
崇明东滩围垦区不同土地利用类型对土壤呼吸及其组分的影响
2022-06-23 05:31何晶仲启铖张桂莲王开运吴悦琦贺贵书李德志彭娜娜
华东师范大学学报(自然科学版) 2022年3期
何晶 仲启铖 张桂莲 王开运 吴悦琦 贺贵书 李德志 彭娜娜







摘要:在崇明东滩围垦区,采用Licor-8100A土壤碳通量观测系统及分层去根法,连续测定5种土地利用类型(芦苇湿地、白茅湿地、幼龄林、中龄林和农田)的土壤呼吸、异养呼吸和自养呼吸,以及0~10cm表层土壤温度、体积含水率和电导率等环境因子,系统地比较了崇明东滩围垦区不同用地类型土壤呼吸及其组分的差异.结果表明:①幼龄林、中龄林和农田样地的土壤呼吸显著低于芦苇湿地和白茅湿地样地;②幼林龄、中林龄和农田样地的异养呼吸占比显著高于芦苇湿地和白茅湿地样地;③土壤呼吸及其组分与土壤温度呈显著的指数关系,但与土壤含水率和电导率的相关性较弱.与残存湿地相比,不同的农林利用方式显著降低了围垦区土壤呼吸,但大幅增加了异养呼吸,这可能说明土壤有机碳库在围垦20年后仍处于净损失状态.因此,需要采取有效措施进一步提升该区域土壤的碳固持能力.
关键词:土壤呼吸;土壤呼吸组分;滨海湿地围垦;土地利用类型
0引言
滨海湿地在碳固持和存储方面发挥着重要作用.按面积计算,与大多数陆地森林相比,滨海湿地是更有效的碳汇[1].滨海湿地生态系统中的植被和土壤储存了大量的“蓝碳”[2].据估测,全球范围内,仅滨海盐沼0~50cm土层就存储了近430Tg碳[3].随着河口海岸地区城市化进程的不断加速和人口增长及经济发展对土地需求量的大幅增加,大量滨海湿地被人工围垦以作为新生土地资源.土地利用方式的转变会影响土壤的环境因子,进而影响土壤呼吸[4].土壤呼吸是陆地生态系统碳循环的重要环节,是碳在陆地生态系统和大气生态系统之间进行转换的主要途径之一[5].通过土壤呼吸,土壤每年向大气排放的碳达到68~98Pg[6],因此,即便土壤呼吸速率变化微小,也会对碳循环造成较大的影响.目前有研究表明,当湿地转变成农田时,土壤呼吸会上升或下降[7-9],但滨海湿地围垦后转变成不同用地类型后,会对土壤呼吸产生怎样的具体影响,迄今研究很少.
土壤呼吸(SoilRespiration,RS)指未扰动土壤中产生CO2的所有代谢活动,主要由根系的自养呼吸(AutotrophicRespiration,RA)以及土壤微生物和动物的异养呼吸(HeterotrophicRespiration,RH)组成[10].有研究表明,土地利用类型的改变会影响RS[11],并且RS对土地利用类型的转变的响应差异很大,这是因为,不同土地类型的RS的组分RH和RA对土壤生物因子和非生物因子变化的响应不同.RH主要受微生物活性、土壤有机质含量、温度、湿度和电导率等的影响,而RA主要受细根生物量、土壤温度、养分有效性、C分配和林龄等的影响[12-13].Hu等[13]研究表明,天然林转变成人工竹林后,主要是RH上升导致了RS上升.Frank等[8]研究表明,草地转变成农田后,主要是RA下降导致了RS下降.因此,将RS分为RA和RH进行定量化研究,有助于从机制上理解土地利用类型的转变对RS的影响[14].另一方面,并不能直接认为RS上升就意味着土壤碳库损失增加,因为RS上升可能是由于初级生产力增加而导致根系获得更多碳分配,从而导致根系呼吸增加,因此,分不同组分对RS进行测量,不仅有助于细致分析滨海湿地围垦区转变成不同土地利用类型后土壤呼吸的基本特征和规律,也可以为评估滨海湿地围垦区在不同利用方式下土壤的碳源或碳汇性质,提供确凿的实证依据[15].
崇明东滩湿地地处长江口,是我国最大的河口冲积岛,盐沼湿地发育典型.长期以来,因人工围垦形成了大面积的滨海围垦湿地,而且这些围垦湿地被陆续转变成了不同的用地类型.为系统研究崇明东滩围垦区不同用地类型土壤呼吸及其组分,本文通过箱式法(LI-8100A)连续监测了芦苇湿地、白茅湿地、幼龄林、中龄林和农田这5种不同土地利用类型的土壤呼吸速率及其组分,探讨了土地利用类型的不同对土壤呼吸、自养呼吸和异养呼吸的影响,及其与土壤温度、含水率和电导率之间的关系,分析了不同用地类型对该区土壤碳循环的影响,以期为滨海湿地围垦区低碳土地利用管理提供理论依据和技术参考.
1材料与方法
1.1 研究区概况
研究区位于上海市崇明岛东滩湿地(31°38′N,121°58′E)(图1),是长江口规模最大、发育最完善的河口型湿地.该地区属于北亚热带海洋性季风气候,年平均气温为15.3℃,年平均降水量为1004mm.该地区土壤类型为滨海盐碱型砂壤土,以河口沉积物为母质.土壤盐度(psu)为0.1~5.0,pH值为8.2~9.2,土壤碳和养分含量相对较低[16].
该研究区域从1998年起,经历了大规模的湿地排水和农林利用.截至2018年年底,该地包含残存湿地斑块和多种农林利用类型.本研究选取5种土地利用類型,包括地下水位较高、以芦苇(Phragmitesaustralis)为优势种群的草本沼泽湿地(Phragmitesaustraliswetland,PW);地下水位较低、以白茅(Imperatacylindrica)为优势种的草本沼泽湿地(Imperatacylindricawetland,IW);树木栽种时间为5年的幼龄人工林(YoungForest,YF)[17],其乔木层为无患子(Sapindussaponaria)、朴树(Celtissinensis)林下种植花菜;树木栽种时间为15年的中龄人工林(Middle-ageForest,MF)[17],其乔木层为杉木(Cunninghamialanceolate)、朴树(Celtissinensis),下层灌木为夹竹桃(Neriumindicum)和伞房决明(Sennacorymbosa),以及轮作玉米-花菜的农田(Cropland,CP).
1.2 样地设置
2018年6月,在芦苇湿地、白茅湿地、幼龄林、中龄林和农田5种土地类型中,各选定20m×20m样地,在每种样地内,选择3块植被均匀、地形完整的小样地.在每个小样地内,按“s”形选定具有代表性的6个样点,其中有3个无去根样点和3个去根样点.去根样点和无去根样点相邻,样点之间的距离不小于5m.各土地类型上共选择18个样点.
1.3 土壤呼吸测定
2018年7月—2019年7月用LI-8100A土壤碳通量系统,测定各样地的土壤呼吸速率(RS)和异养呼吸速率(RH).每月在固定时间段9:30—11:30测定.在各样地上,选择60cm×60cm样点.每个样点上嵌入1个PVC土壤环(内径20cm,高10cm,型号SG-5,下同),用LI-8100A直接测定RS.在无去根样点旁,布置去根样点,向下0~10cm、10~20cm、20~40cm、40~60cm分层挖掘[18].在挖好的60cm×60cm×60cm坑中,按“十”字形,铺设两张100目尼龙网,以隔绝根系.取出的每层土壤过2mm筛,清除植物根系,然后,按顺序将土壤填回.同样,在每个样点中央,分别嵌入1个PVC土壤环.采用去根法结合LI-8100A,測定土壤RH值.RA值即为RS与RH的值之差(表1).
1.4 土壤环境因子测定
于每月测定土壤呼吸,同时使用土壤三参数速测仪(Wet-2,Delta-T,Cambridge,UK),测量土壤0~5cm处的温度、含水率、电导率.每个样点3次重复测量.
1.5 土壤理化性质测定
样方布设完成后,在正式实验前,取样测定本底土壤理化特征.采用环刀法测定土壤容重;采用WET型三参数分析仪测定土壤体积含水率.采集各样点土壤50g,风干后,于烘箱120℃下烘干24h,过100目筛,用元素分析仪(Elementar,modelVarianMacroCN,Hanau,Germany)测定全碳和全氮(表2).
1.6 数据处理与分析
采用MicrosoftExcel2019进行数据初步整理,采用SPSS13.0软件进行统计分析,显著性水平设置为0.05.采用重复测量方差分析,比较不同用地类型之间土壤温度、含水率、电导率的差异和不同用地类型下土壤呼吸及其组分之间的差异,以及不同用地类型、时间以及两者的交互作用对土壤呼吸及其组分的影响.采用偏相关分析方法分析土壤呼吸速率及其组分与环境因子的相关性.将土壤呼吸速率及其组分与环境因子作逐步回归.
土壤呼吸速率及其组分、温度、含水率、电导率的动态变化用OriginPro2019b进行绘制.土壤呼吸速率及其组分与温度的关系,采用指数函数拟合,见式(1)[19]:
式(1)中:R为土壤呼吸速率(μmol·m–2·s–1),T为土壤温度(℃),a为温度为0℃时的土壤呼吸速率,b为温度反应系数.
土壤呼吸的温度敏感性指数Q10依据式(2)[20]计算:
2结果与分析
2.1 不同土地利用类型的环境因子
由图2可知,不同用地类型样地的土壤温度均呈现显著的季节差异.最高温度出现在夏季7月,最低温度出现在冬季1月,但各样地之间的土壤温度没有显著差异.各样地土壤体积含水率的动态变化与土壤温度和降雨量有关,年均土壤含水率表现为PW((48.40±1.25)%)>IW((38.50±1.12)%)>CP((29.40±0.82)%)>YF((25.80±1.36)%)>MF((20.70±1.63)%).年均土壤电导率表现为CP((420.75±49.77)ms·m–1)>PW((359.83±22.20)ms·m–1)>IW((248.84±23.78)ms·m–1)>MF((127.06±8.65)ms·m–1)>YF((128.35±10.13)ms·m–1).
2.2 不同土地利用类型土壤呼吸速率及其组分
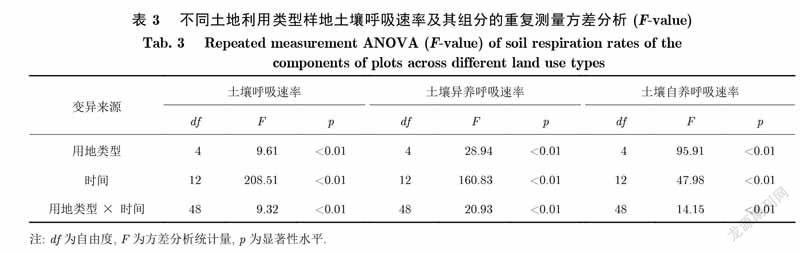
重复测量方差分析表明,样地的用地类型和用地时间以及两者的交互作用对土壤呼吸速率及其组分有显著影响(p<0.01)(表3).
由图3可知,在2018年7月至2019年7月,除农田外,各土地利用类型样地的土壤呼吸速率动态变化整体趋势是先降低后上升,呈单峰曲线;在9月,农田样地的土壤呼吸骤增,这可能与9月收获玉米重新翻耕土壤播种花菜有关.7月和8月人工林样地的RH显著高于残存湿地样地,人工林样地的RA显著低于残存湿地样地.PW样地((8.03±1.15)μmol·m–2·s–1)和IW样地((7.48±1.00)μmol·m–2·s–1)的年平均RS显著高于YF((6.48±0.82)μmol·m–2·s–1)、MF((6.53±0.86)μmol·m–2·s–1)、CP((5.08±0.60)μmol·m–2·s–1).YF((4.67±0.66)μmol·m–2·s–1)、MF((5.18±0.77)μmol·m–2·s–1)、CP((4.10±0.49)μmol·m–2·s–1)的年平均RH显著高于PW((1.98±0.22)μmol·m–2·s–1)和IW((2.41±0.31)μmol·m–2·s–1).PW((6.51±1.02)μmol·m–2·s–1)和IW((5.07±0.90)μmol·m–2·s–1)的年平均RA显著高于YF((1.81±0.25)μmol·m–2·s–1)、MF((1.35±0.21)μmol·m–2·s–1)、CP((1.69±0.20)μmol·m–2·s–1)(图3).
2.3 各用地类型土壤呼吸速率及其组分与环境因子的关系
偏相关分析表明(表4),各样地土壤呼吸速率及其组分与温度呈显著相关,除PW和MF外,样地土壤呼吸速率及其组分与含水率和电导率的相关性较弱.对样地土壤呼吸速率及其组分与温度进行指数拟合,除CP外,各用地类型样地的土壤呼吸速率与温度显著相关,可解释80%以上的变异;相较于其他用地类型样地,人工林样地(YF、MF)的RH与温度相关性较高,可解释80%以上的变异;相较于其他用地类型样地,残存湿地样地(PW、IW)的RA与温度相关性较高,可解释70%以上的变异.人工林样地的RH的Q10显著高于其他用地类型样地,残存湿地样地(PW、IW)的RA的Q10显著高于其他用地类型样地.经对数转换后的样地土壤呼吸速率及其组分与环境因子间的逐步回归结果表明,温度是影响样地土壤呼吸的主要因子,含水率影响较弱,电导率无显著影响.
3讨论
3.1 不同土地利用类型对土壤呼吸及其组分的影响
本文研究结果表明,不同用地类型样地的土壤呼吸速率有显著差异.围垦后的残存湿地样地(PW和IW)转变成农林利用的土地类型(YF、MF和CP)后,RS显著降低.相较于围垦后的芦苇湿地样地,白茅湿地样地的RS降低了7.0%,人工林样地降低了18.8%~19.4%,农田样地降低了27.9%(图3).这与Frank等[8]和Ouyang等[9]研究结果相似,但与Song等[7]研究结果相反:Song等[7]研究表明,裸露湿地转变成农田后,RS会增加,这可能是因为裸露湿地植被根系较少.然而,相较于PW的RH,IW、YF、MF和CP样地的RH分别增加了21.7%、135.9%、161.6%和107.7%(图3).这与李愈哲等[21]研究结果相似.这表明围垦后的残存湿地转变成农林用地后,将会加强土壤碳库的流失.本研究表明,土地利用类型的转变改变了RH和RA對RS的贡献率[13](图3).这与Huang等[22]研究结果相一致.PW和IW样地的土壤呼吸主要来自于自养呼吸,其RA对RS的贡献率分别为75.4%和67.7%,而YF、MF和CP样地的土壤呼吸主要来自于异养呼吸,其RH对RS的贡献率分别为72.1%、79.3%和70.8%.PW和IW样地RS的贡献率主要来自RA,因为芦苇和白茅地下根系发达,细根生物量大,而土壤呼吸速率与细根生物量呈正相关关系[23].YF、MF和CP样地的RS的贡献率主要来自RH,可能有以下几个原因:①新造林单位面积生产力低、根系不发达、根系呼吸低;②施肥会输入更多的有机碳,施肥使微生物的活性不受限于土壤有机碳的含量,翻耕、播种和除草会改变土壤结构,增加土壤的通气性,而良好的土壤通气环境更有利于微生物的活动[24-25];③PW和IW样地的土壤含水率显著高于YF、MF和CP样地,土壤含水量高限制了土壤与大气之间的气体交换,降低了土壤氧气浓度,从而限制了土壤微生物的呼吸[26],也阻碍了CO2的释放.
3.2 环境因子对不同土地类型土壤呼吸及其组分的影响
样地的不同用地类型会影响土壤呼吸及其组分.一部分原因是土地利用方式的改变会改变土壤的环境条件[4].本研究表明,样地土壤温度与土壤呼吸速率及其组分呈显著相关,揭示出温度是影响土壤呼吸速率及其组分的主要环境因子(表4和图4),这与Mathew等[27]、Raich等[28]和Wang等[29]的研究一致.图4表明,残存湿地样地RA的Q10是RH的2倍,这说明残存湿地样地的土壤呼吸变化,主要是由根系及根际微生物对温度变化的响应所决定[20].然而,人工林样地RH的Q10是RA的2倍,MF样地的RA与温度无相关性,这说明人工林样地的土壤呼吸变化主要是由土壤微生物代谢活动对温度变化的响应所决定[30].植物根系以及根际微生物利用的碳主要来源于地上部分的光合作用所同化的碳[31],而土壤微生物利用的碳主要来源于土壤存储的有机碳[32].本研究表明,相比于残存湿地样地RH的Q10,人工林样地的Q10有所增加.可见,在全球变暖的背景下,围垦后的湿地转变成人工林后,会导致土壤碳存储的减少,并向大气释放更多的CO2,由此将进一步加剧区域乃至全球气候变暖的趋势.CP样地的土壤呼吸与温度间的回归方程拟合优度低是因为CP样地受到人为干扰因素多(如施肥、除草、翻耕等),导致影响土壤呼吸的因素更为复杂(表5).
本研究表明,除MF样地外,各用地类型样地土壤含水率与土壤呼吸速率及其组分的相关性很小.Davison等[33]研究表明,土壤含水率对土壤呼吸的影响很小.当植物和微生物生长不受水分胁迫时,土壤CO2释放量主要受温度影响,而土壤含水率对土壤呼吸无显著影响.MF样地含水率显著低于其他用地类型.其含水率较低可能会限制植物和微生物的代谢活性,因此,其土壤呼吸速率及其组分与含水率间呈显著正相关关系.
盐度会对植物生长和土壤微生物活性产生有害影响[34-35],所以随着土壤盐度的提高,土壤呼吸会下降.然而本研究表明,除PW和MF样地外,各用地类型样地的电导率与土壤呼吸及其组分无显著相关关系.这可能是因为围垦区前期的排水洗盐等管理措施已降低了土壤盐度,这与仲启铖等[36]研究结果一致.PW样地的RH与电导率呈显著负相关,这与戚志伟等[37]研究结果一致.这可能是因为PW样地的含盐率过高,对芦苇根系产生了毒害作用,从而抑制了其根系呼吸.
3.3 研究结果对滨海围垦区土地低碳利用和管理的启示
随着人口不断增加和土地利用方式的转变,全球沿海生态系统普遍发生了大规模退化和破碎化,进而在不同程度上,丧失了包括碳固存在内的生态系统服务功能[38].我国东部沿海地区许多地方政府,长期以来,大力实施沿海围垦地造林计划,以期提高其碳汇效率[39].本实验地也实施了造林计划,但由前文分析可知,人工林样地和农田样地的RH显著高于残存湿地样地.可见,原生湿地被围垦并转变成人工林和农田后,因RH提高而导致土壤碳流失更为严重,因此,对于原生湿地的围垦需要慎重行事.He等[40]研究发现,农林复合围垦降低了长江口滨海湿地的CO2收支.虽然崇明东滩的残存湿地土壤呼吸高于人工林,但因残存湿地的初级生产力高于人工林,将各用地类型的初级生产力减去土壤异养呼吸,最终结果表现出残存湿地的净生态系统生产力高于人工林.因此,崇明东滩原生湿地围垦后建植人工林,并没有达到预期的增加碳汇的目标.Paul等[41]和Gao等[42]研究结果表明,人工林在短期内可能不会表现出明显的碳汇效果,要使人工林最终实现增加碳汇的目标,需要合理的可持续管理,并选择适宜的树种.
滨海湿地围垦区土地利用管理应该因地制宜,合理制定土地利用政策和管理措施,合理规划土地类型分布空间格局.在满足当地基本的粮食生产和经济发展需求的前提下,应尽可能通过改善水文状况,来恢复当地的优势种,以加快恢复一部分天然湿地.戚志伟等[43]发现,通过人工水位调控,适当提高地下水位,可以抑制白茅的生长,从而有助于恢复以芦苇为单优种的东滩湿地群落结构和功能.建植人工林时,应尽可能选择耐盐、生长快且具有固氮能力的树种[41],同时,适当延长轮伐期,以充分利用森林的固碳潜力,但不超过林木生长量顶点[44].对于农林复合林,应采取多层次、资源循环利用的经营模式.对于农田,应采取以免耕或少耕为基础的耕作方式[45],尽可能采取生物技术和物理方法防治病虫害、减少化肥和杀虫剂的使用,同时,将农业生产和生活的废弃物归还农田,以实现农业生态系统的物质和能量的闭合循环[46].
4结论
在崇明东滩围垦区5种不同土地利用类型中,幼龄林、中龄林和农田样地的土壤呼吸显著低于芦苇湿地和白茅湿地样地,年平均土壤呼吸速率分别为(6.48±0.82)、(6.53±0.86)、(5.08±0.6)、(8.03±1.15)和(7.48±1.00)μmol·m–2·s–1;幼龄林、中龄林和农田样地的异养呼吸分别占土壤呼吸的72.1%、79.3%和70.8%,而芦苇湿地和白茅湿地样地的异养呼吸则分别占土壤呼吸的24.6%和32.3%.与残存湿地相比,农林利用显著降低了土壤呼吸,但是在这些土地利用模式下,土壤异养呼吸和土壤呼吸的温度敏感性显著增加,可能导致土壤碳的持续排放.另一方面,土壤呼吸速率及其组分与土壤温度呈显著正相关.因此,在全球气候变暖的背景下,围垦区的土壤碳排放可能会增加.为减少土壤碳净损失,围垦区可通过调控水位等管理手段来修复和恢复更多以芦苇为优势种的湿地.
