
长期增温和氮素添加对荒漠草原不同植物功能群特征的影响
2022-07-05 08:47王冰莹韩国栋武倩朱毅鞠馨
草原与草坪 2022年2期
王冰莹,韩国栋,武倩,朱毅,鞠馨
(内蒙古农业大学草原与资源环境学院草地资源教育部重点实验室,农业部饲草栽培、加工与高效利用重点实验室,内蒙古自治区草地管理与利用重点实验室,内蒙古 呼和浩特 010020)
气候变暖和大气氮沉降增加是全球变化的重要驱动因子。气候变暖与氮沉降增加对生态系统产生的影响以及生态系统对气温上升和氮素沉降的反馈是国际生态学领域的研究热点[1]。温度的变化会对植物的物质积累、生长、发育产生直接影响[2]。由于人类活动的影响,大气氮沉降的速率也在增加[3]。不同植物的生长随着可利用氮含量的变化而改变,进而使植物群落结构发生变化[4]。氮素输入的持续增加会改变植物群落的物种丰富度和群落的动态特征[5]。在研究增温和氮素添加对植物多样性的影响时,可以缩小研究尺度,将局部细节从各种角度分析描述。功能群是指对特定环境因素有相似反应的一类物种,植物功能群多样性是维持群落结构复杂性和稳定性的基础,一定程度上反映了草地生态系统的健康状况[6]。因此,探究长期增温和氮素添加对草地植物功能群特征的影响对于维持草地生态系统稳定具有重要意义。
荒漠草原是草原向荒漠过渡的旱生性草原生态系统,目前为止,对荒漠草原不同功能群的研究还相对薄弱[7]。武倩[8],王晨晨等[4]对短花针茅荒漠草原的研究表明,长期增温与氮素添加使得荒漠草原群落结构发生变化,植物功能群的引入对研究荒漠草原物种多样性的生态系统功能和维持机制注入了新的活力[9],虽然目前对功能群的研究还不是很完善,但仍有一些研究认为功能群的多样性比物种多样性更加深刻地影响生态系统的整个过程[10]。因此,本试验以内蒙古四子王旗短花针茅荒漠草原为研究对象,利用长期增温和氮素添加试验平台,探讨短花针茅荒漠草原不同植物功能群对模拟增温和氮素添加的响应,旨在揭示全球气候变暖和氮素添加对荒漠草原植物功能群的影响,为干旱区荒漠草原科学合理利用提供一定的理论依据。
1 材料和方法
1.1 试验区概况
试验区位于内蒙古农牧业科学院综合试验示范中心四子王基地(N 41°46′43.6″,E 111°53′41.7″,海拔1 456 m)。四子王旗地处大青山以北的内蒙古高原中部,属于典型的中温带大陆性季风气候。春季干旱多风,夏季炎热,冬季寒冷。四子王旗(1960-2018年)多年平均气温3.7 ℃,月平均最高温度集中在6-8月,≥10 ℃的年积温为2 200~2 500 ℃,无霜期90~120 d。年均降水量221.7 mm,降水主要集中在6-9月,年平均日照时数为3 117.7 h。试验地区的土壤类型大多为淡栗钙土,土壤较瘠薄,钾含量丰富,氮和磷较少,有机质含量较低。植被层低矮,种类组成较贫乏。主要是以短花针茅(Stipabreviflora)为建群种的荒漠草原,其他优势物种主要有冷蒿(Artemisiafrigida)、无芒隐子草(Cleistogenessongorica)、银灰旋花(Convolvulusammannii)、木地肤(Kochiaprostrata)、细叶葱(Alliumtenuissimum)等。
1.2 试验设计
该试验采用2×2因素随机裂区设计,共6个区组,其中包括增温和不增温2种主处理样方,共12个主样方。每个主处理样方又被分裂为面积相等的2个有氮素添加和无氮素添加副处理样方。试验共4种处理:无增温和氮素添加(CK)、增温(W)、氮素添加(N)和增温+氮素添加(WN),每个处理6次重复,在每个处理样方中央各设置一个1 m×1 m的永久监测样方。共设计了24个处理样方,面积共计144 m2。从2006年3月开始,6个增温区内各安装一个红外线辐射器进行加热,距地面2.25 m,功率设置为2000 W,全年不间断。同时在每年5月末6月初雨季来临之前选用硝酸铵(NH4NO3)进行施肥处理,施肥量为纯氮10 g/m2。
1.3 测定内容和方法
1.3.1 植物群落物种组成及功能群划分 2019年8月中旬取样,在永久监测样方内记录物种数目及名称,并记录每种植物的高度、盖度、密度。具体方法是把活动样方框(1 m×1 m)各边平均划分成10 cm等间距的小孔,然后用线绳相连,活动样方框就被平均划分成100个小格。密度是每种植物在100个小格中株丛总数;盖度是每种植物垂直投影面积占1 m×1 m的百分比。按照物种生活型差异,将群落中出现的物种划分为4个功能群:多年生禾草(Perennial grasses,PG),多年生杂类草(Perennial forbs,PF),灌木和半灌木(Shrubs and semi-shrubs,SS),一、二年生植物(Annuals and biennials,AB)。样地内出现的物种、划分出的功能群以及结果中使用的功能群见表1。
1.3.2 地上生物量 采用直接法估测地上生物量,在植物生长旺盛期(8月),每个小区选取50 cm×50 cm样方,然后分物种齐地面刈割并收集枯落物,把样品置于65 ℃恒温下烘干,48 h后烘至恒重,称其干重(即物种生物量)。
1.4 测度方法
本研究采用3个参数,即Margalef丰富度指数(A)、Shannon-Wiener多样性指数(H),Pielou均匀度指数(E),对植物群落物种功能群进行评价。3个参数的计算公式如下:
Margalef指数A=(S-1)/LnN
Shannon-Wiener指数H==∑PiLn(Pi)
Pielou指数E=-∑PiLn(Pi)/Ln(S)=H/Ln(S)
式中:S为群落中物种数目;N为所有物种个体总数;Pi为重要值。
重要值Pi=(RC+RH+RD)/3
式中:RC为相对盖度;RH为相对高度;RD为相对密度。RC、RH和RD的计算方法详见文献[11]。
1.5 数据分析
在SAS 9.2统计分析软件中采用双因素方差分析,分析增温、氮素添加及它们的交互作用对上述各指标的影响。最后再对各指标进行单因素方差分析,研究对照、氮素添加、增温、增温+氮素添加4个处理间的差异,所有图均使用Sigmaplot 14.0进行绘制。
2 结果与分析
2.1 增温和氮素添加对不同功能群植物高度、盖度、密度的影响
增温对多年生杂类草的盖度有显著影响(P<0.0001),对其他3个植物功能群的高度、密度、盖度没有显著影响(P>0.05)(表2)。氮素添加及其与增温的交互作用对不同植物功能群的高度、盖度、密度没有显著影响(表2)。与对照相比,增温处理下多年生杂类草的盖度增加了66.3%(P<0.05)(图1);氮素添加处理对多年生杂类草的盖度影响不显著;增温+氮素添加处理下多年生杂类草的盖度增加了62.8%。增温处理下灌木和半灌木的高度减少了70%;氮素添加、增温+氮素添加处理对灌木和半灌木的盖度、密度影响不显著。增温、氮素添加、增温+氮素添加处理对多年生禾草的高度、密度、盖度影响不显著。氮素添加处理下一、二年生植物的盖度和密度分别减少了53%和71%。
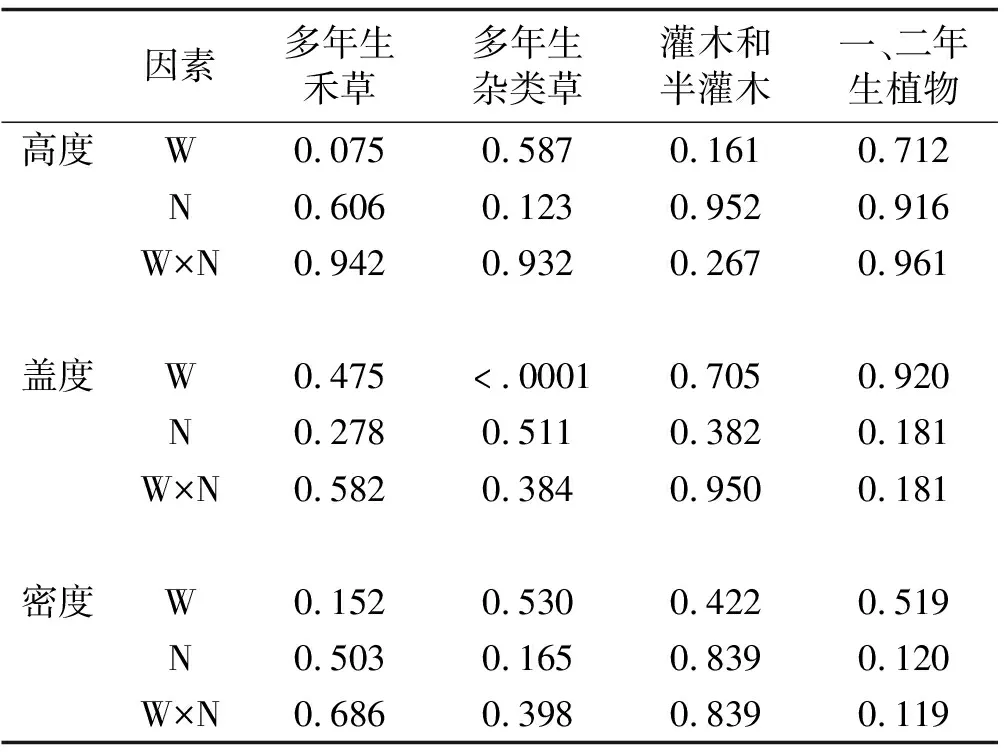
表2 增温、氮素添加及其交互作用对不同功能群植物群落高度、盖度、密度影响的方差分析结果
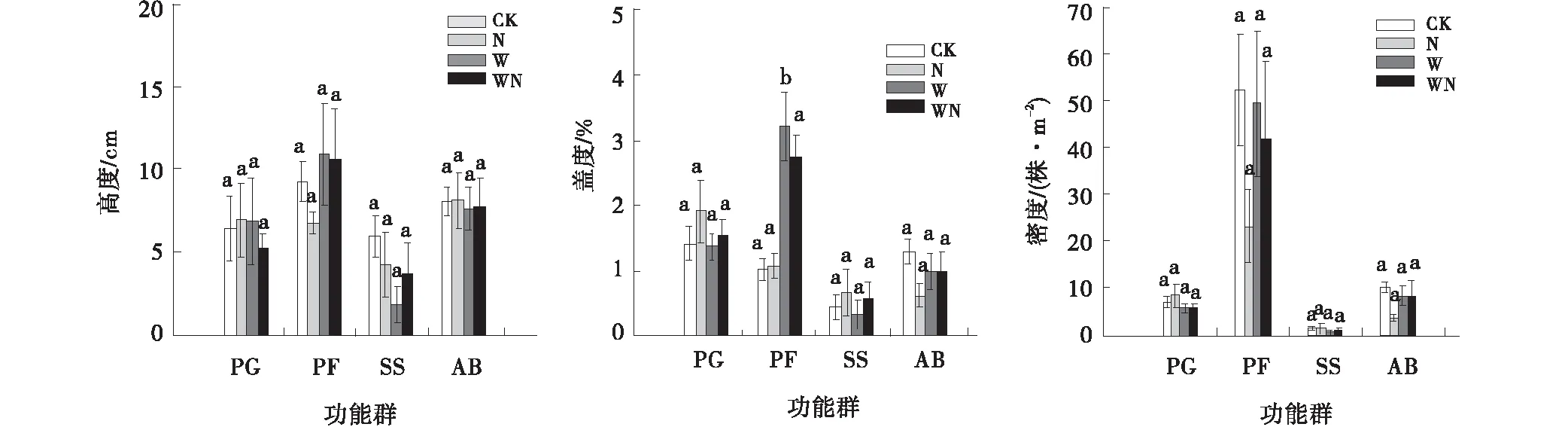
图1 不同植物功能群的高度、盖度、密度
2.2 增温和氮素添加对不同功能群植物地上生物量的影响
氮素添加处理对多年生杂类草的地上生物量有显著影响(P<0.001),对其他3个植物功能群的地上生物量没有显著影响(P>0.05),增温及其与氮素添加的交互作用对4个植物功能群地上生物量没有显著影响(表3)。与对照相比,增温处理下多年生禾草地上生物量减少了47.7%;氮素添加处理下灌木和半灌木地上生物量增加了51.7%,一、二年生植物地上生物量减少了47.4%,多年生杂类草的地上生物量减少了46.1%(P<0.001);增温+氮素添加处理下多年生禾草地上生物量减少了43.7%(图2)。
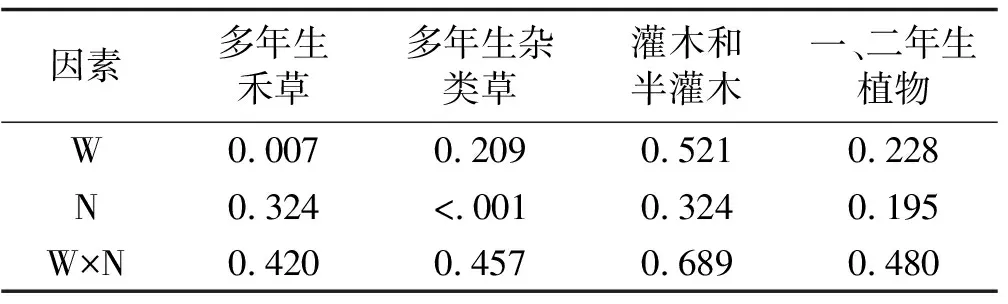
表3 增温、氮素添加及其交互作用对不同功能群植物群落地上生物量影响的方差分析结果
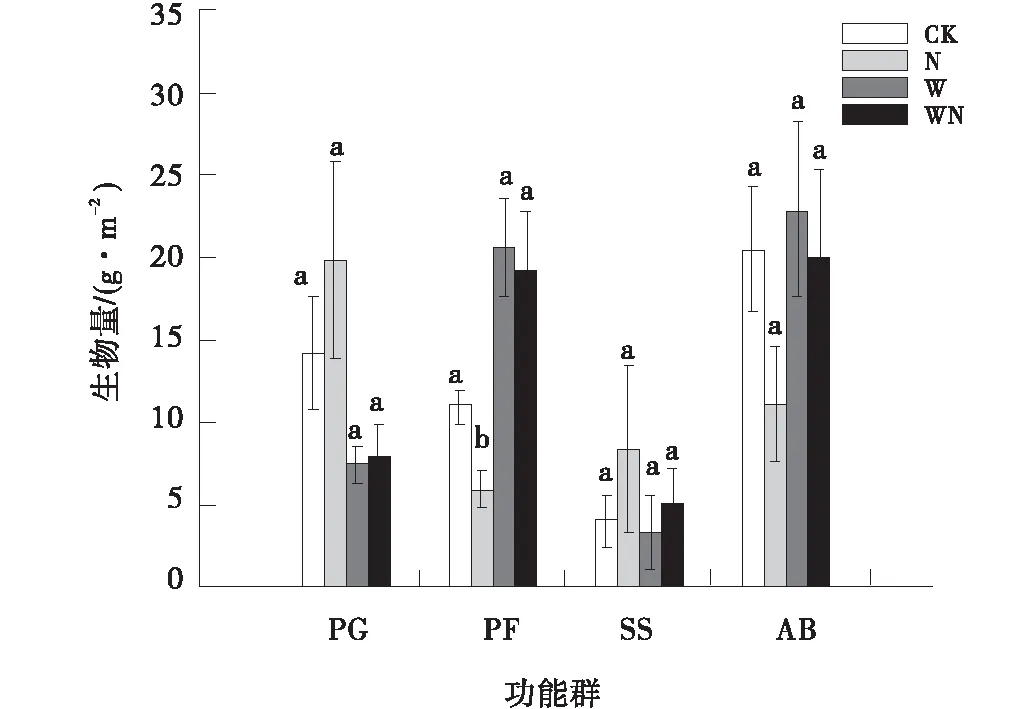
图2 植物功能群的地上生物量
2.4 増温和氮素添加对植物功能群多样性的影响
氮素添加对多年生禾草和多年生杂类草的Margalef丰富度指数有显著影响(P<0.01),对Shannon-Wiener指数和Pielou均匀度指数没有显著影响。增温对4种功能群的多样性指数都没有显著影响(表5)。功能群Margalef丰富度指数在氮素添加处理和增温+氮素添加处理下显著增加(P<0.05),在增温+氮素添加处理下Margalef丰富度指数最大,为1.74,显著大于对照处理。而Shannon-Wiener指数和Pielou均匀度指数在氮素添加处理和增温+氮素添加处理下有下降的趋势,且均不显著。Margalef丰富度指数、Shannon-Wiener指数和Pielou均匀度指数在增温处理下变化不大(图4)。
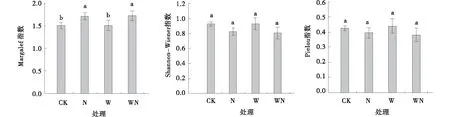
图3 植物功能群的多样性
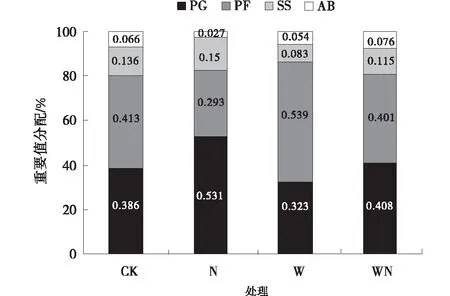
图4 不同功能群的植物重要值
2.5 増温和氮素添加处理下不同功能群植物的重要值
增温处理多年生杂类草重要值最高,达到53.9%,对照处理,增温+氮素添加处理重要值变化不明显。一、二年生植物重要值在各处理间变化不明显,在增温+氮素添加处理下重要值最高,达到7.5%。灌木和半灌木以及多年生禾草的重要值均在氮素添加处理下最高,分别达到15.0%、53.1%(图5)。
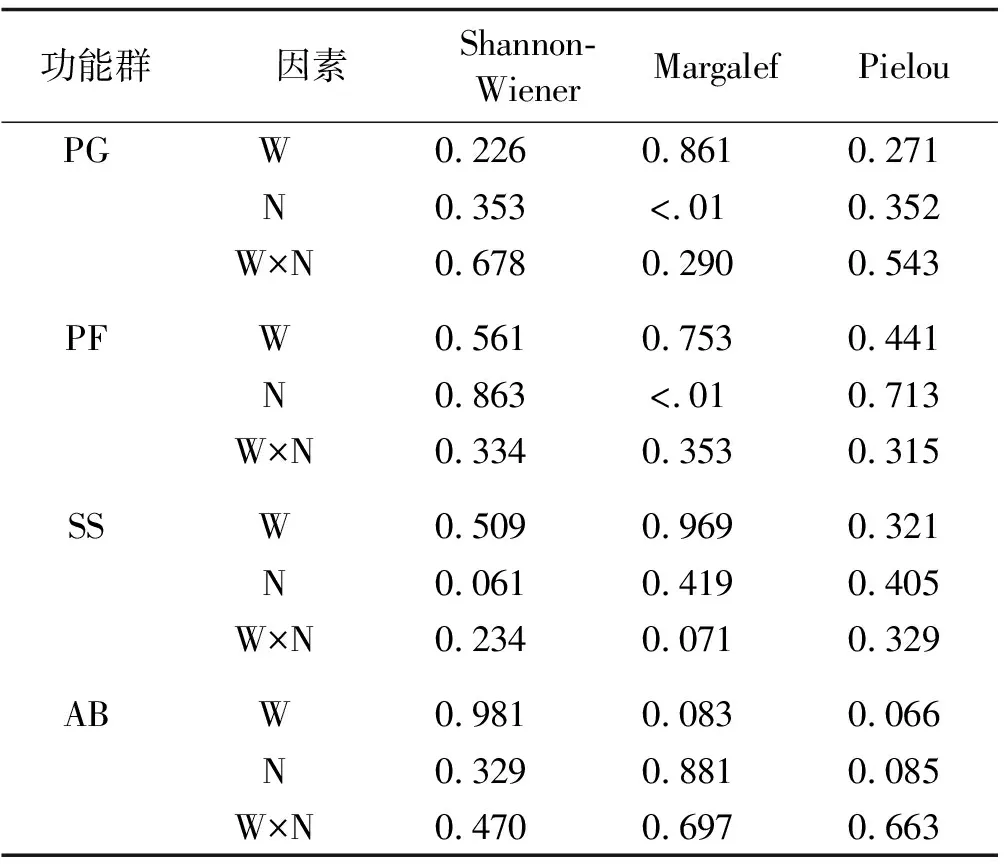
表4 增温、氮素添加及其交互作用对不同功能群植物群落α多样性的方差分析结果
3 讨论
在全球气候变暖的大背景下,研究生态系统与气候变暖作用机理的方法层出不穷,例如电阻加热、远红外线照射、互交移植等多种模拟气候变暖的方法[12]。在本研究中,采用野外自然条件下红外线增温的试验设计,通过试验发现增温导致灌木和半灌木的高度有所下降,但是其他3种功能群植物的高度呈现增加的趋势。这与Muhamed等[13]的研究一致。Harte等认为,增温满足了植物对热量的需求,且对植物生长有利,使植物种群高度在整体上有所增加[14-15],但是本研究中不同功能群植物群落高度对增温的响应不同,这可能是因为研究草地的类型及年限不同,导致研究结果出现差异。增温处理导致多年生杂类草的盖度显著增加,而其他3种功能群盖度呈现降低趋势,氮素添加对不同功能群植物的盖度没有显著影响,但呈现增加的趋势,增温+氮素添加处理对不同功能群植物的盖度也无显著影响,这与李元恒[11]的研究一致。本研究发现,氮素添加导致多年生禾草的密度增加,其他3种功能群植物的密度减少。这与Gough等[16]试验认为随着氮沉降的增加,物种密度逐渐减小的结论相一致。温度的升高和降低能够影响植物的蒸腾作用,改变土壤中水分的含量,使植物对水分的吸收受到影响,其生长和生物量也不同程度受到影响。而植物自身通过改变土壤中物质的含量引起的植物的改变,都将会对生物量产生一定的影响,而生物量是大多数生态系统结构和功能的最直接的综合体现,它是植物生态学特性与其生活环境之间相互作用的产物[17-19]。在本研究中,氮素添加导致多年生杂类草的地上生物量显著减少,这与Su等[20]、Xi等[21]的研究一致,而多年生禾草、灌木和半灌木的地上生物量有增加的趋势,这与潘占磊[22]研究不一致。这可能是因为在丰水年份,干旱压力缓解后,氮素成为了限制植物生长的主要因素[23],氮素添加使土壤中可利用氮含量增加,从而促进植物的生长。有研究表明增温提高[24],或抑制[25],或不改变植物地上生物量[26],这可能与生态系统的初始气候条件、优势物种的特性、生物多样性和生物地球化学有关。在本研究中,增温导致多年生禾草、灌木和半灌木的地上生物量减少,多年生杂类草和一、二年生植物的地上生物量增加,这与潘占磊[22]研究结果一致。由于一、二年生植物和多年生禾草对养分的利用策略不同,一、二年生植物可以快速利用养分,使其在资源竞争中占据有利地位,而多年生禾草对资源利用比较保守,与一、二年生植物相比,在资源竞争中处于劣势[27-28],因此,增温改变种间关系可能是导致多年生禾草地上净初级生产力降低的原因。
本研究中,增温+氮素添加处理下多年生杂类草和多年生禾草重要值极大,灌木和半灌木和一、二年生植物重要值极小,因此推测以短花针茅为主的多年生杂类草和多年生禾草存在一定的正相互作用,但具体相互作用的机制尚不清楚,仍需进一步的探究。本研究发现,氮素添加对多年生禾草和多年生杂类草的Margalef丰富度指数有显著影响,这与Zong等[29]研究认为,植物的物种丰度指数及多样性指数并未随氮沉降量的变化而产生显著改变的结论不一致,这可能与土壤原来的氮素含量水平及物种间的差异存在密切关系,同时也有大量研究表明,植物物种多样性会随着氮素的增加而减少,而对减少机理的研究却相对甚少。本研究中增温导致Shannon-Wiener多样性指数、Pielou均匀度指数有增加的趋势,但是变化趋势不大,这与武倩[8]的研究结果一致。功能群Pielou均匀度指数在增温、氮素添加、增温+氮素添加处理下均与对照处理相比变化趋势不大,可能是因为本研究增温与氮素添加试验已在短花针茅荒漠草原开展13年,处于长时间增温和氮素添加处理,不同小区都已经存在较为稳定的群落,各个小区内不同功能群植物种类基本相似,因此功能群Pielou均匀度指数差异不显著。刘思雅[30]研究表明,在植物生长的不同时期增温和施氮所起的作用有所不同。温度较低、雨水相对不足的月份,增温对优势种的生长起抑制作用,有利于群落的物种数目和多样性指数的增加,而人工施氮的效应则恰好相反,使群落的物种多样性指数降低。并不是所有植物群落特征都会随着温度的升高而有明显的变化。说明荒漠植物群落在短时间增温的条件下并不会产生明显影响。
4 结论
长期增温和氮素添加对植物群落的增温及其交互作用对不同功能群植物的高度、盖度、密度有不同程度的影响,增温显著增加多年生杂类草的盖度。氮素添加处理显著降低多年生杂类草的地上生物量,显著增加多年生禾草和多年生杂类草的Margalef丰富度指数。长期增温和氮素添加改变了短花针茅荒漠草原植物群落的组成和结构,促进了草地的可持续利用。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
作物学报(2022年10期)2022-07-21
作物学报(2022年9期)2022-07-18
安徽农学通报(2022年9期)2022-06-14
林业调查规划(2022年3期)2022-06-13
作物学报(2022年3期)2022-01-22
安徽农学通报(2020年7期)2020-05-26
诗潮(2019年10期)2019-11-19
阅读与作文(英语高中版)(2019年9期)2019-11-07
