
评“茶树起源于第三纪宽叶木兰”的观点
2022-07-08 05:43杨世雄余香琴
广西林业科学 2022年3期
杨世雄,余香琴
(中国科学院昆明植物研究所 东亚植物多样性与生物地理学重点实验室,云南昆明 650201)
“茶树起源于第三纪宽叶木兰”的观点源自“从木兰化石论茶树起源和原产地”一文(下文简称“木兰化石”),此文于1997年在《农业考古》上发表[1]。“木兰化石”基于1978年出版的《中国植物化石》(第三册:中国新生代植物)[2]描述的宽叶木兰(Magnolia latifolia)和中华木兰(M. miocenica)两个化石种(均为叶化石),认为“茶树叶片的形态、叶脉构造、侧脉对数及夹角大小、侧脉不达叶缘并向上弯曲与上方侧脉相连、叶尖形态等特征与宽叶木兰和中华木兰古植物化石有较多的相似之处(实际上上述特征多为中华木兰的特征,笔者注)。所以,推想“茶树是在第三纪特定的生态环境中由宽叶木兰演化而来”。近十多年来,“茶树起源于第三纪宽叶木兰”的观点在茶叶界颇为流行,在有关地方茶产业的论著中时有出现[3-5],在互联网上更是充斥着各种版本的引用和解读,一些地方博物馆里也摆上了标注为“宽叶木兰”的化石。
在植物分类学上,茶树(Camellia sinensis)隶属山茶科(Theaceae)山茶属(Camellia)茶组(Sect.Thea),是现代植物[6-7]。宽叶木兰和中华木兰隶属木兰科(Magnoliaceae)木兰属(Magnolia),是化石植物;前者出土于云南景谷的渐新世(Oligocene)地层;后者发现于山东临朐的中新世(Miocene)(山旺组)地层,《中国植物化石》一书中并没有该化石在云南等其他地方的分布[2]。“木兰化石”在介绍中华木兰时,其产地只有云南景谷傣族彝族自治县、临沧(现为临沧市)、沧源佤族自治县、澜沧拉祜族自治县、景东彝族自治县、梁河县和腾冲(现为腾冲市),其模式产地——山东临朐只在后面的论述“向东传至浙江嵊县、河南南阳、山东山旺。向南海南岛和东南亚也有中华木兰化石报道”中提及[1],也没给出其他地方“中华木兰化石报道”的文献。在地质年代上渐新世和中新世分别是新生代第三纪(Tertiary)的第三和第四个时期,第三纪从老到新依次划分为古新世(Paleocene)、始新世(Eocene)、渐新世、中新世和上新世(Pliocene)5 个地质时期,再往前是中生代的白垩纪(Cretaceous)和侏罗纪(Jurassic)[8]。
茶树与宽叶木兰和中华木兰之间的关系实际上是山茶科与木兰科两个科之间的系统演化关系,属被子植物系统进化方面的学术问题。被子植物起源和早期演化的研究一直是进化生物学的研究热点,吸引了世界上许多顶级科学家穷其一生去探讨,迄今只在高级分类阶元(目级或科级)水平上提出了一些大致的演化框架,而“木兰化石”依据较少的文献资料、通过简单的研究方法、在种级水平上得出了精确结论。本文拟从植物系统分类学的角度对这一观点进行评述。
1 白垩纪被子植物大爆发是业内广为熟知和接受的观点,包括山茶科在内的被子植物各类群早在中生代的白垩纪就已经大量出现[9-12],用新生代第三纪的化石(而且仅仅只是中国的化石)来探讨被子植物科与科之间的系统演化关系欠专业
关于被子植物的起源,上世纪曾有“古生代起源说”和“白垩纪(或晚侏罗纪)起源说”两种观点[13]。近年来,随着研究手段和技术的更新,资料大量累积,特别是早期被子植物的花和其他生殖结构化石不断地被发掘,后一观点被广为承认和接受。上世纪末的化石证据“毫无疑问地表明被子植物最早出现在早白垩纪的巴雷姆期(Barremian)和更早的欧特里夫期(Hauterivian)”[14],近20 多年来,数个侏罗纪被子植物化石的发现将被子植物起源的时间推向了更早的地史时期[15-21]。近年来的分子钟估算结果表明,被子植物可能早在侏罗纪之前的三叠纪就已经起源[22-23]。现有化石资料表明,“在白垩纪距今90 ~125 百万年(~135 百万年)前,被子植物现代类群的不同演化水平及不同传代线的代表几乎都出现了”[24]。也就是说,包括山茶科和木兰科在内的被子植物各类群在白垩纪就已经起源了[9-12],(山茶)科与(木兰)科之间的系统演化关系发生在白垩纪早期甚至之前。宽叶木兰化石出土自白垩纪之后的新生代渐新世地层(距今23.5 ~33.7 百万年),不适合用作探讨科间系统演化关系的证据,“木兰化石”应该查阅《中国植物化石》的第二册(中生代植物),并在世界范围内查阅,不能仅限于中国。
2 山茶科的化石在白垩纪地层中就已出现,远早于宽叶木兰和中华木兰的生活年代,后者不可能是茶树的祖先
山茶科的化石从中生代的晚白垩纪到新生代的上新世在全球许多地方都有发现[25-59],花、果实、种子、叶和木材的化石均有,表1对已报道的山茶科大化石进行了粗略统计(按文献发表的顺序排列)。根据文献,最早的山茶科化石发现于晚白垩纪;有两个报道,其一是在埃及发现的类似于现今木荷属(Schima)植物的Schimoxylon dachelense木材化石[25],其二是分布在欧洲的紫茎属(Stewartia)和古木荷属(Palaeoschima)果实和种子化石[26]。与茶树更为亲缘的山茶属植物化石记录也不少,在欧洲,有保加利亚早渐新世的Camellia abchasica[27]、罗马尼亚晚中新世的Camelliasp.[28];在北美,有美国早渐新世的Camellia multiforma[29];在亚洲,记录更多,有日本晚始新世的Camellia abensis[30]、中新世的Camellia protojaponica[31]、Camelliasp.[32]和Camellia japonoxyla[33],韩国早中新世的Camellia japonoxyla[34],以及我国广西南宁地区渐新世晚期的Camellia nanningensis(南宁古茶)[35]和台湾桃园县早中新世的Camellia kueishanensis(龟山茶树)[36]。这些山茶科化石记录中超三分之二在“木兰化石”发表之前已经发表,“木兰化石”中写道“查阅了古植物等有关资料”,实际上,这些山茶科的化石资料文献均没有被引用,文中缺乏山茶科的化石资料。探讨茶树起源首先应研究它隶属的山茶属乃至山茶科的化石,而不是将茶树活植物与另一个科的化石直接比较。
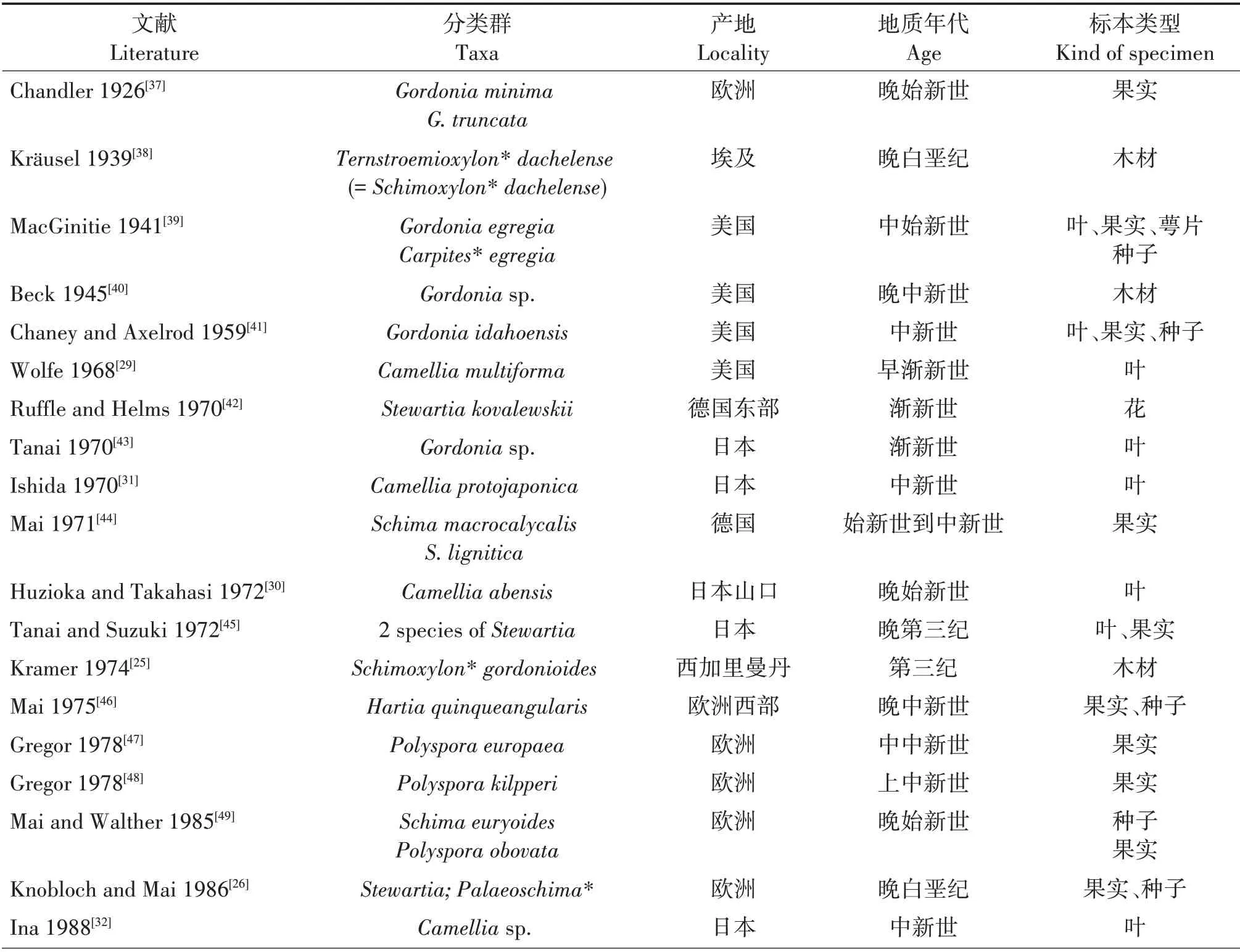
表1 山茶科(狭义)大化石记录Tab.1 Macrofossils of Theaceae s.str.
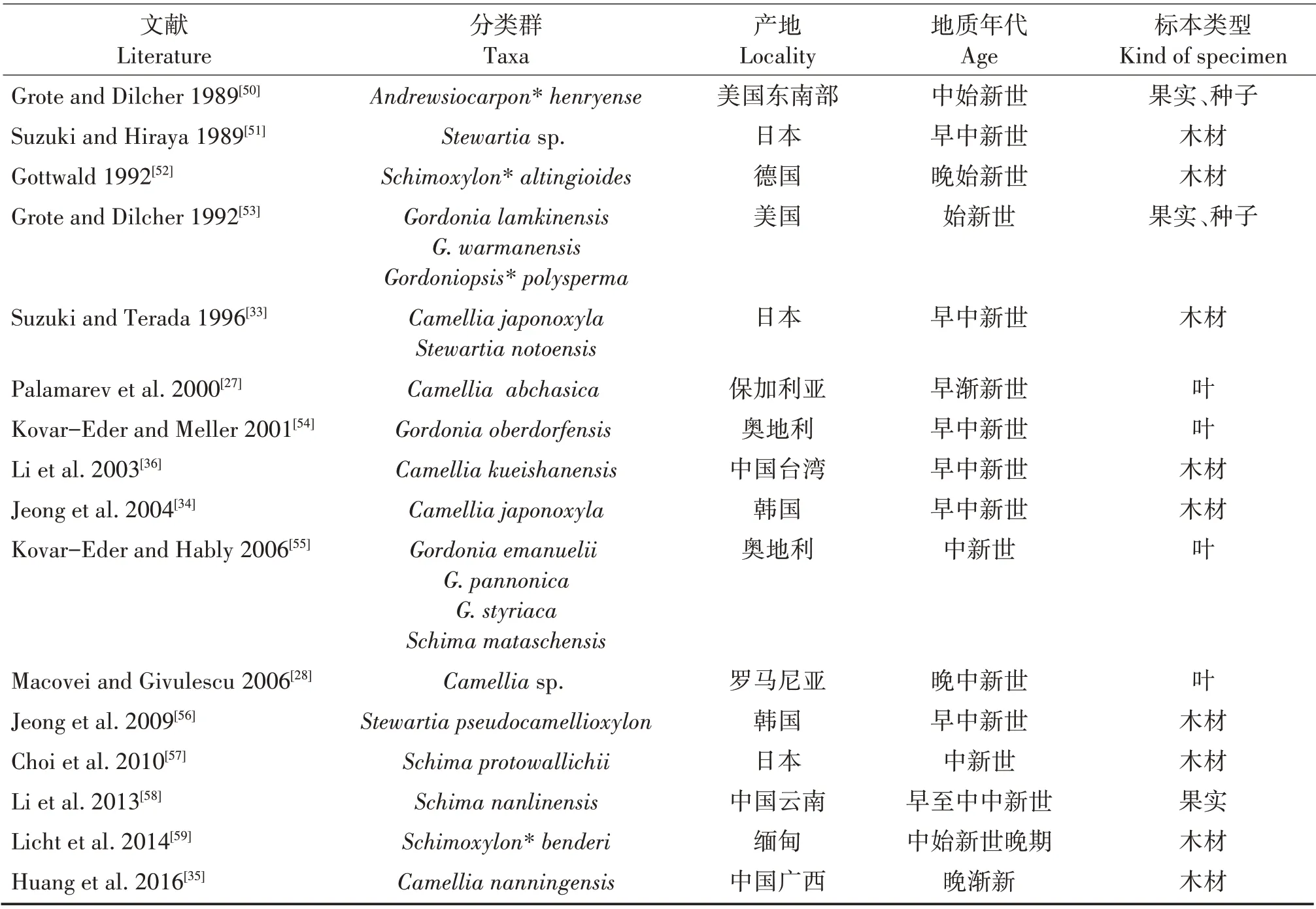
续表1 Continued
在我国发现的南宁古茶和龟山茶树均为木材化石,其原始文献中有详细的解剖学描述。山茶科的木材解剖特征曾有系统的研究和报道[60-61],因此,南宁古茶和龟山茶树鉴定的科学性和可靠性是有依据的。Liang and Baas(1990)[60]曾系统研究过山茶科的木材,取样涉及山茶科15属95种,其中40种为山茶属植物。依据木材解剖特征,这40种山茶属植物被划分为4 个特征群(group),其中特征群Ⅲ以茶组、连蕊茶组(Sect.Theopsis)和毛蕊茶组(Sect.Eriandria)植物为主。南宁古茶和龟山茶树的木材解剖特征均支持将它们归入特征群Ⅲ[35-36];其中,龟山茶树在射线的宽度和其导管缺少螺纹增厚上与茶树尤为相似(The fossil is similar toC. sinensisespecially in the ray width and the lack of spiral thickening in vessels)[36];南宁古茶具有已报道的茶(茶组植物,笔者注)、肖长尖连蕊茶(连蕊茶组植物,笔者注)和大理茶(茶组植物,笔者注)中具有的等宽多列和单列射线,证实了与这个特征群(特征群Ⅲ,笔者注)在解剖学上的亲缘(Their anatomical affinity to this group is confirmed by the occurrence of rays with equally wide multiseriate and uniseriate portions that have been reported forC.sinensisO.Ktze,C.subacutis-simaChang andC. taliensisMelch.)[35]。南宁古茶和龟山茶树是迄今报道的最为可靠、最为古老的与茶组植物亲缘关系最为接近的植物化石,分别与宽叶木兰和中华木兰几乎生活在同一地史时期,不可能存在相互之间链锁式的演化关系,是分别由各自的祖先演化而来的。南宁古茶和龟山茶树的发现说明,至少在晚渐新世(late Oligocene)(23.5 ~28 百万年前),与茶树十分类似的山茶属植物在中国的地界上就已经出现了。
贵州晴隆的“距今100万年左右晚第三纪至第四纪时期的四球茶(Camellia tetracocca)茶籽化石”[62-64]被直接鉴定为现代植物,且是在地面上捡到的,难以依据地层确定其确切的年代,在物种鉴定和地质年代鉴定上疑点重重。将其鉴定成四球茶的鉴别特征“化石的外形,大小,具种脐,其周边稍突起,种脐旁边有凹痕,种子顶端扁平或微突”在山茶属植物中是比较常见的,依此特征在现代山茶属植物的分类上连组都鉴定不到,更别说具体鉴定到种。即使鉴定正确,以100 万年左右的化石来论证茶树的起源也是不合适的,现存的被子植物在那个年代大都已经出现[65]。
3 山茶科与木兰科之间没有直接的演化关系,二者间的系统演化关系是相当疏远和复杂的,仅凭叶片的形态特征下不了结论
在几个有名的现代被子植物分类系统中,如哈钦松(1973)系统[66]、塔赫他间(1980)系统[67],木兰目(Magnoliales)确实是被放在系统演化的基部位置(多数现代被子植物分类系统并不持此观点[68-74]),但没有哪个系统认为山茶目(Theales)是由木兰目直接演化而来。在“木兰化石”依据的塔赫他间(1980)系统里(图1)(该系统自上世纪50年代提出后,一直在不断修订[70,75-76]),木兰目是通过五桠果目(Dilleniales)再演化到山茶目的。探讨木兰目与山茶目之间的系统演化关系需先搞清楚五桠果目在系统演化关系上是如何承前启后的,因此,五桠果目的研究材料不可或缺,“木兰化石”的这种“跳跃式”特征比对欠专业。由于起源历史的悠久以及化石资料的极度欠缺,现代植物系统学的研究通常是探讨最近的祖先,对远祖的推测是非常粗线条的。在哈钦松系统[66]和塔赫他间系统[67]里,在“目”这个分类阶元上,木兰目与山茶目之间的系统演化关系看似比较简单,但每个“目”包含若干个“科”(如在塔赫他间(1980)系统里山茶目由19 个科组成),科下的种类成百上千,种间的系统演化关系更为复杂,探讨不同科间种级水平演化缺失的研究材料就更多了。
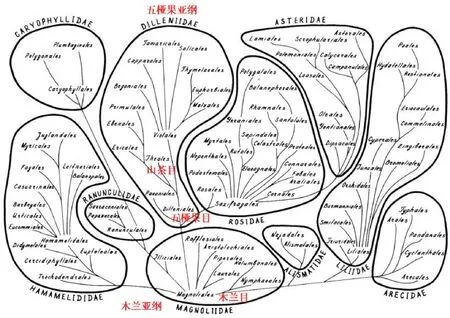
图1 塔赫他间(1980)被子植物系统(引自Takhtajan 1980) [67]Fig.1 Classification system of angiosperm(from Takhtajan 1980)[67]
目前,最为流行的以分子生物学证据为主要依据的APG 被子植物分类系统揭示了木兰科与山茶科间更为疏远和复杂的系统演化关系。图2是最新的APG Ⅳ被子植物分类系统[77]的基本构架,木兰科位于该系统的“木兰分支(Magnoliids)”中,包括白樟目(Canellales)、胡椒目(Piperales)、木兰目和樟目(Laurales)4 目,木兰目含木兰科等6 科。山茶科被放在在菊分支(Asterids)的杜鹃花目(Ericales)里,杜鹃花目含山茶科等22个科,图3是APG Ⅳ系统揭示的杜鹃花目内的系统演化关系[77]。由此可见,仅在科级水平上,山茶科与木兰科的系统演化关系就非常复杂,种级水平的系统演化关系就更加密如蛛网,盘根错节,不是“木兰化石”的研究方式能够解决的。
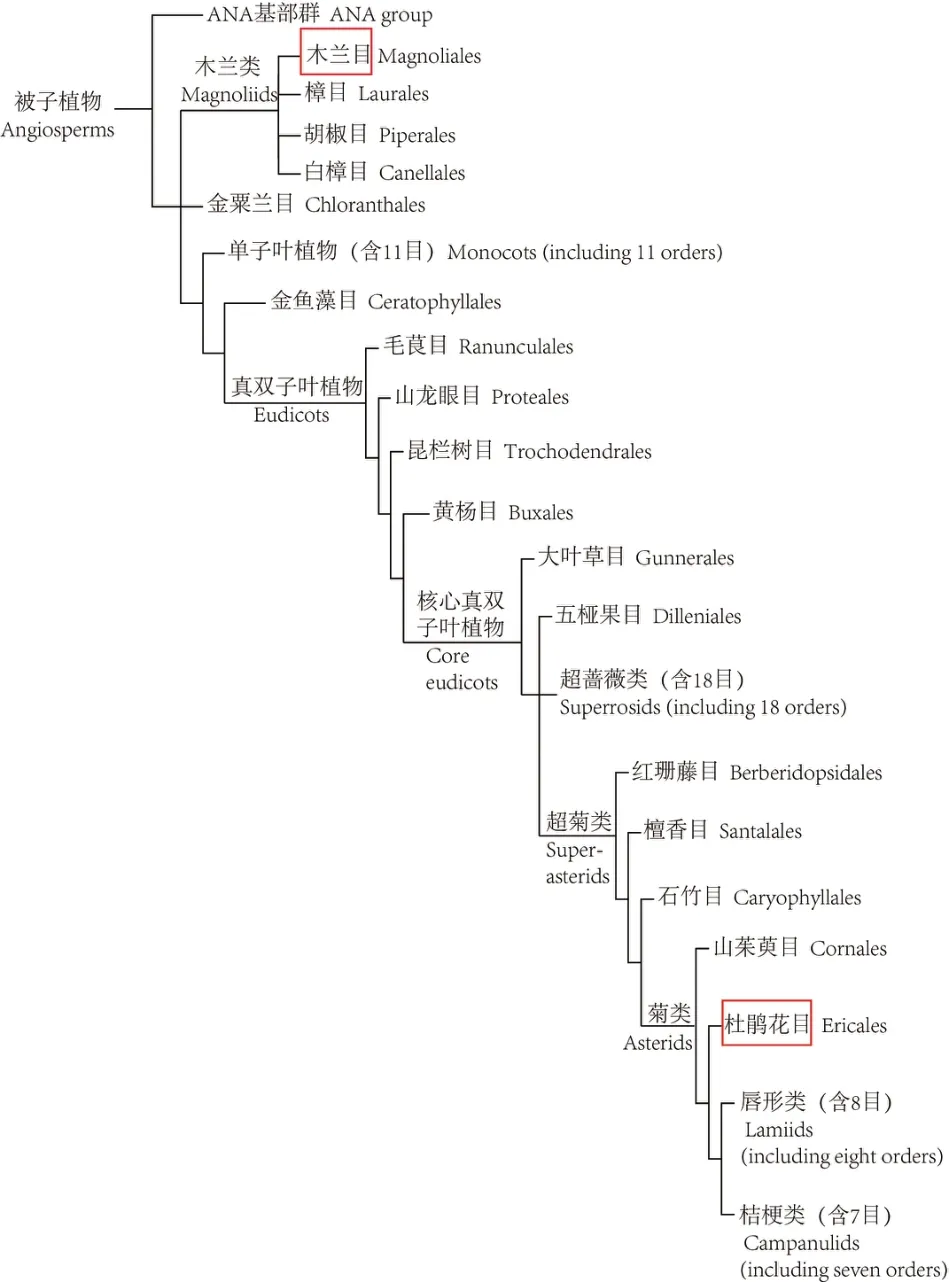
图2 被子植物系统发育(蒋银子据APG Ⅳ,2016绘制)Fig.2 Phylogeny of angiosperm(by Jiang Yinzi according to APG Ⅳ,2016)
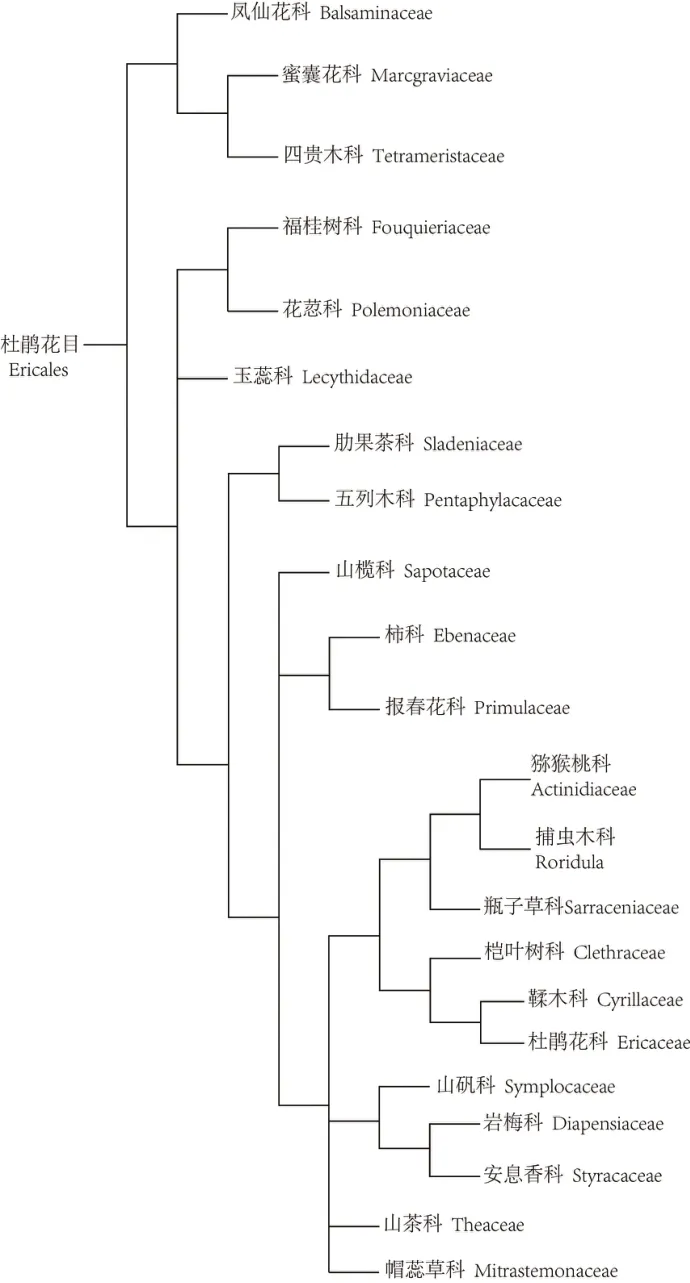
图3 杜鹃花目的系统发育(蒋银子据APG Ⅳ,2016绘制)Fig.3 Phylogeny of Ericales(by Jiang Yinzi according to APG Ⅳ,2016)
APG 系统另一个值得注意的结果是:木兰科不是最原始的被子植物,被子植物的基部类群由无油樟目(Amborellales)、睡莲目(Nymphaeales)和木兰藤目(Austrobaileyales)构成。那些基于“木兰(科)是被子植物之源”从而推断茶树起源于木兰科的观点[1,78]需重新考虑和修正。
4 现代茶树与宽叶木兰和中华木兰在叶片的形态结构上并非“有较多的相似之处”,并且,叶片属营养器官,受环境的影响较大,其特征的相似性更多反映的是在生态环境上的相似,在系统演化上的意义多局限在科以下比较低级的分类阶元,不适合用于亲缘关系很远的木兰科和山茶科之间系统关系的探讨
从特征描述来看[1],除“侧脉不达叶缘,在近叶缘处网结”外,宽叶木兰和中华木兰与茶树的叶结构没有更多实质性的相似之处。两个化石植物叶缘全缘,而茶树叶缘有明显的锯齿,差异非常明显。更值得注意的是,“脉不达叶缘,在近叶缘处网结”这一特征并不是两种化石植物和茶树独有的,在现代植物中,木犀科(Oleaceae)、山矾科(Symplocaceae)和冬青科(Aquifoliaceae)等科中有相当多植物叶的形态结构与茶树相似,且分布在同一区域,依“木兰化石”推理,这些植物的起源均可追朔到宽叶木兰。同理,化石植物中具有类似脉序结构的也不仅是宽叶木兰和中华木兰,木兰科的化石也不仅仅只有中国有,按“木兰化石”的研究方法,茶树可能有多个祖先。
宽叶木兰和中华木兰均为叶片化石,保存并不完全,能提供的特征信息非常有限和欠精细。目前,山茶科和木兰科比较精细的叶结构(Leaf architecture)分析已有相当多的研究积累[79-84]。孙航和闵天禄(1995)[82]报道了山茶属108 个种的叶结构分析结果,取样几乎涵盖了当时茶组的所有种类,是一个非常全面的研究;Pray(1954)[79]、Dickison(1975)[80]、Yu and Chen(1991)[81]、Liao et al.(2000)[83]和Zhang and Xia[84]等也报道了不少木兰科叶结构分析的研究成果。这些文献不少在“木兰化石”之前已经发表,但这些现代植物叶结构方面的研究“木兰化石”均没有引用,化石方面也只引用了少量国内的化石资料,而且还是被子植物起源之后相对年轻的新生代化石资料。由此可见,“木兰化石”的基本文献储备较欠缺。
在生物的起源和演化研究中,化石证据确实很重要,甚至是关键证据。但是,化石的形成和保存状态与特定的地史条件密切相关,化石的发现又是一个极具偶然性的过程,动植物系统演化上大部分化石证据的缺失是一个不争的事实。因此,不了解现代植物的形态特征和地理分布,仅凭化石证据去推论一个生物类群的起源通常是难以成行的。在化石证据不足的情况下,在植物地理学上,通常将类群内原始种类最集中和多样性最丰富的地区推测为这个类群的起源中心[85]。依此原理,闵天禄(1992)[86]提出,滇黔桂交汇地带是茶组植物的起源中心,云南是茶组植物起源中心的一部分,这是目前关于茶组植物起源最为专业的论证。
植物的起源、类群间的系统演化关系是十分重要和极其复杂的科学问题,需有真凭实据,有专业的研究方法和思路,需在查阅和掌握大量专业文献资料的基础上进行研究,力求科学性、专业性和真实性。
致谢:衷心感谢《普洱》杂志社段兆顺先生、中国普洱茶研究院王兴华先生、普洱市茶办刘伦先生提供部分文献资料;衷心感谢本单位硕士研究生蒋银子为本文图版的制作提供帮助。
猜你喜欢
教学月刊(中学版)(2022年12期)2022-12-27
小哥白尼(神奇星球)(2021年3期)2021-07-22
故事作文·高年级(2021年2期)2021-02-04
科学(2020年5期)2020-11-26
广东园林(2020年3期)2020-07-03
文苑(2020年6期)2020-06-22
青年生活(2020年6期)2020-03-28
动漫星空(兴趣英语)(2019年5期)2019-05-14
东坡赤壁诗词(2018年4期)2018-11-07
科学启蒙(2016年4期)2016-03-23
