
宁夏贺兰山国家级自然保护区岩羊(Pseudois nayaur)种群数量及结构
2022-06-28 03:12谢建冲孟德怀李宗智张致荣刘振生滕丽微
生态学报 2022年10期
谢建冲,孟德怀,李宗智,张致荣,刘振生,2,滕丽微,2,*
1 东北林业大学野生动物与自然保护地学院, 哈尔滨 150040 2 国家林业和草原局野生动物保护学重点实验室, 哈尔滨 150040
岩羊(Pseudoisnayaur)是青藏高原分布最广的有蹄类动物之一,被世界自然保护联盟(International Union for Conservation of Nature)收录为低危种(Least Concern)[1],在我国被列入国家Ⅱ级重点保护野生动物名录[2]。国内主要分布于西藏、云南、四川、内蒙古、新疆、甘肃、青海、宁夏和陕西等省区[3],其中宁夏贺兰山国家级自然保护区是岩羊的一个重要集中分布区域[4]。自20世纪80年代保护区建立以来,保护和管理力度逐年加强,采取了禁止和限制家畜放牧等一系列保护措施[5],使贺兰山植被得以恢复,环境承载力得到提高。刘振生等对贺兰山地区岩羊的种群数量及分布调查显示,岩羊种群数量约为12 178(6592—22499)只,其中贺兰山东坡是岩羊的主要分布区[6]。张明春等利用Distance 6.0软件对宁夏贺兰山国家级保护区岩羊进行数量估计,数量约为11767(9696—14282)只[7]。目前世界上野生岩羊的数量为47000—414000只,种群数量变化趋势未知[8]。
距最近的一次宁夏贺兰山岩羊数量调查已近十年,在贺兰山岩羊数量趋于稳定并存在少量波动的情况下,保持对岩羊种群数量的调查和分析是持续更新岩羊种群动态的重要组成部分,有利于对保护区整体群落及无机环境进行评估,针对特定物种制定或修改保护策略。此外,对岩羊的种群数量调查,也是对其潜在捕食者的猎物量评估,有利于对雪豹(Pantherauncia)、狼(Canislupus)、欧亚猞猁(Felislynx)等濒危捕食者的保护。本文利用样线法,先后于2020—2021年度的夏季和冬季,对保护区内的岩羊进行数量调查,采用R 4.0.1中的Distance 1.0.2进行数据分析,估测种群数量和种群密度,并对其种群结构进行探讨[6—7,9]。

图1 宁夏贺兰山国家级自然保护区Fig.1 Ningxia Helan Mountain National Nature Reserve
1 研究地概况
研究地位于宁夏贺兰山国家级自然保护区(38°19′—39°22′ N,105°41′—106°49′ E)内(图1),属贺兰山东坡[10—11],保护区面积为193535.68 hm2[12]。贺兰山处于荒漠与半荒漠草原之间,具有典型的温带大陆性气候特征,年平均气温-0.8℃ (2900 m处),年平均降水量在200—400 mm,主要集中于7—9月。山体大致呈南北走向,南北长约250 km,东西宽20—40 km,山地海拔2000—3000 m,主峰敖包疙瘩海拔3556 m[10—11,13]。贺兰山植被类型丰富,乔木包含云杉、油松、杜松、灰榆、山杨等,本次研究主要在海拔1100—2500 m左右的山地疏林草原带和山地针叶林带中进行[10—11]。贺兰山地区野生动物共计54科218种,其中国家Ⅰ级保护动物8种,Ⅱ级保护动物32种[10,12]。
2 研究方法
2.1 距离取样法(Distance sampling)
传统的随机抽样法统计,随机选取样区,假设样区内统计对象的密度对整个调查区域具有代表性,关键还在于假设样区内所有统计对象被计入。距离取样法放宽了这一关键假设,不必要统计样区内所有统计对象,相对于样区抽样法适用于高密度的静态物种,距离取样法更适用于低密度的流动物种。样线调查中,假设统计对象是均匀分布的,距离样线越远的目标发现概率越低,使用每个统计对象发现点相对样线的距离建立发现概率函数模型,可以推算多少统计对象被遗漏,从而估计整个调查区域的密度,消除较远距离的观察值可以减少对发现概率函数模型的过度影响[14]。发现概率主要受到统计对象距离样线距离、天气状况、地表植被、统计对象社群大小等因素影响[15]。
2.2 样线调查
在宁夏贺兰山国家级自然保护区内,依据地形梳齿形走势,顺从沟道东西走向设置样线。样线在保护区内南北均匀排布,尽量包括其中的主要沟道,样线起始点尽量接近沟道的东西边缘,以求调查地区覆盖更广,对保护区整体区域的调查更为详尽。岩羊的活动具有家域性[16],且岩羊日活动方式为早晨在低海拔地区饮水,随着觅食过程逐渐向高海拔移动[17],因此岩羊的日活动范围应当呈现带状,样线设置彼此之间最短距离相隔2 km以上[7],可以避免过多的个体被相邻样线重复记录。岩羊活动节律呈现双峰型,主要活动时间集中在早晨和黄昏[18—19],故样线观察设定时间为每日8:00—12:00和15:00—17:00,以求更全面的记录个体。观察者沿样线自沟口向上步行,速度2 km/h上下,使用8×32 Kowa双筒望远镜辅助观察,使用BUSHNELL YARDAGEPRO 1000 型激光测距仪测量岩羊群与观察者直线距离,使用65式军用罗盘仪测定岩羊群相对于观察者位置,与样线前进方向所成夹角,使用集思宝G120记录航迹。由于岩羊具有社群性,调查需记录每群岩羊的个体组成[19]。
2.3 数据分析
种群密度和种群数量分析由R 4.0.1完成。将野外调查所获得的数据,通过Distance 1.0.2进行数据分析,将两次调查的数据分别分析,得到每个季节岩羊的种群密度和95%的置信区间[6—7,20—21]。种群密度的计算公式如下:
式中,D:种群密度(只/km2);n:岩羊社群数量;f(0):垂直距离为零的概率密度函数;E(s):群大小,s为岩羊社群序列号;L:样线总长度。
获取探测函数g(x),即垂直距离为x处动物被发现的概率,若0距离处的所有对象都能被检测到,即g(0)=1,根据探测函数,可以估计f(0),公式如下:
式中,w:样线单侧宽度,x:样线垂直距离。
探测函数g(x),包括半正态分布(Half-normal)、均匀分布(Uniform)和风险率(Hazard-rate)3种统计分布。利用级数展开,包括余弦(Cosine)、赫米特多项式(Hermite polynomial)和简易多项式(Simle polynomial),调整上述3种统计分布。模型和调整项的最佳拟合程度由赤池信息准则(the second order Akaike index criterion, AIC)确定。探测函数右侧5%的数据右截断(Right-truncation),距离样线最远的观测值不继续分析,减少误差。
R语言具体操作:首先加载Distance包,读取整理后的数据集文件,包括每条样线编号、研究地区名、研究地区总面积(hm2)、每条样线长(km)、每个岩羊社群发现点位距样线距离(m)、每个岩羊社群大小等。将截断距离设置为1 km,得到默认模型拟合数据;用ds函数中的“key”来指定探测函数的不同形式;用“adjustment”来调整探测函数的的拟合程度;将赤池信息准则的最小者作为最优模型;用summary函数提取最优模型的详细信息。

3 结果
本研究先后于夏季和冬季对岩羊种群数量进行2次重复调查,夏季调查时间为2020年8月,冬季调查时间为2020年12月至1月,选取的19条样线,两次重复调查共计281.108 km,样线最长13.477 km,最短2.991 km,平均每条样线长7.398 km。夏季观察到岩羊92群576只,遇见率为4.098只/km,岩羊群距样线平均垂直距离为72.961 m;冬季观察到岩羊94群382只,遇见率为2.718只/km,岩羊群距样线平均垂直距离为53.843 m。
3.1 岩羊种群数量
探测函数和调整项的组合中,风险率、风险率+余弦、风险率+赫米特多项式和风险率+简易多项式,均为拟合观测距离和发现岩羊概率效果的最优模型(表1),用summary函数提取出这四个模型组合的详细信息,其中风险率+余弦、风险率+赫米特多项式和风险率+简易多项式3种组合的分析结果相同,而风险率的分析结果不同且较不合理,因此认为风险率+余弦、风险率+赫米特多项式和风险率+简易多项式三种组合的分析结果为最优结果,选择其中一种组合分析结果作展示(表2)。其中均匀分布函数必须添加调整项使用,所以运算产生报错。分析得出夏季岩羊种群数量为5 176(2554—10490)只,种群密度为2.674(1.320—5.420)只/km2;冬季岩羊种群数量为15752(7294—34017)只,种群密度为8.139(3.769—17.58)只/km2(表2)。

表1 不同组合模型的赤池信息量准则

表2 R中Distance估计的宁夏贺兰山岩羊种群密度和数量
把最优模型的探测函数进行5%的右截断(right-truncation),消除距离较远的观测值,有效样线观测值为400 m左右,从垂直距离和探测函数的观测图(图2)中可以发现,岩羊在0—100 m这个区间范围内被观测到的概率较大,说明可被观测到的大部分岩羊实体距样线较近,夏季岩羊被观测到的概率更高,冬季岩羊被观测到的高概率区域距离样线更近。
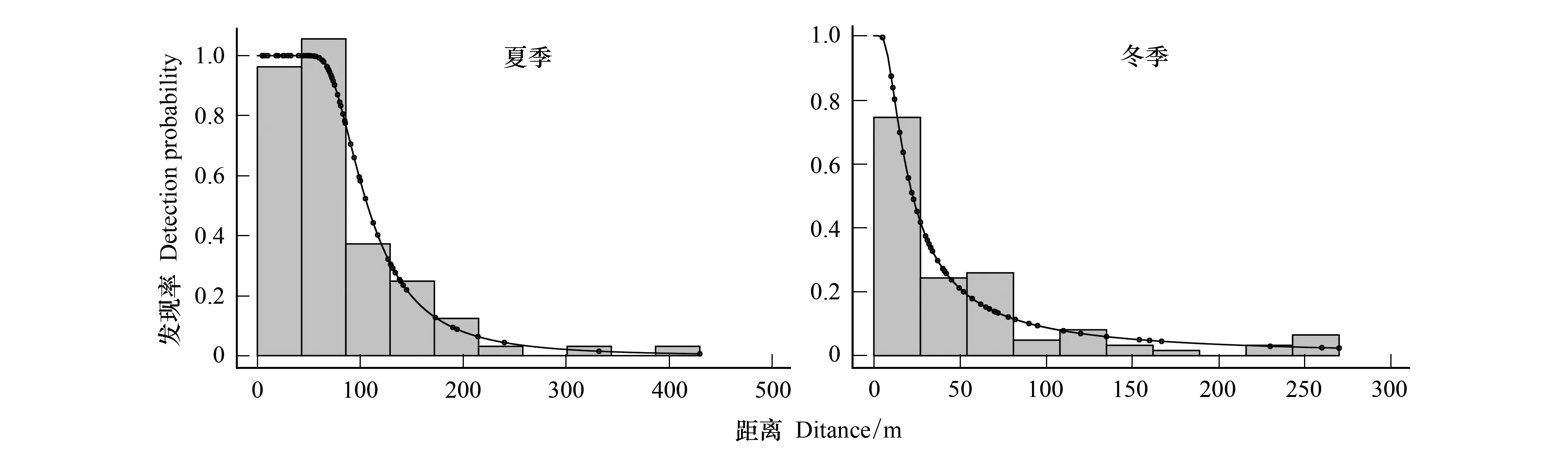
图2 岩羊的垂直距离和探测函数的直方图Fig.2 The histogram of distance and detection function of Blue Sheep
3.2 岩羊种群结构
依据岩羊群社群组成结构划分后的结构显示:夏季调查中出现频次最高的是雌幼群(52.17%),其次为混合群(14.13%);冬季调查中出现频次最高的是混合群(26.60%),其次为雌幼群(20.21%);冬季相较于夏季,混合群和雌雄群数量上升了近一倍,雌幼群下降明显(60.42%),另外独雌和独雄也有所增加(表3)。此外,两次调查都发现了脱离群体的亚成体,冬季还发现了一次迷失的幼体。

表3 夏季和冬季观测到的不同类型岩羊群数量
夏季调查中,岩羊的平均群大小为(6.261±8.023)只,最大群为47只,最小为1只,混合群的平均群大小最大;冬季调查中,岩羊的平均群大小为(4.064±4.229)只,最大群为25只,最小为1只,混合群的平均群大小最大。冬季相比于夏季,除了雄性群大小上升外,其它类型群大小均有所下降(表4)。Kruskal-WallisH检验表明,夏季和冬季的群大小差异显著(χ2=6.592,df=1,P=0.010),混合群的群大小在不同季节的差异不显著(χ2=0.961,df=1,P=0.327),雌幼群的群大小在不同季节的差异显著(χ2=4.356,df=1,P=0.037)。将不同季节的岩羊群按照群大小分成1—5、6—10、11—15和 >15四个区间组,可以看到2020—2021年度的夏季和冬季,岩羊集群大小主要集中在1—5之间(72.04%),卡方检验表明,岩羊群大小不同区间组在不同季节组的差异不显著(χ2=3.574,df=3,P=0.311)(表5)。
将可辨别的个体依据性别和年龄划分为雄性、雌性、幼体/亚成体。夏季调查中,雄性岩羊占比11.27%,雌性岩羊占比56.74%,幼体/亚成体占比31.99%;冬季调查中,雄性岩羊占比29.61%,雌性岩羊占比50.15%,幼体/亚成体占比20.24%。无论是夏季还是冬季,雌性都占有很高的比例,冬季相比夏季,雄性有所增加,幼体和亚成体有所下降。卡方检验表明,夏季和冬季岩羊组成成分之间差异极显著(χ2=47.574,df=2,P<0.001)(表6)。

表4 夏季和冬季不同类型集群岩羊的平均群大小

表5 夏季和冬季不同集群大小的岩羊群数量

表6 岩羊不同组成成分观测数据及比率
4 讨论
4.1 岩羊种群数量
与刘振生等[6]和张明春等[7]使用Distance统计的结果相比,本研究的结果中,夏季的岩羊数量低于前两者,冬季的岩羊数量高于前两者。岩羊出生时间几乎固定[25—27],而死亡时间全年不固定,保护区外围封闭,内部与内蒙古国家级自然保护区相连,并且岩羊存在家域性[16],因此岩羊迁入和迁出率极低,岩羊的种群数量主要由出生和死亡调控。鉴于岩羊的繁殖行为,岩羊的新增出生个体应主要集中在每年的5月和6月,而后逐渐死亡衰减,直至来年新增个体陆续出生补充[25—27]。因此理论上,实际岩羊数量,夏季应当是多于冬季的[9]。本研究的结果与这一理论相悖,对岩羊群距样线平均垂直距离统计显示,虽然冬季发现岩羊数量更少,但发现距离更近,相比夏季几乎全部发现于山坡上,冬季有更多的个体被发现于沟底,发现距离的远近直接影响了对岩羊数量的统计计算。一方面,冬季食物匮乏,岩羊需要下降到低海拔区域觅食和饮水;另一方面,冬季正是岩羊的繁殖期[23,28],冬季在沟底发现的雄性个体要多于夏季,雄性需要在平坦的地形中争斗。夏季岩羊的栖息地选择会更多考虑荫蔽的环境,只有在比较窄的沟道中,岩羊才会出现在沟底,在开阔沟道中,岩羊偏好活动于阴坡上。此外,岩羊冬夏栖息地的选择差异还可能受其它因子所影响[29—31]。山地地形的阻隔会使样线调查的数据收集相对平地更困难,偶尔会听到幼体的叫声却会因为地形阻挡,人目光无法直接观测到个体只能被迫忽略不计入,这使得无论夏季还是冬季,岩羊数量的统计结果都会少许偏低。贺兰山家畜等主要同域竞争者迁出后[32],岩羊的可利用生境范围增大,增加了种群扩散的机会和条件,因此对于冬季调查的岩羊数量,本研究认为是确实的,并且可能偏低。
刘振生等的数据收集时间为2003年11—12月,与本研究的冬季调查时间相近,岩羊密度为5.144只/km2,相比之下,17年间增长了2.995只/km2, 平均年增长率为3.42%。张明春等的数据收集时间为2010年3—12月,冬季岩羊密度为6.48只/km2,相比之下,10年间增长了1.659只/km2, 平均年增长率为2.56%。张明春等的结果相对于刘振生等的结果,冬季岩羊密度平均年增长率为3.71%,这说明岩羊密度增长速率在下降。在没有较大环境和资源变化,以及疫病爆发的情况下,种群密度的增长速率下降,表明岩羊种群数量可能已经超过环境容纳量的一半。保护区的合理管理保护对岩羊种群数量的持续增长提供了良好的空间环境,但是在应对重大疫病时,缺乏捕食者的调控和合理的基因交流,将会成为贺兰山岩羊种群的弱点。
由于距离取样法是基于样线调查法收集数据,所以更高的抽样强度意味着获得的数据将更全面。距离取样法假设统计对象是均匀分布的,而实际上宁夏贺兰山岩羊分布是不均的[6],本研究的调查中也发现岩羊多分布于马莲口和苏峪口这两个沟道。因此样线总长度越长,对贺兰山岩羊密度的估算越准确,本研究的总样线长度少于刘振生等[6]的496.2 km,这使得本研究的岩羊数量调查结果误差,较刘振生等可能偏大。样线调查的时间是不相同的,空间上也不完全相同,因此本研究和刘振生等的样线调查中对岩羊发现概率的影响因素,是不完全相同的。本研究选择晴天调查,以求减少天气状况这一主要影响因素;地形地貌和地表植被等天然遮蔽伪装等因素是所有调查者都难以规避的影响因素;岩羊距样线距离和岩羊社群大小是与岩羊自身行为相关的,实验不可控的随机因素,只能通过更高的抽样强度降低偶然性。
4.2 岩羊种群结构
两次调查中,岩羊的主要集群类型为混合群或雌幼群,夏季的雌幼群比混合群多了近三倍,此时是岩羊生产时间刚过,相比冬季,幼体更多,所以雌幼群可能是夏季岩羊的主要集群类型,而冬季的雌幼群数量会有所下降[26—27]。冬季的混合群、雄性群、雌雄群都要多于夏季,这可能与发情期的争斗行为有关,冬季也的确看到雄性聚群争斗的现象[26—27,33]。冬季相较于夏季,除雄性群外,各类型平均集群大小均有所下降,雄性聚群增大应当也与发情期争斗有关。混合群聚群夏季较大的原因在于,观测到几群较大规模的混合群,可能是多群岩羊共同利用同一片草场资源较好的冲积滩地,而组成的大规模集群。夏季部分亚成体即将体成熟,亚成体雌性的体型和角大小已接近成年雌性个体,雌性个体得到补充,这可能是夏季雌性群、雌雄群和雌幼群平均群大小相较于冬季大的原因,与夏季比冬季雌性记录到更多这一现象相符合。夏季平均群大小略高于冬季平均群大小,这与之前的研究相似[19,24,34—35]。且无论夏季还是冬季,集群大小都主要集中在1—5这个区间,说明集小群仍然是贺兰山岩羊的主要特征[24,36]。
两次调查结果中,成年雌性数量占比都占半数以上,与之前的研究相近[7,35],说明成年雌性个体是贺兰山岩羊的主要组成部分。冬季的求偶争斗行为,让雄性在冬季比夏季更易被观察到。夏季幼体/亚成体的数量比冬季幼体/亚成体的数量多一倍有余,一方面是部分亚成体体成熟转变为成体,另一方面新生的幼体较成体脆弱,成年之前会部分衰减。无论是夏季还是冬季,幼体/亚成体在群体中的占比都比之前研究略高,原因可能是本研究将亚成体的数量一并计入,而其只计入幼体数量[7]。由于生殖潜力的不等,岩羊的雄性一般都要少于雌性,但本研究观测到的雄性个体占比是要少于之前的研究的[7,35],特别是夏季雌雄比已经超过了5,对比之前研究的雌幼比来看,夏季的雌幼比与之前的研究相近[7,35],说明观测到的雌性个体数量较为可信,而雄性可能偏低。夏季雄性个体不必面临像雌性个体一样哺育幼体的生殖压力,因此可能不必要去往高风险的沟道中觅食,而且相比雌性个体更为强壮的体型,雄性个体无须畏惧空中天敌的威胁,敢于向山顶草场觅食,雌性个体则必须带领易被捕食的幼体在狭窄且险峻的沟道两侧躲避来自地面和空中的天敌。
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
生物学通报(2021年5期)2021-03-16
海洋通报(2020年3期)2020-10-20
——以北长山岛为例
海洋开发与管理(2020年9期)2020-10-13
水产养殖(2020年9期)2020-09-24
自然保护地(2020年2期)2020-06-25
现代农业科技(2020年10期)2020-06-04
中外文摘(2019年10期)2019-11-12
阅读与作文(小学低年级版)(2019年6期)2019-07-26
中外管理(2019年5期)2019-05-22
