tropical Plant Pathology,2020,45(1): 84-90.
(责任编辑:田 喆)69033D07-FF88-4A2B-9A1F-6EC5CF4241CA
猜你喜欢
少儿科学周刊·儿童版(2022年10期)2022-06-27
少儿科学周刊·儿童版(2022年10期)2022-06-27
小天使·一年级语数英综合(2022年4期)2022-04-28
作文·小学低年级(2021年8期)2021-11-02
学苑创造·A版(2017年9期)2017-09-25
小溪流(画刊)(2017年1期)2017-03-16
学生天地·小学低年级版(2016年7期)2016-05-14
红领巾·萌芽(2015年7期)2015-09-10
红领巾·萌芽(2015年6期)2015-08-14
小天使·一年级语数英综合(2014年7期)2014-06-26
