
3~6岁儿童动物共情的发展及其与人际共情的关系研究①
2022-07-09 07:29阳泽董佳琦
西南师范大学学报(自然科学版) 2022年7期
阳泽, 董佳琦
1.西南大学 教育学部, 重庆 400715; 2.深圳市龙岗区特殊教育学校, 广东 深圳 518116
共情是人与动物共同具有的、 与生俱来的能力. 共情最初源于母性行为, 即对后代的痛苦作出回应的能力, 经过自然选择、 演化发展, 因之既能增加后代的生存机会, 又能确保基因的延续[1], 具有重要的生物适应性, 所以在自然进化中得以保留. 共情广泛存在于相同物种、 不同物种之间, 同种动物间、 跨物种动物间的共情证据发人深省, 这表明共情不仅存在于养育环境中, 同时也能迁移到该环境之外. 人类的共情不单靠血缘、 亲缘的维系, 更延伸至靠地缘和业缘维系的社会环境, 甚至是与自己毫无关联的他者或它者. 如庚子2020年的新冠肺炎疫情来袭, 各地积极支援湖北[2]; 人类共情沙滩上搁浅的海洋生物, 并将其放回海洋. 此时, 共情已然超出了进化起源的范畴.
动物共情是人类个体由关注自身的“唯我主义”转向关注“它者”, 是人类个体对动物的处境及其心理感同身受的能力. 动物共情的研究源于21世纪初人类对人与动物关系的重新审视, 其目的在于改善人类对待动物的态度、 提升动物福祉[3]. 这种能力能够发展个体亲动物的态度, 重构人们对生命的感悟, 维护生物多样性, 推进动物保护运动, 促进人与自然和谐共生. 共情这一神奇的能力使人、 动物都不再是孤立的存在, 映射出人与动物对等的共生关系. 人与动物共处于一个生态系统[4], 是一种相互依存的共生关系, 即人为动物提供有利于生存的帮助, 动物也能反作用于人, 为人提供有利于生存的帮助, 这种关系具有生物意义上的适应性[5]. 共生是群体生存的重要法则, 个体无法离开群体而独善其身, 任何一个个体都是群体中的个体, 群体中的个体之间如果不能共生, 个体会衰亡, 群体也会随之衰落乃至消亡. 因此, 共生于任何群体的生存发展都至关重要[6]. 通过共生可以实现优势互补、 共赢发展. 一方面, 人类对动物共情, 体会动物的不利处境, 致力于保护动物赖以生存的家园, 关爱处于困境中的动物, 保护生物多样性, 促进人与自然的可持续发展; 另一方面, 动物可以是人类的老师、 治疗师或伙伴, 以文化智慧和道德教诲的形象出现在谚语或寓言中, 以治疗师的角色出现在世界各地的医院、 疗养院、 福利院, 以伙伴或朋友的身份成为人们生活、 工作、 学习中必不可少的成员.
3~6岁是儿童人际共情发展的关键期[7], 有研究表明[8], 3岁时儿童的人际共情开始萌芽, 愈加关注他人需要, 能够理解他人的情绪情感有别于自身. 该阶段儿童共情的发展建立在自我和情绪发展的基础上, 一方面, 儿童自我发展包括自我概念的建立和自我价值感的萌芽. 儿童自我发展经历了从以自我中心到指向他人的质的飞跃, 儿童自我概念的建构随着年龄的增长而不断完善和清晰; 其次, 自我价值感在儿童期萌芽, 是儿童对于自身价值的判断, 在4岁时儿童就有了自我判断, 只不过这种判断存在缺陷, 它不是基于对自身准确、 完整的认识, 所以判断可能不准确或存在偏差[9]. 另一方面, 除了自我的发展, 儿童的情绪能力也有极大的发展, 表现为自我意识情绪、 情绪理解和情绪调节. ①3岁时儿童自我意识情绪出现, 如儿童在冲突后感到生气, 在成功后感到高兴, 随后, 儿童自我意识情绪渐进发展, 儿童判断情绪的标准会逐渐内化, 并影响对自身的看法; ②情绪理解即儿童清楚地知道他人当前的情绪状态, 譬如, 4~5岁的儿童能够根据他人表现, 推断他人情绪; ③儿童情绪调节能力逐渐发展, 体现在儿童能够有效应对消极情绪, 控制不良情绪.
动物共情与人际共情关系复杂, 一方面, 动物共情和人际共情可能具有同源性, 即有某一机制同时作用于动物共情和人际共情或是动物共情和人际共情就是同一共情结构的两个方面. 譬如: 早在1972年Mehrabian等[10]编制的情感共情量表将动物共情纳入了条目中, 动物共情条目在其量表中信效度良好[11]. 该工具基于这样一种假设, 即个体的共情水平越高, 人际共情、 动物共情的水平越高; 反之, 个体共情水平越低, 人际共情、 动物共情水平越低[12]. 进一步的实证研究结果显示, 动物共情与人际共情存在显著但低度的正相关, 即对动物友好或共情的人可能对人也有类似的正向的情感[13]; 另一方面, 个体可能先获得动物共情或人际共情中的某一种, 在其他因素的作用下共情能够发生迁移[14]. 有研究者[15]认为个体容易最先产生动物共情, 且动物共情能促进人际共情, 因此鼓励个体与动物接触来提升人际共情. 有的国家还开展了“共情之根”等人文教育项目, 旨在提升儿童的社会性发展. 通过培养儿童对动物共情, 间接提升儿童对人的共情, 以减少儿童的攻击性行为, 促进儿童的同伴交往、 亲社会行为、 利他行为、 分享行为[16]. 此外, 相似性可能在动物共情和人际共情间起到中介作用. 以往研究[17]发现, 感知相似性在群体内认同和心理融合中起到了中介作用. 相似性能够增加个体对自我、 它者、 他者的认同, 缩短了自我、 它者和他者的群际心理距离, 将“我们”扩大, 促进个体对它者或他者的共情. 因此, 动物共情与人际共情的关系复杂, 二者是否具有同源性或是否存在迁移需要进一步深入探究.
1 研究方法
1.1 研究对象
采用目的抽样结合随机抽样的方式, 选取农村、 城市的2所幼儿园, 在各幼儿园抽取小、 中、 大班各2个班, 共406名儿童, 剔除无效数据27份, 最终得到有效数据379份, 有效率为93.35%. 其中男孩197人, 女孩182人; 农村儿童168人, 城市儿童211人; 3岁组儿童87人, 3.5岁组儿童24人, 4岁组儿童96人, 4.5岁组儿童41人, 5岁组儿童86人, 5.5岁组儿童28人, 6岁组儿童17人.
1.2 研究工具
1.2.1 动物共情
采用修订的3~6岁儿童动物共情工具. 动物共情工具根据Janet Strayer的共情连续体修订而来. ①梳理已有文献, 初步认为情境故事测验法适用于我国3~6岁儿童, 即通过虚构的动物情境故事来诱发儿童动物共情. ②根据已有文献, 情境故事从高兴、 伤心、 生气、 害怕4个维度展开, 每个维度下虚构3个情境故事. ③对不同年龄段的儿童及学前教育专业学生进行测验, 最终筛选出情绪易识别、 情绪程度强、 熟悉度强的4个情境故事, 以确保情境故事能作为有效的诱发材料. 在每个情境故事的问题设计上参照Jenet Strayer的共情连续体工具, 通过5个问题考察儿童在不同情感维度上的动物共情. 经检验, 该工具的重测信度为0.937, 通过因子分析, 析出一个主成分, 解释变异量为63.7%, 积差相关分析发现该工具的效标关联效度良好. 因此该工具信效度良好, 使用该工具测量3~6岁儿童动物共情可行且有效.
1.2.2 共情连续体
采用Janet Strayer共情连续体工具(Empathy Continum), 对3~6岁儿童人际共情进行情境故事测验, 此工具包含人际共情的情绪情感得分和经过认知调节的共情得分. 该工具适用于儿童. 苏金莲对工具中的情境故事进行了一定的修改, 使其表述适用于中国儿童. 翟晓婷参考该工具, 对幼儿进行了情境故事测验. 本研究采用翟晓婷修订的Janet Strayer共情连续体工具, 对3~6岁儿童人际共情进行情境故事测验.
1.2.3 人类—动物相似性检验
参考Baston和Catherine的感知相似性测量方式, 个体报告他们认识到人类与动物之间的相似性越多, 对动物的共情就越强. 通过口头询问3~6岁儿童认为自己与故事中的角色是否相像来检验, 采用3点计分, “不相像”得0分, “有点相像”得1分, “完全相像”得2分. 而后通过询问儿童哪里相像来做进一步的了解. 同时, 在所有故事讲述完后, 研究者会问儿童自己与动物有哪些相同之处以及不同之处.
1.3 研究程序
在研究开始之前, 研究者会主动征得儿童本人及儿童家长的知情同意. 在儿童进行区角活动时主动参与到儿童的活动中, 并问儿童是否喜欢听故事, 将其带到教室安静的一角, 告诉他听完故事后可以再回来玩. ①研究者出示4张表情图片, 问儿童是否认得这些表情, 如果不认识, 研究者会主动告诉儿童不同表情图片的含义, 直到儿童能准确识别4种情绪. ②给儿童讲述故事, 故事讲完后让儿童复述故事, 确保儿童听清每个情境故事中发生了什么事情以及故事中的主要角色, 在讲故事的过程中辅以图片, 帮助儿童更好地理解故事内容. ③问儿童问题, “故事中的主人公(动物、 人)的心情如何?”伴以表情图片辅助儿童作答, 当儿童提到的情绪不止一种, 研究者会让儿童说出最主要的情绪, 然后儿童需要对自身的情绪及情绪强度作出判断, 并说出自己产生这种情绪的原因. ④对儿童进行人类—动物相似性检验. ⑤儿童可以选择一个自己最喜欢的贴纸, 然后回到之前的活动区角中.
1.4 数据处理
采用SPSS 25.0、 Amos 22.0统计软件对可量化的数据进行分析与处理. SPSS 25.0主要用于信度检验、 差异分析及相关关系检验等, Amos 22.0主要用于中介效应检验.
2 研究结果
2.1 3~6岁儿童动物共情的发展特点
图1可知, 3~6岁儿童动物共情的发展总体上呈递增趋势, 随年龄的增长而增长. 就发展速度而言, 3~6岁儿童动物共情的发展大致可以分为3个阶段: 3~3.5岁为萌芽期, 3.5~5岁为快速发展期, 5~6岁为平稳发展期. 萌芽期的儿童动物共情总体得分在1分以下, 趋近于0分但又不等于0分, 表明儿童无法感知故事中动物的情绪情感, 或是能够感知但自身情绪与故事中动物的情绪不匹配, 此阶段的儿童无动物共情. 快速发展期的儿童, 动物共情总体得分从1分快速上升至5分, 表明儿童已经产生了动物共情, 且对自身的情绪归因从无关归因发展至能够基于故事的事件进行归因. 平稳发展期的儿童动物共情总体得分稳定在5~6分, 该阶段保持了基于故事的事件进行归因. 在3~6岁的年龄段, 3.5岁是儿童动物共情的重要转折期, 此时儿童动物共情总体得分从1分以下转变为1分以上, 即从无动物共情向有动物共情转变, 实现了质的变化. 此外, 各维度的动物共情发展趋势相当, 相较于其它3个情感维度, 儿童动物高兴共情水平更高.
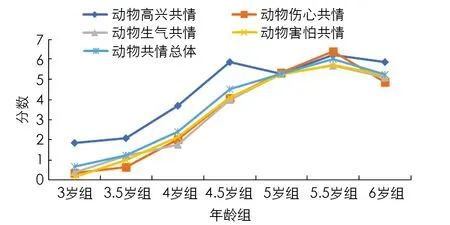
图1 3~6岁儿童动物共情发展的年龄趋势
2.2 3~6岁儿童动物共情发展的差异
由表1可知, 3~6岁儿童动物共情在地区上的差异有统计学意义, 城市儿童的动物共情在高兴、 伤心情感维度及动物共情总体上显著高于农村地区的儿童. 3~6岁儿童动物共情在性别及是否为独生子女上的差异无统计学意义.
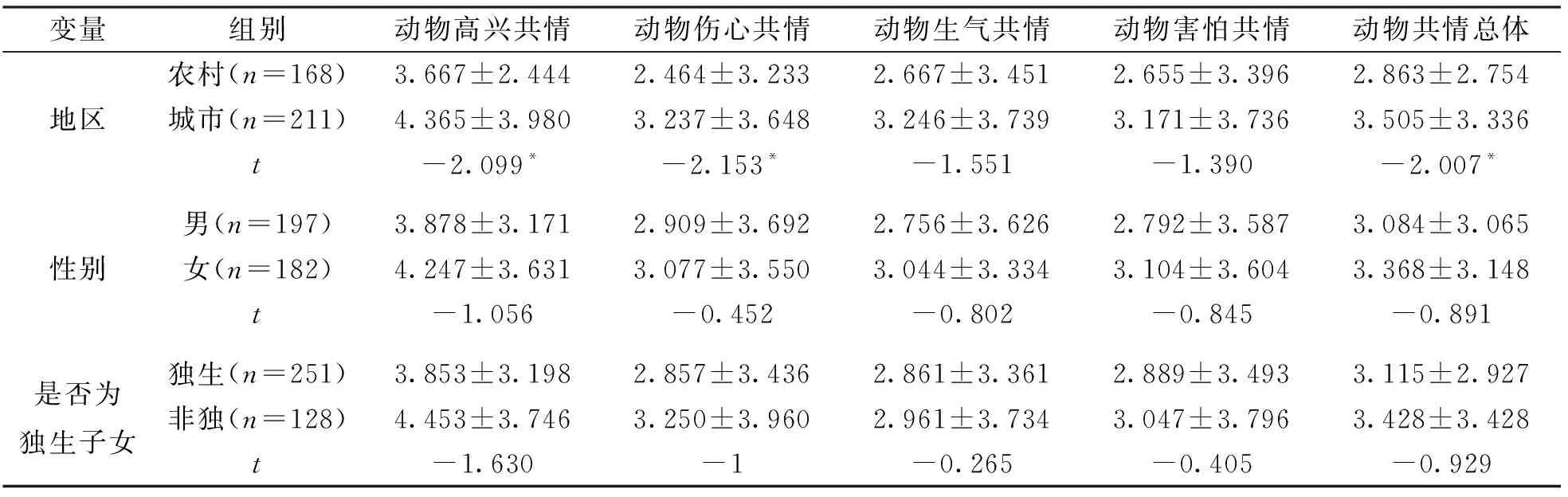
表1 3~6岁儿童动物共情的差异分析(n=379)
2.3 3~6岁儿童动物共情和人际共情发展的相关分析
对3~6岁儿童动物共情、 人际共情进行相关分析. 由表2相关矩阵可知, 儿童动物共情在高兴、 伤心、 生气、 害怕各维度及总体上与人际共情显著正相关.

表2 3~6岁儿童动物共情、 人际共情的相关矩阵
2.4 相关关系随年龄的变化情况
采用相关分析考察儿童在不同年龄段上的动物共情和人际共情之间的相关, 如表3所示, 各年龄段儿童人际共情总体与动物共情总体均显著相关, 3岁组相关系数偏低, 3.5~5岁组相关系数相对平稳, 5.5~6岁组相关系数降低.

表3 不同年龄段儿童动物共情与人际共情之间的相关分析
2.5 人类—动物相似性、 动物共情和人际共情的关系
2.5.1 人类—动物相似性、 动物共情、 人际共情的相关分析
如表4所示, 3~6岁儿童动物共情、 人际共情、 人类-动物相似性三者之间的相关分析结果显示, 3个变量间两两均显著相关, 满足进行中介效应的前提, 可进行中介效应分析.

表4 动物共情、 人际共情、 人类-动物相似性的相关矩阵
2.5.2 人类—动物相似性在动物共情和人际共情关系之间的中介作用
以动物共情为预测变量, 人际共情为效标变量, 人类—动物相似性为中介变量建构模型, 用极大似然法(Maximum likelihood)对假设模型进行检验, 各项拟合指数如表5所示, 卡方自由度比值为2.905, 介于1~3之间, 说明假设模型与样本数据的契合度可接受; 渐进残差均方和平方根为0.076, 数值界于0.05~0.08, 模型良好, 有合理适配.

表5 相似性的中介模型拟合指数
形成的拟合模型如图2所示, 动物共情的4个测量指标“高兴”“伤心”“生气”“害怕”纳入模型进行作用. 用偏差校正的非参数百分位Bootstrap程序检验中介效应显著性, 随机共抽取5 000组样本, 置信区间设置为95%, 计算出总效应、 各路径系数以及间接效应的估值.
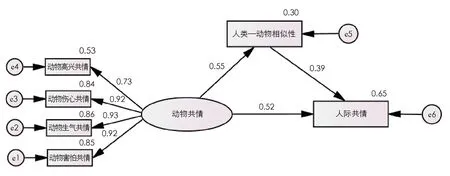
图2 相似性在3~6岁儿童动物共情与人际共情的中介效应模型
由表6可知总效应、 间接效应、 直接效应的具体情况, 动物共情→人类—动物相似性→人际共情的中介效应所在区间是0.153~0.286, 不包含0, 作用大小为0.215, 达到0.000的显著水平, 表明效应具有统计意义和实际意义[18]. 直接效应的所在区间0.419~0.625, 不包含0, 说明加入中介变量后, 直接效应具有统计学意义. 即人类—动物相似性在动物共情和人际共情之间存在部分中介效应, 中介效应占总效应的比值为29.1%.
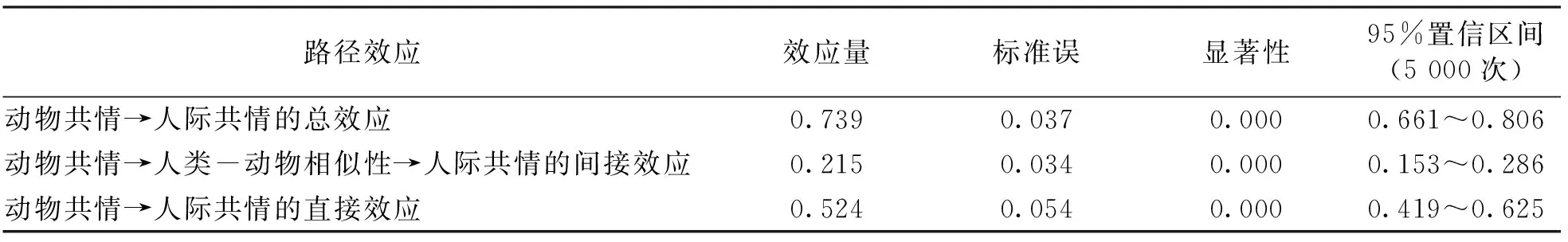
表6 Bootstrap法的中介检验
3 讨论
3.1 3~6岁儿童动物共情的发展特点分析
3~6岁儿童动物共情随年龄的增长而增长, 这一结果与Emde对共情的研究结果相同, 也与陈杰[19]的人际共情的研究结果一致. 说明动物共情的发展与人际共情一样, 会受到个体自身成长的阶段性和发展性的影响. 共情能力贯穿于整个生命周期, 而在生命历程中, 共情与自我意识、 道德成熟、 心理意志、 情绪情感、 认知能力等平行发展[20]. 动物共情是一种技能, 在儿童早期随着年龄的增长, 儿童逐渐有了自我概念、 道德判断能力提升、 知识不断丰富, 因而能更敏感而准确地洞察身边的事物, 表现为理解关心除自身以外的它者.
在发展速度方面, 3.5岁以前的儿童动物共情发展缓慢, 3.5~5岁的儿童动物共情发展较为迅速, 而5岁以后的儿童动物共情保持平稳发展, 速度的变化可能是因为儿童情绪情感的发展和认知水平的提升. 一方面, 从3岁开始, 儿童陆续产生多种情感[21], 依据情绪分化理论, 3.5岁以前儿童的动物共情主要涉及情绪情感的变化, 此时, 儿童情绪情感快速发展, 儿童情绪情感的变化是面对刺激的适应状况. 另一方面, 3.5~5岁期间, 儿童动物共情认知成分增加. 3.5~5岁阶段儿童符号系统开始形成, 不再靠单纯的感知觉来认识这个世界[22], 儿童除了简单的情绪变化, 也试图去理解自己产生情绪变化的原因, 表现在动物共情上为快速发展. 5~6岁的儿童认知和情感发展较为稳定, 因此表现为动物共情的稳定发展.
3.5岁是儿童动物共情从无至有转变的关键阶段. 依据霍夫曼的共情发展模型, 3岁的儿童有了共情的所有基本成分, 但这只是简单的共情, 更为复杂和成熟的共情能力需要儿童理解它者的情感状态并作出相应的情感反应[23]. 3岁以后儿童情绪情感日渐丰富和成熟, 3~4岁是儿童情绪理解能力的关键期[24], 3.5岁的儿童正处于此关键阶段, 儿童能够了解到自己与他人有不同的情绪和想法[25], 这为儿童动物共情的发展做了铺垫. 此外, 3岁以后儿童认知逐渐发展, 观点采择对儿童来说是十分复杂的任务, 3~3.5岁儿童的观点采择初步发展[26]. 随着个体观点采择能力的提高, 认知共情能力才能日渐发展成熟[27]. 儿童逐渐从无法理解动物, 到能够站在动物立场上感同身受, 就需要观点采择能力.
在各维度得分上, 高兴维度得分较其它维度略高, 这可能与儿童情绪识别的顺序有关. 具体情绪识别的发展呈现一定的顺序性, 从早到晚依次为高兴、 伤心、 生气、 害怕[28]. 儿童识别高兴情绪较早, 识别伤心、 生气、 害怕情绪较晚. 儿童对高兴的原始情绪认知最早, 这是因为它是人类在婴儿时期就获得的情绪, 人类婴儿时期是极度软弱无助的, 若离开成人的保护就难以成活, 于是, 人类婴儿必得以某种方式取悦母亲, 笑就成了婴儿的适应状[29]. 而儿童的伤心、 生气、 害怕等情绪发展较迟, 因此儿童的动物共情在这3个维度的得分较低.
3.2 3~6岁儿童动物共情的发展差异分析
城市儿童的动物共情水平显著高于农村儿童. 在动物共情总体及高兴和伤心维度上, 城市儿童的动物共情水平显著高于农村儿童, 而在生气和害怕维度上未达到显著性水平. 在总体上城市儿童得分高于农村儿童得分, 可能的原因是城市中的儿童能享受更优质的教育资源, 经历更丰富, 通过动物园、 博物馆等媒介认识到更为多样的动物, 认知水平更高, 因此对于自身产生的情感变化有更合理的归因; 而农村的交通较为闭塞, 与外界的联系更少, 教育资源匮乏, 父母文化水平偏低, 儿童的认知水平偏低, 归因浮于表面. 在高兴和伤心维度上存在显著差异, 可能是因为高兴和伤心是儿童最先发展起来的情绪, 儿童能够识别相应的情绪, 但因为认知水平的差异, 农村的儿童听完情境故事只是觉得故事好听, 无法对自己的情感进行合理的归因. 而生气和害怕是更为复杂的情绪, 儿童发展的较迟, 因此在这2个维度上不存在显著差异.
男孩和女孩在动物共情总体及各维度上无显著差异. 这与Angantyr等[30]的研究结果一致, 未见显著的性别差异, 但与国外动物共情研究的结果不一致, 女性明显表现出更高水平的动物共情[31]. 在此, 可能的解释为教育者对男孩或女孩的教育方式一致, 并未区别对待[32]. 当前我国在性别角色上提倡双性化教育, 打破传统的性别刻板印象, 在儿童早期的性别教育上就提倡吸收异性优秀的性别特质, 儿童也就具备了更多积极的人格特征, 如有爱心、 富有同情心、 将心比心等.
3~6岁儿童动物共情总体及各维度在是否为独生子女上无显著差异, 表明是否为独生子女对儿童动物共情的影响不大. 以往研究认为儿童的共情倾向可能会通过其他敏感性的事物或是关心的角色榜样(如同胞)而增强, 但本研究结果显示, 同胞对儿童动物共情影响不大. 一方面, 父母教养方式改善, 育儿更加科学合理, 无论是有几个孩子, 父母都能做到一碗水端平, 不偏不倚; 另一方面, 随着物质生活水平的提升, 同胞间不会因为资源缺乏而产生恶性竞争, 同胞关系和谐. 因此, 独生子女与非独生子女的动物共情未出现明显分化.
3.3 3~6岁儿童动物共情和人际共情的关系分析
3.3.1 3~6岁儿童动物共情和人际共情显著正相关
相关分析的研究结果显示, 动物共情总体及各维度与人际共情显著正相关, 这与Paul的研究结果一致. 由此可知, 对动物有较高共情力的个体, 对人的共情能力也较高. 这可能是因为动物共情和人际共情有共同的潜在特性或是动物共情和人际共情都是通过同一机制来调节. 一方面, 共情的概念源于非人类环境, 即个体对一件艺术品共情[33]. Eisenberg[34]认为共情同等地适用于动物目标和人类目标, 无论共情的目标是什么, 个体的共情水平通常是稳定的, 因此动物共情和人际共情的水平大致相当. 根据个体对于不同的目标做出类似的反应, 可以推测无论是动物共情还是人际共情, 可能都存在一些潜在的共同特性[35], 这可能与个体早期成长环境或是基因息息相关[36-37]. 另一方面, 动物共情和人际共情可能存在共同的调节机制, 在具体情境下由具体目标所引发的共情可能都通过某种机制来调节, 这种机制能够增强共情反应[38].
3.3.2 相关关系随年龄的变化而变化
儿童人际共情总体和动物共情总体在不同年龄段上显著相关, 但3岁组的相关系数偏低可能与儿童的自我意识有关. 依据拉康的镜像理论, 3岁的儿童无法区分“我”与“物”, 且该阶段儿童以自我为中心, 儿童的自我意识处于混沌状态[39], 边界尚在建立, 无论是动物共情或是人际共情都是儿童自身做出的一种模糊的判断. 而随着个体自我意识的发展, 5.5~6岁组儿童已经能够区分自我和它者, 进而能作出准确的判断, 明晰自我、 他者、 它者的区别与联系. 因而, 5.5岁组、 6岁组的人际共情和动物共情相关系数稍有降低.
3.3.3 人类—动物相似性在动物共情与人际共情关系之间的中介作用
对3个变量的相关分析结果显示, 动物共情、 人类—动物相似性、 人际共情两两显著正相关, 动物共情通过直接和间接两条路径作用于人际共情. 一方面, 动物共情可以直接作用于人际共情. 在共情的对象间存在一种迁移, 威尔逊的亲生物假说认为儿童有亲生物性, 儿童天生亲近动物[40], 儿童与动物的亲密情感会促使儿童与动物形成稳定的情感联结, 这为动物共情奠定了深厚的情感基础. 动物成为了儿童自我意识的重要参考点[41], 通过与动物的积极互动, 提升了儿童对动物设身处地的共情能力. 库利的“镜中我”理论中有一种镜像效应, 动物对儿童的态度像一面镜子能照出儿童的形象, 儿童会依此形成自我概念的印象, 会对动物眼中的自我影像产生认同, 促进了儿童对自我的理解. 随着儿童对自我的理解, “自我”的疆域会不断扩大, 逐渐将“它者”涵盖进来, 动物与人都是有生命的事物, 是除自我以外的它者, 而自我可能会泛化到一切有生命的事物[42], 儿童对动物的共情会推及儿童对人的共情, 人际共情能够帮助儿童与人建立满意的关系, 促进儿童社会性的发展. 另一方面, 3~6岁儿童的动物共情会通过提升儿童的人类—动物相似性感知进而提升儿童的人际共情水平, 中介效应占总效应的29.1%, 换言之, 3~6岁儿童的动物共情水平越高, 则儿童所感知到的人类与动物的相似之处越多, 从而提升了儿童的人际共情水平. 人类和动物同处于一个生态系统, 人类和动物同属于除自身以外的它者或他者. 与动物的非语言交流提升了儿童通过非语言线索了解他人内心的能力, 这就需要动物共情, 当动物共情产生时, 与动物的联系赋予个体与他人沟通和共鸣的选择性优势[43]. 相似性是人际共情的主要影响因素[44], 处于社会中的儿童在社会交往中能够逐渐发现人类和动物在外貌、 生活习性、 交往等方面的相似之处, 个体对人类—动物相似性的感知提升, 人类—动物相似性的感知调控了个体对人类的心理感受, 缩短了个体与人类之间的心理距离, 将他者与自我联结在一起, 减少了偏见和歧视, 增加了对他者的认同. 当儿童渐渐发现自己与他者的某些相似之处或生命的同源性就能够表现出对他者的人际共情.
4 结论
1)3~6岁儿童动物共情发展随年龄的增长呈递增趋势. 3~6岁期间的发展可划分为3个阶段, 分别是3~3.5岁的萌芽期, 3.5~5岁的快速发展期, 5~6岁的平稳发展期; 其中, 3.5岁是一个重要的转折期; 各维度的动物共情发展趋势相当, 相较于其他3种情感维度, 儿童动物高兴共情水平更高.
2) 城市儿童的动物共情在高兴、 伤心维度及动物共情总体上显著高于农村儿童.
3) 3~6岁儿童动物共情与人际共情显著正相关. 3岁组相关系数偏低, 3.5~5岁组相关系数相对平稳, 5.5~6岁组相关系数降低; 人类—动物相似性与动物共情和人际共情显著正相关, 且在动物共情对人际共情的关系中起中介作用.
猜你喜欢
商业评论(2022年4期)2022-05-05
天津外国语大学学报(2021年3期)2021-08-13
小学生学习指导(当代教科研)(2021年6期)2021-05-23
教书育人(2020年11期)2020-11-26
甘肃教育(2020年17期)2020-10-28
河北画报(2020年8期)2020-10-27
闽南师范大学学报(哲学社会科学版)(2020年4期)2020-02-05
意林·全彩Color(2019年6期)2019-07-24
雪莲(2017年2期)2017-05-12
环球市场信息导报(2017年1期)2017-04-08
