
竞争性内源 RNAs 在骨关节炎发生机制中的调控作用
2022-07-22 07:30蓝奉军杨华刘淼吴鸿斌孙红
中国骨与关节杂志 2022年7期
蓝奉军 杨华 刘淼 吴鸿斌 孙红
骨关节炎 ( osteoarthritis,OA ) 又称退行性骨关节病,多见于中老年人群,好发于负重和活动量较多的关节。其多由年龄、肥胖、劳损、创伤和关节畸形等多因素引起关节软骨损伤退化、关节外缘和软骨下骨反应性增生。OA 发生后可导致关节疼痛、僵硬、畸形以及活动受限。虽然 OA 晚期可行人工关节置换,但关节假体寿命有限且功能恢复疗效仍不理想。因此,进一步探索 OA的致病因素、发病机制以及病理分子机制对早期防治 OA显得尤为重要。最新的研究表明,非编码 RNA 分子在基因调控中的研究有助于 OA 发病机制的探索。据报道,由长链非编码 ( long noncoding RNA,lncRNA )、环状 RNA( circle RNA,circRNA )、微小 RNA ( microRNA,miRNA )和信使 RNA ( messenger RNA,mRNA ) 等构建的竞争性内源 RNA ( competitive endogenous RNA,ceRNA ) 调控网络已广泛应用于 OA 发病机制研究及治疗靶点探索。笔者就ceRNAs 的生物学功能以及其在 OA 发生、发展中的分子机制作如下综述,以期进一步阐明 ceRNA 调控网络在 OA发病机制中的作用。
一、构建 ceRNA 网络相关 RNAs 分子
ceRNA 是一种全新的基因表达调控模式。相比其它调控网络,ceRNA 调控网络更精细和复杂,涉及更多 RNA分子,如 lncRNA、circRNA、miRNA 等。lncRNA 是一类长度超过 200nt 的非编码 RNA,且存在种类繁多的外显子转录,多数都不能被翻译成蛋白质而发挥作用。由于 lncRNA 在序列和空间结构上的不同、表达水平的不同以及结合蛋白的不同,其表现出的功能作用不同。根据不同的功能作用,可将其分为信号转导分子、诱饵分子、导向分子和支架分子四类。研究表明,lncRNA 不仅参与细胞发育和个体发育,还与疾病的发生发展密切相关。不仅如此,lncRNA 在表观遗传、细胞周期和细胞分化等生命活动中也发挥着重要调控作用。与此同时,通过研究 lncRNA 表观遗传学发现,其主要机制包括DNA 甲基化、基因组印记、基因沉默和 RNA 编辑等。circRNA 存在于绝大多数生物中,真核生物中最为丰富,其结构上既没有游离的 5’ 端和 3’ 端,也没有聚腺苷化结构,不易受外切酶的影响,较线性 RNA 更加稳定。而且 circRNA 中包含的 miRNA 结合位点丰富,在竞争性基因调控中发挥重要作用。与 miRNA 结合后可影响转录沉默、翻译和特异性的 mRNA 降解。通过对 circRNA 在基因表达调控研究的不断深入,其在疾病发生机制以及疾病修复过程中的作用较前更加清晰。miRNA 是一类由内源基因编码的单链非编码小分子 RNA,不仅可以有多个靶基因,而且多个 miRNA 也可以调节同一个基因。研究发现,miRNA 调控基因主要是通过抑制靶基因的翻译或阻断靶基因在转录后水平的基因表达。miRNA 及其相关靶基因在细胞内构建了全方位多层次的基因表达调控系统,参与细胞分裂、增殖和凋亡等,不仅能参与疾病的发生与发展,而且还参与疾病的修复过程。
二、lncRNA 作为 ceRNA 在 OA 发病机制中的作用
OA 是一种进行性退化的骨关节疾病,主要病理变化表现为关节软骨破坏和缺失、软骨下骨质增生、炎症反应、关节间隙变窄和细胞外基质代谢等。研究表明,软骨受损后软骨细胞会启动相应的修复系统,分泌修复蛋白修复受损组织。也不可否认,修复蛋白对组织稳态的维持以及细胞外基质代谢的调节发挥着重要作用。针对 OA 发病机制、病理表型以及修复过程的研究发现,应用基因网络探寻 OA 发病机制及修复过程越来越广泛。ceRNA 网络作为一种新兴的机制,在调节软骨细胞功能作用中的重要程度也将被更清晰地揭晓。
根据目前研究发现,lncRNA 无论是在个体发育、细胞发育,还是在疾病发病过程都发挥着重要作用。与此同时,lncRNA 在转录沉默、转录激活、染色体修饰以及核内运输等显示出的功能作用与多种疾病的发生有关。其中,OA 就是受 lncRNA 异常表现影响的多因素疾病。然而,对 lncRNA 在 OA 中的功能作用和表达方式的认知还微乎其微。近年来通过研究 OA 发现,lncRNA 在软骨细胞、细胞外基质、间充质细胞以及炎症反应发生发展过程中起着重要作用。
1.lncRNA 作为 ceRNA 在 OA 软骨细胞中的调控作用:越来越多的研究发现,lncRNAs 在不同组织表达量不同,而且在同一组织不同状态下表达也不相同。通过研究表明,lncRNAs 在 OA 发生后的软骨细胞中表达量及表达方式不同 ( 表1 )。Liu 等研究发现,在软骨细胞中 lncRNA 还原酶 ( methionine sulfoxide reductase,MSR )与 miR-152 竞争调控胸腺素 4 ( thymosin beta 4,TMSB4 )的表达,而 MSR 上调可引起软骨降解、导致 OA 的发生。Wang 等报道,lncRNA-移行细胞癌相关因子 1( urothelial cancer-associated 1,UCA1 ) 表达上调能促进基质金属蛋白酶 ( matrix metalloproteinase,MMP )-13 表达,抑制 miR-204-5p 表达,从而促使 OA 的发生。而且 lncRNA 分化拮抗非蛋白编码 RNA ( differentiation antagonizing non-protein coding RNA,DANCR ) 也可与mi-1275 竞争调节 MMP-13 表达,促进滑膜间充质干细胞增殖和软骨形成。Xu 等通过实验验证表明,lncRNA母体表达基因 3 ( maternally expressed gene 3,MEG3 ) 敲除或下调引起母体对抗同源生长因子 7 ( mothers against decapentaplegic homolog 7,SMAD7 ) 表达下降、miR-16 过表达,导致软骨细胞增殖并促进细胞凋亡,促使 OA 发生。而且 MEG3 可通过调节同源磷酸酶 ( phosphatase and tensin homnlog,PTEN ),抑制滑膜细胞增殖、促进细胞凋亡,促使骨关节炎的发生。Huang 等发现,lncRNA肿瘤坏死因子超级家族成员 10 ( tumor necrosis factor superfamily member 10,TNFSF10 ) 过表达通过调节 miR-376-3p / 成纤维细胞生长因子受体 1 ( fibroblast growth factor receptor 1,FGFR1 ) 轴引起软骨细胞增殖和炎症反应,同时抑制细胞凋亡,促进 OA 的发展。
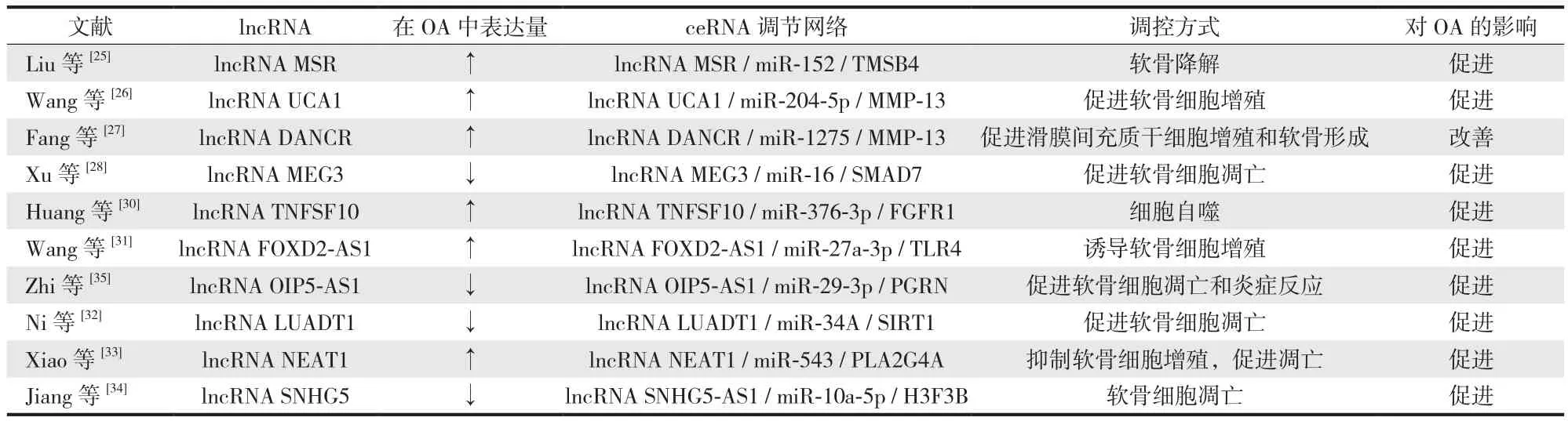
表1 lncRNA 作为 ceRNA 在软骨细胞中的调控作用Tab.1 The regulation role of lncRNA as ceRNA in chondrocytes
通过实验研究发现,lncRNA 叉头基因盒 D2 反义RNA1 ( forkhead box D2 adjacent opposite strand RNA 1,FOXD2-AS1 ) 与 OA 的严重程度成正比,主要通过调控miR-27a-3p / 跨模样受体 4 ( tol-like receptor 4,TLR4 ) 轴来发挥关键作用,而且 FOXD2-AS1 过表达可诱导软骨细胞增殖、炎症和细胞外基质 ( extracellular matrix,ECM ) 降解。通过研究发现 miR-34a 能促进软骨细胞凋亡,lncRNA肺腺癌基因 1 ( lung adenocarcinoma transcript 1,LUADT1 )过表达可特异性结合去乙酰化酶 1 ( sirtuin 1,SIRT1 ) 引起miR-34a 下调,故提高 LUADT1 的表达能有效地抑制软骨细胞凋亡,从而延缓 OA 的发展。Xiao 等研究 OA 发现,lncRNA 核富集转录物 1 ( nuclear paraspeckle assembly transcript 1,NEAT1 ) 在 OA 中高表达,通过调控 miR-543 /磷脂酶 A2 组 ⅣA 基因 ( phospholipase A2-ⅣA,PLA2G4A )轴,海绵化 miR-543 并诱导 PLA2G4A 的表达,从而抑制软骨细胞增殖、促进软骨细胞凋亡,诱发 OA 的发生发展。Jiang 等研究发现,lncRNA 小核仁 RNA 宿主基因 5 ( small nucleolar RNA host gene 5,SNHG5 ) 可通过调节 miR-10a-5p / H3 组蛋白家族成员 3B ( H3 histone family member 3B,H3F3B ) 轴促进软骨细胞增殖,抑制凋亡并延缓 OA 的发生。Zhi 等实验研究发现,白介素 1( interleukin 1,IL-1 ) 诱导 lncRNA 相互作用蛋白翻译RNA 1 ( opa-interacting protein 5 antisense RNA 1,OIP5-AS1 )下调,通过调控 miR-29b-3p / 颗粒蛋白前体 ( progranulin,PGRN ) 轴,抑制软骨细胞的增殖和迁移能力,促进凋亡和炎症反应,加重 OA 的发生。Lu 等通过研究发现,在 OA 患者中 lncRNA 软骨损害相关因子 ( cartilage injuryrelated,CIR ) 明显上调,同时伴有 miR-130a 下调和 Bcl-2蛋白家族促凋亡蛋白 ( Bim ) 上调,通过生物信息学分析发现 miR-130a 是 lncRNA-CIR 和 Bim 的靶点,若敲除lncRNA-CIR 基因 Bim 表达显著增加,然而 miR-130a 能明显抑制 Bim 的表达,升高活性氧、炎症介质的水平,增加软骨细胞凋亡,促进 OA 的发展。综上可知,lncRNAs 在OA 软骨细胞中发挥着重要的作用,调控软骨细胞的增殖或凋亡,为治疗 OA 提供了新的方向。
2.lncRNA 作为 ceRNA 在 OA 软骨细胞 ECM 中的作用:通过 lncRNAs 与 OA 研究发现,lncRNAs 通过促进软骨 ECM 降解或合成,可促进或抑制软骨破坏或损伤( 表2 )。Zhu 等研究发现,在 OA 中 lncRNA 脂肪酸合成酶反义 RNA 1 ( antisense transcript of Fas,FAS-AS1 ) 表达增加,而过表达的 FAS-AS1 可增加 MMP-1、MMP-13 的表达,降低 Ⅱ 型胶原蛋白 α1 ( collagen type Ⅱ alpha 1,COL2α1 ) 的表达,抑制细胞增殖,促进细胞凋亡,促进细胞外基质的降解。Chen 等的研究结果显示,lncRNA MEG3 过表达可促进 IL-1 诱导的软骨细胞增殖,抑制细胞凋亡并缓解 ECM 的降解,同时延缓 OA 的进程。Zhang等通过实验观察发现,OA 患者中 lncRNA 转移相关肺腺癌转录本 1 ( metastasis associated lung adenocarcinoma transcript 1,MALAT1 ) 和 AKT 丝氨酸 / 苏氨酸激酶 3( serine / threonine kinase 3,AKT3 ) 表达上调,而 miR-150-5p表达明显下降,提高了细胞增殖能力,抑制了细胞凋亡和 ECM 的降解,故 MALAT1 通过 miR-150-5p / AKT3 轴参与了 OA 的调控。而在 lncRNA MALAT1 / miR-145 / 含血小板反应蛋白基因的金属蛋白酶 5 ( a disintegrin and metalloproteinase with thrombospondin motifs 5,ADAMTS5 ) 信号通路的调节作用下 IL-1 可诱导软骨细胞 ECM 降解。Liu 等实验研究发现,lncRNA X 转录子 ( X-inactive specific transcript,XIST ) 在 OA 软骨组织中高表达,然而敲除软骨细胞中 XIST 发现,能明显增强软骨细胞活力,抑制 ECM 降解和细胞凋亡,抑制 OA 的发生。Tan等研究观察发现,软骨修复主要特点是增加细胞活力和迁移,同时降低 ECM 降解,而 lncRNA H19 通过调控miR-106-5p / 金属蛋白酶 2 组织抑制剂 ( tissue inhibitor of metalloproteinases 2,TIMP2 ) 轴可抑制 ECM 降解,同时促进 OA 软骨细胞增殖和迁移。因此,H19 将来有可能成为治疗 OA 潜在靶点。Chen 等通过实验研究发现,lncRNA 同源转录反义基因间 RNA ( hox transcript antisense RNA,HOTAIR ) 在小鼠关节软骨和 IL-1 诱导的软骨细胞中明显上调,过表达的 HOTAIR 通过调控 miR-20b / PTEN轴促进 ECM 降解、细胞增殖和凋亡,促进 OA 的发展。

表2 lncRNA 作为 ceRNA 在软骨 ECM 代谢中的作用Tab.2 The role of lncRNAs as ceRNAs in cartilage ECM metabolism
3.lncRNA 作为 ceRNA 在 OA 关节滑膜炎症中的作用:骨关节滑膜的炎症反应是引起 OA 的病因之一,而 lncRNAs在骨关节滑膜炎症反应中扮演重要的角色,故 lncRNA可通过调节滑膜炎症反应调节 OA ( 表3 )。Zhao 等研究发现,lncRNA 浆细胞瘤变异异位 1 ( plasmacytoma variant translocation 1,PVT1 ) 在 OA 的表达升高,若降低其表达可减轻炎症反应和骨代谢失衡,改善 OA 的进展。Pan 等报道,在小鼠软骨源性成软骨细胞 ATDC5 中lncRNA MALAT1 表达上调,其过表达可逆转脂多糖诱导的细胞炎症损伤,延缓炎性反应引起 OA 的进展。而 Liu等研究发现,肿瘤坏死因子和异质性胞核核糖蛋白 L 相关的免疫调节因子 ( tumor necrosis factor and heterogeneous nuclear ribonucleoprotein L-related immunoregulatory,THRIL )过表达能增强脂多糖诱导细胞炎性损伤。也有研究发现,免疫球蛋白重连基因 γ1 ( immunoglobulin heavy constant genes γ1,IGHCγ1 ) 在 OA 中表达上调,通过调控miR-6891-3-p / TLR4 轴调节炎症反应。同时 Yu 等发现 lncRNA 心肌肥厚相关因子 ( cardiac hypertrophy related factor,CHRF ) 在 OA 中过表达可使 miR-146a 表达下调,增强 IL-6 诱导炎症反应,加重细胞炎症损伤,促进 OA进展。

表3 lncRNA 作为 ceRNA 在滑膜炎症中的作用Tab.3 The role of lncRNA as ceRNA in synovial inflammation
三、circRNA 作为 ceRNA 在 OA 发病机制中的作用
circRNA 作为一类新型的 RNA 分子,具有高稳定性、组织特异性和作用广泛性等特点。它具有的 miRNA“海绵”功能,主要通过影响基因转录的调控与表达,参与 OA 发生发展过程。而且大部分 circRNAs 具有miRNA 结合位点,作为 ceRNA 可与 miRNAs 结合并隔离miRNAs。根据目前 circRNA 的研究表明,在作为海绵吸附、关节软骨的退变和修复方面起着重要作用。
越来越多的研究表明,circRNA 不仅参与骨形成过程,而且还参与骨关节疾病的发生过程( 表4 )。Li等研究发现,circ-0045714 可以通过改变 miR-193b 的靶基因表达水平调节 ECM 合成和软骨细胞的增殖和凋亡。也有研究发现,经白介素-1β ( interleukin-1 beta,IL-1β ) 刺激的软骨细胞,circ-0005105 表达明显上调,从而抑制 miR-26a 转录活性,上调其靶基因酰胺磷酸核糖转移酶 ( nicotinamide phosphoribosyltransferase,NAMPT )的表达,从而促进 ECM 降解诱发 OA。Li 等研究发现,circRNA-0136474 可竞争性结合 miR-127-5p,增强 MMP-13 表达,促使软骨细胞凋亡,加重 OA 的进展。而且 circRNA-33186 也可竞争性结合 miR-127-5p 参与 OA的发病机制调控。Zhou 等也报道,OA 软组织中的钙黏蛋白 13 ( cadherin 13,CDH13 ) 上调,竞争性结合miR-296-3p,可显著诱导软骨细胞凋亡,促进 ECM 降解、抑制 ECM 合成,促进 OA 的发展。Bai 等也证实,环状RNA 跨膜蛋白抑制剂模体 6 ( transmembrane BAX inhibitor motif 6,TMBIM6 ) 可通过调节 miR-27a / MMP-13 轴促进ECM 降解。与此同时 Shen 等发现,环状 RNA 周期性蛋白依赖激素 14 ( cyclin dependent kinase 14,CDK14 ) 对关节磨损具有保护作用,主要通过 miR-125a-5p 介导,下调类似于母系反义同源序列 2 ( Smad2 ),引起转化生长因子 β信号通路障碍,从而调节软骨代谢,抑制细胞凋亡,延缓OA 的发生。综上所述,circRNA 参与了 OA 的病理发生发展过程,可作为该疾病诊断的生物标志物。

表4 circRNA 作为 ceRNA 在 OA 中的作用Tab.4 The role of circRNAs as ceRNAs in osteoarthritis
四、小结与展望
ceRNA 网络调控机制是近期发现的体内 RNA 之间相互作用、相互影响,并且可调控基因的新机制。近年来,ceRNA 网络调控机制已成为研究的热点,其在疾病发生发展过程中发挥了重要的作用。OA 目前仍是临床上最常见的退行性关节疾病,基因诊断和治疗是目前研究的热点,相信这也是未来的诊疗趋势。ceRNA 网络调控机制将为探究 OA 发病机制和治疗靶点提供新思路和方向。ceRNA 调控网络与 OA 发生、发展过程以及预后密切相关,并且有可能成为 OA 早期诊断、治疗和预后评估的靶点。然而,ceRNA 在软骨细胞、细胞外基质、炎症反应和 OA 的发病机制的研究尚处于初始阶段,需要进一步的深入研究。
猜你喜欢
中国骨与关节杂志(2022年10期)2022-10-20
中学生数理化·高一版(2021年12期)2021-09-05
现代农业科技(2018年13期)2018-10-20
祝您健康(2018年10期)2018-10-11
现代养生·上半月(2016年9期)2016-05-14
中国信息化周报(2014年4期)2014-02-17
