
两种互逆资源梯度影响下刺叶锦鸡儿的种子选择性败育格局
2022-08-03 02:53王晓伟韩佳玲张爱勤
生态学报 2022年12期
王晓伟,韩佳玲,张爱勤,*
1 新疆大学生命科学与技术学院,乌鲁木齐 830017 2 新疆生物资源基因工程重点实验室,乌鲁木齐 830017
在不确定的传粉和母本环境中,大多数有花植物常出现“花多果少”或“胚珠多种子少”的现象[1—2],这一现象通常与植物的选择性败育有关。选择性败育是指在那些生殖资源受限的植物上或者以提高后代适合度为选择力时,植物有选择地败育部分发育中的幼果或种子的现象[3]。其败育方式具明显的位置效应。如在豆科植物中,根据荚果内种子败育位置的不同可分为不同的类型[4]:(1)成熟荚果顶部和中部的胚珠,选择性败育基部的胚珠[5—6];(2)成熟荚果中部胚珠,而败育荚果顶部和基部的胚珠[7—8];(3)仅在顶部、中部或基部形成一粒种子,其余胚珠均败育[9];(4)为交替性败育类型,即在荚果偶数或奇数位置的胚珠败育等[10]。
影响植物选择性败育的因素有多种,如源-库关系;分配给果实/种子的有效营养成分和光合产物的量;果实和种子分化的时间和位置;花粉来源和数目;病虫害;各种环境因素等[4—5,11—12]。其中,资源的运输方向、分配模式及植物的传粉受精过程等是种子形成的重要影响因素被广泛关注[4,7,13—14]。如在一些豆科植物的研究中显示近柱头端的胚珠因为优先接触到花粉管,在受精时间上具优势,表现为顶端胚珠的败育率低[15—16];而位于基部的胚珠因为更接近营养物质、水和光合产物的源头,在资源竞争中具位置优势,出现基部胚珠的低败育率[17—18]。另一些研究则发现子房内柱头端第一位的胚珠并不总是由生长最快的花粉管受精,第一个花粉管或生长最快的花粉管会倾向于子房中特定位置的胚珠受精[7]。对十字花科葱芥(Alliariapetiolata)的调查显示,由于传粉环境差异,不同分布区域的种群种子形成模式不同,存在角果内中部胚珠发育良好和基部胚珠发育良好两种类型[18],这些现象都反映了两种资源作用方式的复杂性和结籽格局的多样性。
胚珠的发育既要经历传粉不足所导致的受精前败育风险,还要经历胚珠间资源竞争所导致的受精后败育风险。在豆科/十字花科植物的线性荚果/角果中,花粉管生长方向和资源运输方向是两个互逆的资源梯度。同样受两种资源梯度的影响,为什么有的果实选择性败育顶端胚珠,而有的果实却选择性败育基部或中间位置的胚珠? 要回答这一问题,不仅需要对开花后胚珠发育动态和败育过程进行追踪,以了解两种互逆资源梯度的作用方式及作用过程,还需要在不同选择性败育类型的植物中开展传粉和资源分配状况的调查,探究两种资源产生作用的条件。而已有研究更多处于现象的解释和多种假说并存的状态[19],对两种资源梯度的作用过程及方式关注不足[4,12,20]。
刺叶锦鸡儿(Caraganaacanthophylla)是豆科锦鸡儿属的一种多年生灌木,是干旱区植物区系组成的一个重要类群。因其根系发达、具固氮、抗逆性强等特点,成为防风固沙和水土保持的重要植物资源。同时,因其花色鲜艳、株丛高大且叶量较多又是早春的景观植物和主要牧草之一。前期调查显示,该植物结荚率很高,但荚果内结籽率较低,胚珠的形成具明显的位置效应,其结籽格局受两种资源梯度如何作用所致并不清楚。为此,从传粉和资源分配两个方面着手,在对植物的传粉生态、交配系统进行研究的基础上,通过资源调控、种子形成及败育过程的跟踪调查,探讨两种互逆的资源梯度如何影响胚珠的发育,为豆科植物种子选择性败育的深入研究及刺叶锦鸡儿种子产量和种群保护的研究积累新的资料。
1 材料与方法
1.1 研究材料与观测地点
刺叶锦鸡儿是豆科锦鸡儿属的一种多年生灌木。植株基部多分枝,托叶和叶轴在长枝上则硬化成针刺。花冠黄色,旗瓣宽卵形,二体雄蕊。生于干山坡、山前平原、河谷、沙地,常成片生长,花期4—5月。实验地点位于乌鲁木齐市近郊干旱荒漠化的山坡上(43°44′7.52″N,87°37′53.85″E;海拔 900—950 m)。年平均温度5—15℃,年降雨量199.7mm。刺叶锦鸡儿为群落的优势种,高0.7—1.5m,覆盖度为60%—70%。伴生种主要为百合科(Liliaceae)、禾本科(Poaceae)和豆科(Fabaceae)的一些植物,处于零星分布状态。可能由于生境的异质性,所调查刺叶锦鸡儿居群在开花期呈明显的斑块状分布,表现为极少开花的植株斑块和正常开花的植株斑块两类。前者比例较低,多分布在地势较高的砾石质荒坡上,其植株上开放花的花部特征、开花动态与正常植株上的花无任何差异,也无个体大小依赖现象。正常开花的植株斑块仍是居群的主要组分,下述实验均以正常开花植株为研究对象。
1.2 传粉特性
1.2.1花展示与花设计
(1)开花习性与开花动态:盛花期,随机选取观测样地正常开花的15个植株,每株标记2—3朵待开放的花,分别记录一天中花的开放时间、开放后花的持续时间和闭合时间等。同时,对居群花期的持续时间、植株间开花的同步性、单株开花后的持续时间、开花强度等进行记录。
(2)花部特征参数:盛花期,随机标记15个正常开花的植株,每株标记2朵待开放的花,在花开放后取下,进行雌雄蕊高度、旗瓣长度、翼瓣长度、龙骨瓣长度的测定(测量工具为游标卡尺,分辨率为0.01mm)。
(3)单花花粉量与胚珠数的测定:参考Cruden的方法[21]。盛花期,随机标记15个正常开花的植株,每株取1朵待开放且未散粉的花进行以下操作:(1)将单花的10枚花药小心取下放入EP管中,同时对其子房中的胚珠数进行统计;(2)每个EP管加入100μL 20%蔗糖浓度的番红溶液制成悬浊液,从中取0.5μL液体滴在载玻片上,进行液滴中花粉粒数目的统计,后用统计数据计算单花花粉量和花粉胚珠比(P/O)值;单花花粉量=统计出的液滴中花粉粒数目×100/0.5;P/O=单花花粉量/胚珠数。
1.2.2传粉者访花动态及访花频率
盛花期,选取10株正开花的植株,每株随机标记10—20朵当日开放的花,从早上9:00至下午18:00进行连续观察,记录昆虫访花行为和标记花的被访次数,后计算访花频率,并拍照记录昆虫的种类。另外,随机选取15个开花植株,每株标记2朵未散粉的花进行去雄处理,并在当天统计去雄后花的柱头花粉数,检测传粉强度。
1.3 交配系统
盛花期,随机选取30个正常开花的植株,每株用细线标记12朵未散粉的待开的花,进行以下5种处理,每种处理2朵花,另外2朵不做任何处理作对照,待荚果成熟后统计结实率,5种处理如下:
(1)孤雌生殖:对未散粉的花进行去雄处理并套袋;
(2)人工自花授粉:对标记的花进行套袋,待花开放后去掉袋子,用镊子轻轻夹龙骨瓣使其弹开,让柱头和花药充分接触后再套袋。每操作完成一朵花后,用湿纸巾将镊子擦拭干净后再继续下一朵;
(3)去雄人工异花授粉:对未散粉的花进行去雄处理后套袋,待花开放后去掉袋子进行人工授异花粉,后继续套袋;
(4)自动自花授粉:对标记的未散粉的花套袋不做任何处理;
(5)去雄不套袋:对标记的未散粉的花进行去雄处理后使其自然授粉。
1.4 胚珠败育与种子的形成动态
在盛花期,随机标记120个正常开花的植株,每株标记10朵待开放的花(累计1200朵),待花开放后根据胚珠发育状况,将胚珠发育过程分为开花后5d、8d、11d、14d、17d、20d 共6个阶段。每个阶段从120株中随机选20株,取其上边标记的花或幼果(样本量:200个/每阶段)进行豆荚中每个位置胚珠发育状态的记录,把荚果基部第一个胚珠设为1号位,依次对各位置的胚珠进行编号,到顶部最后1个胚珠为17号位。同时,根据胚珠发育状况,将胚珠分为3种类型,分别是:败育胚珠、正常大小胚珠(与受精前成熟胚珠大小一致,且有活力)和膨大胚珠(受精后已经发育的胚珠,体积大于正常大小胚珠)(图1C)。后根据统计结果进行胚珠发育动态的描述。
1.5 资源调控下的结籽格局
在花期,随机选择4个极少开花的斑块,每斑块标记5株(共20株)有花开放的植株,分别在末花期和结荚初期各进行一次水肥处理(方法:在4个斑块内布滴灌喷头,先在植株的根部施加磷钾肥,后滴灌4h)。同时,另选4个极少开花的斑块,每斑块内标记5株(共20株)有花开放的植株,不进行任何处理做对照。荚果成熟后,对每标记株随机取8个荚果,分别统计荚果中败育种子和成熟种子的位置及总的种子数,比较资源增加前后植株的结籽率及结籽格局。
1.6 数据分析
用SPSS 20.0和Excel 2010软件进行数据的处理、分析与图表绘制,其中用单因素方差分析(One-Way ANOVA)比较花大小参数;用独立样本t检验分析资源调控下植株的结籽率;用广义线性模型(Generalize Linear Model)比较交配系统检测中不同处理的结实率、结籽率。所有统计数据用平均值±标准误表示。
2 结果与分析
2.1 传粉特性
2.1.1花展示与花设计
开花习性与开花动态:刺叶锦鸡儿为早春开花植物,4月中旬进入花期,可持续到5月上旬。在花期,所调查居群的开花呈斑块状分布,表现为大多数植株处于大量的同步开花状态(图1A),而少数植株上没有花芽或花芽极少,处于不开花或少量开花的状态。居群中,单花开放集中在早上9:00左右,开放后的花可持续4—5d,雄蕊在开放前1天开始散粉。不同类型植株上开放的花具有同样的开放特点和花寿命。
花部特征参数:刺叶锦鸡儿为蝶形花,花冠由旗瓣、翼瓣和龙骨瓣组成(图1B)。龙骨瓣两瓣合生,翼瓣着生于龙骨瓣两侧,旗瓣包裹着翼瓣和龙骨瓣基部。雌、雄蕊的花柱、花丝顶端形成一定弧度的弯曲,离生雄蕊最短,其余长短雄蕊交错排列,形成上下两轮,雌蕊被围在雄蕊之间(图1B)。高位雄蕊(25.93±0.29)mm显著长于低位雄蕊(24.45±0.36)mm(F=10.280,P=0.002)。旗瓣(27.89±0.45)mm和翼瓣(27.33±0.63)mm同长,均长于龙骨瓣(F=5.208,P=0.009)(表1)。

表1 刺叶锦鸡儿的花大小参数Table 1 Floral size parameters in Caragana acanthophylla
单花花粉量与胚珠数:刺叶锦鸡儿的单花花粉量为(247400±27450)粒,胚珠数为(14.00±0.14)粒。P/O值为(41122.55±5811.23),推断为专性异交的交配系统。
2.1.2传粉者访花动态及访花频率
传粉者类群及访花行为:刺叶锦鸡儿的传粉者种类多样,为泛化的传粉系统。主要的传粉者有无垫蜂属、条蜂属的一些体型较大或中等的种(图1D、图1E),均以花蜜为食。体型稍大的传粉者访花时,可以通过自身的重量将龙骨瓣弹开,完成花粉的传递;但体型小的传粉者(图1F)在访花时很难将龙骨瓣弹开,无法进行花粉的传递,为无效访花者。

图1 刺叶锦鸡儿植株、花结构、不同阶段的胚珠及主要传粉者Fig.1 Plant, anatomic structure of flower, ovule at different stages and main pollinators of Caragana acanthophyllaA:植株;B:花解剖图;C:不同阶段的胚珠;D—E:体型较大的蜂(条蜂和无垫蜂属的种);F:体型较小的蜂(彩带蜂亚科的种)
访花动态及花粉传递效率:在晴朗天气,传粉者一般在早上9:00左右开始出现,访花频率为0.0857次数花-1h-1;在13:30—15:00,访花昆虫活动频繁,访花频率达0.2926次数花-1h-1;17:00之后昆虫数量开始减少,访花频率(0.0196次数花-1h-1)明显下降;18:00时基本不再有访花者(图2)。在开花当天,去雄后的柱头花粉数达(12.11±5.87)粒,说明柱头有较充足的异花粉。
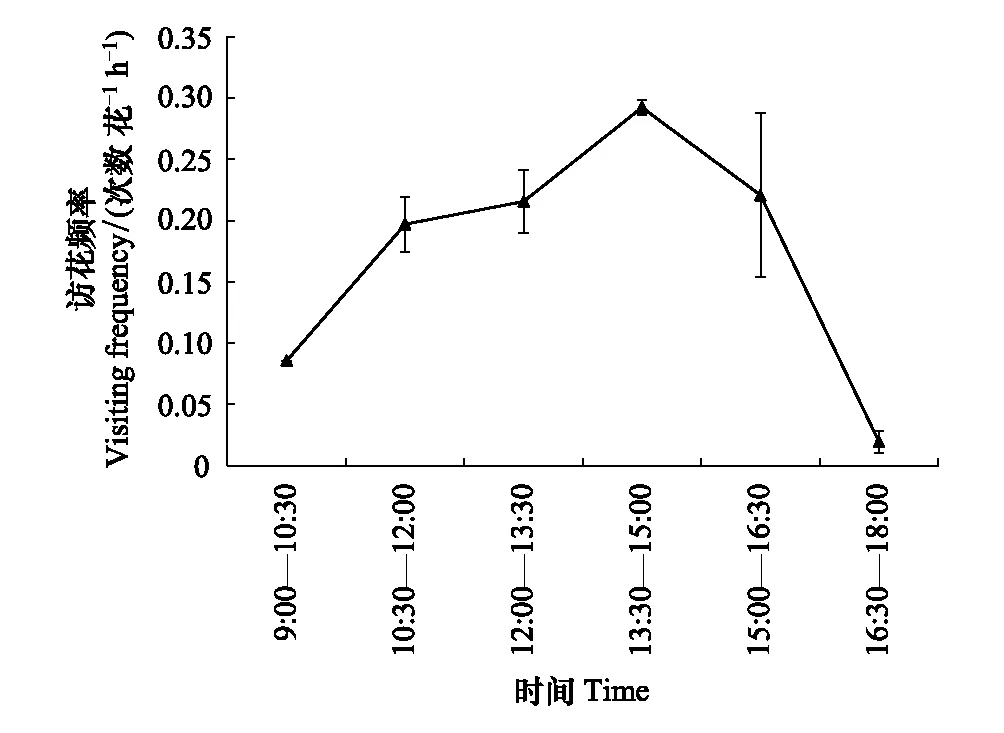
图2 刺叶锦鸡儿主要传粉者的访花动态 Fig.2 The visiting frequency of main pollinators in Caragana acanthophylla
2.2 交配系统
人工控制授粉实验显示不同的授粉处理具不同的结实状况(Waldχ2=789.662,P<0.01)(图3)。其中,孤雌生殖、自动自花授粉和人工自花授粉的结实率均为0,说明刺叶锦鸡儿无孤雌生殖现象,具严格的自交不亲和性;而去雄自然授粉、人工补充授异花粉和自然状态的结实率分别为(82.00±5.49)%、(84.00±5.24)%、(86.00±4.96)%,三种处理无显著差异(Waldχ2=0.298,P=0.861),均具较高的结实率,说明不存在传粉限制,其交配方式与P/O的检测结果一致,为专性异交的交配系统类型。

图3 刺叶锦鸡儿不同授粉处理下的结实率与结籽率Fig.3 The fruit setting rates and seed setting rates under different treatments in Caragana acanthophylla不同大写字母表示结实率的差异显著(P<0.05),不同小写字母表示结籽率的差异显著(P<0.05)
对不同授粉方式下所结荚果进行结籽率统计显示,各处理的结籽数都较低,无显著差异(Waldχ2=8.906,P=0.113)。其中自动自花授粉、人工自花授粉和孤雌生殖均为0;人工补充授异花粉、去雄自然授粉和自然状态的结籽率分别为(14.29±7.82)%、(13.04±7.18)%、(15.00±8.19)%,三者也无显著差异(Waldχ2=0.035,P=0.983)。
2.3 胚珠败育及种子形成动态
对不同发育阶段的荚果进行种子发育状况的调查显示:刺叶锦鸡儿胚珠的发育从顶部开始,多数胚珠能受精膨大,但最终在中部形成2—3粒种子,胚珠的败育从基部开始(图4)。其胚珠败育与种子形成动态如下(图5):在开花后的第3天,子房内开始出现胚珠膨大现象,主要集中在顶部。第5天,膨大胚珠的比例进一步增加,此时,绝大多数子房中只有膨大胚珠和不膨大胚珠两种类型。在开花后的第8天,子房基部的膨大胚珠比例有所上升,中部的8—13号位所占比例最高(58.25%—68.82%)。此时,个别子房基部开始出现胚珠败育现象,败育比例仅有0.52%。在开花后的第11天,顶部和中部的胚珠膨大比例继续增加,出现大多数胚珠的膨大,子房基部的胚珠败育比例有所增加,在1号位所占比例最大(10.20%)。在开花后的第14天,顶部和基部胚珠败育显著增加,在基部1号位达到最高峰(44.76%);胚珠膨大比例在子房中呈现正态分布,并在11号位达到最高(48.25%)。与第11天相比,膨大胚珠的比例有所下降,顶部胚珠出现明显的膨大后败育现象。在开花后的第17天,子房内两端胚珠继续败育,1号位的败育比例继续上升(60.41%)。在第20天,子房内两端胚珠的败育率继续增加,中部胚珠的发育相对稳定。在1号位败育率达到最高值(82.00%),中部11号位最低(30.00%),种子形成格局基本稳定。
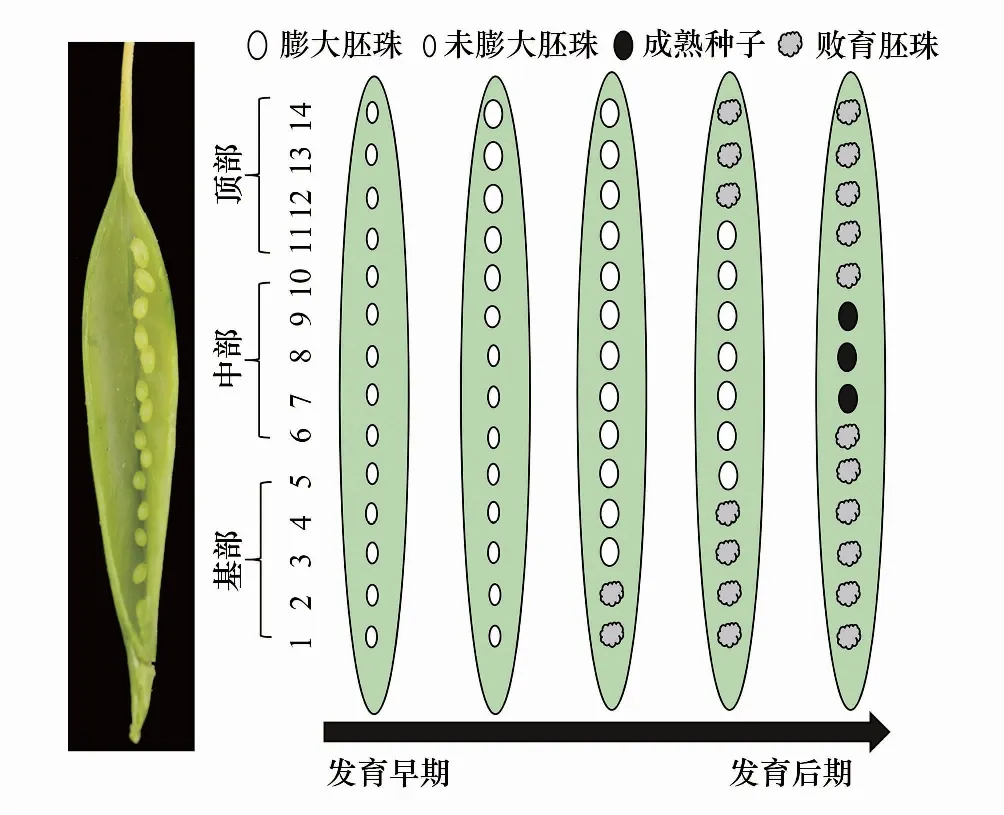
图4 刺叶锦鸡儿豆荚内不同时期胚珠的发育动态示意图 Fig.4 A diagram of legume pods showing ovule development at different positions in Caragana acanthophylla
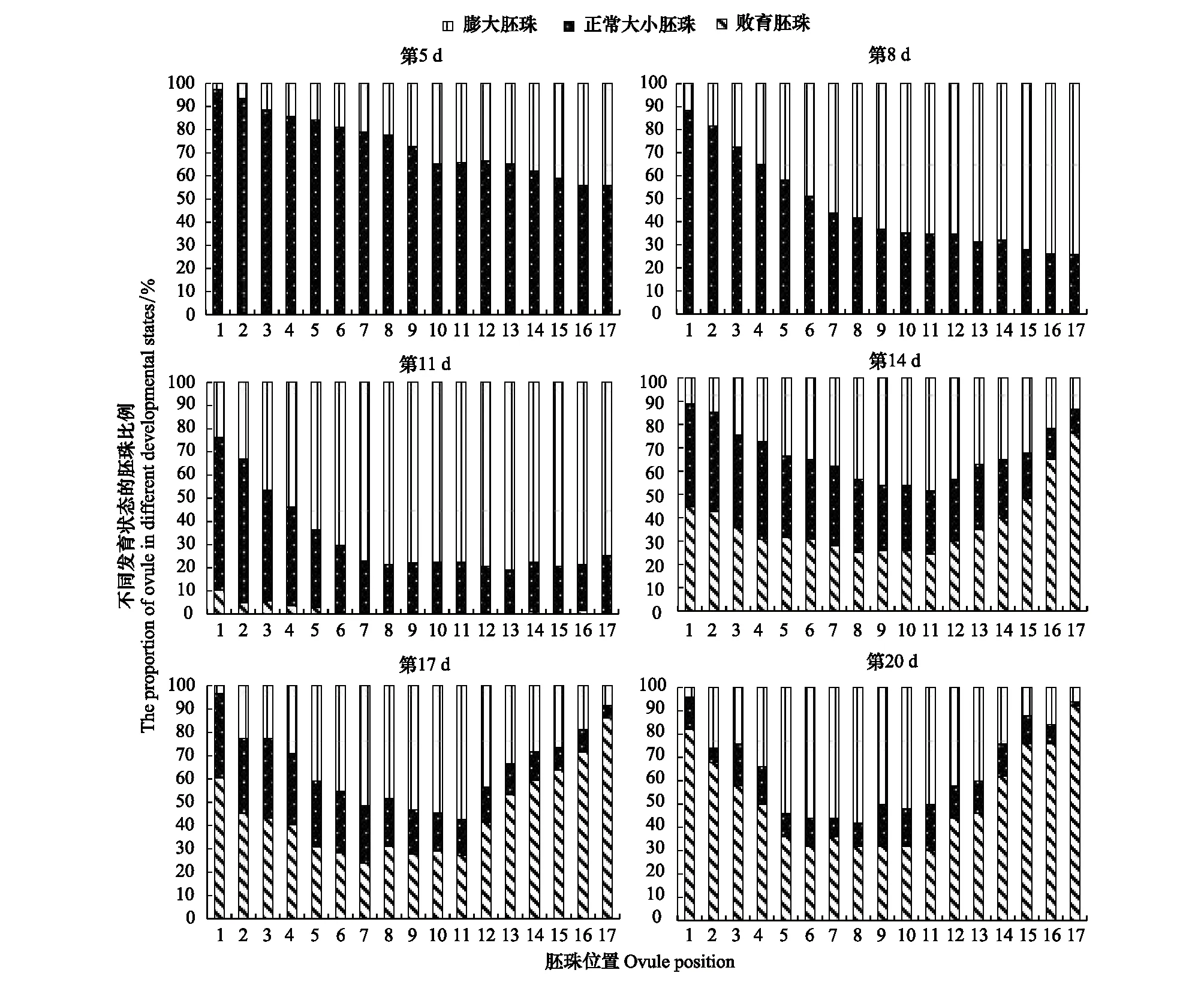
图5 刺叶锦鸡儿胚珠败育及种子形成的时间和位置效应Fig.5 The timing and location effects of ovule abortion and seed formation in Caragana acanthophylla基部从1号位开始,17号位为最顶部
2.4 资源调控下的结籽格局
在花期和结荚初期对极少开花植株添加水肥后(处理),荚果的结籽率(18.45±1.70)%较对照株(11.19±0.80)%提高了64.9%(F=4.468,P=0.001)(图6)。但资源调控并没有改变荚果的结籽格局,两类荚果仍然表现为中间位置高、两端低的结籽状况,与图4和图5所表现的胚珠发育动态一致。
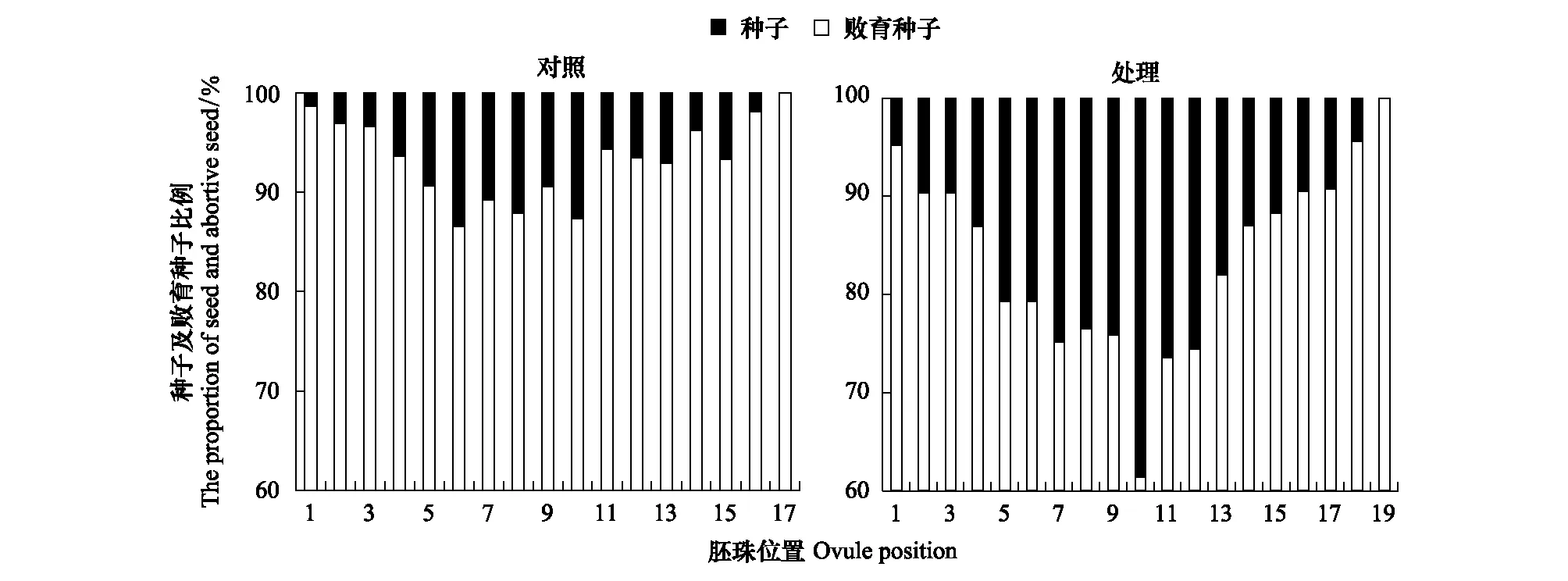
图6 刺叶锦鸡儿资源调控下植株的结籽格局Fig.6 The seed set pattern under resources regulation in Caragana acanthophylla对照:极少开花植株结籽格局; 处理:资源调控后植株结籽格局
3 讨论
传粉与资源限制是影响植物种子形成的两个重要因素,在豆科或十字花科植物中常常成为互逆的两个资源梯度。本文通过对刺叶锦鸡儿传粉特性、交配系统、胚珠发育动态及资源调控下植株结籽格局的调查,发现刺叶锦鸡儿为专性异交的交配方式,具充足的传粉者,不存在传粉限制,但资源限制及胚珠受精的不同步性使其发育具有明显的时间和位置效应。
3.1 种子形成的时间和位置效应
传粉与资源限制对种子发育的影响有多种途径,如传粉强度、柱头上花粉来源与数目、子房内胚珠位置、受精时间、果实在花序和植株上的位置等[4—5,11—12]。这些因素大多跟生殖单位受精的时间和所处位置有关。表现为花序或果实水平上,早发育的或近轴端(资源库)的生殖单位在资源竞争方面具更强的时间和位置优势[22—24],即种子或果实形成的时间和位置效应。刺叶锦鸡儿的结籽格局也反映这一现象(图4),即胚珠发育最先从子房的顶部开始,而败育从基部开始,最终形成两端结籽率低、中部高的种子选择性败育类型。此类结籽格局在荆豆属植物(Ulexgallii)[7]、十字花科芸薹属甘蓝(Brassicaoleracea)[8]和野豌豆属歪头菜(Viciaunijuga)[17]中也存在。
关于结籽格局,不少豆科植物显示近柱头端的胚珠因为在受精时间上具优势,表现出顶端胚珠结籽率高的现象[15—16]。而刺叶锦鸡儿等虽然顶端胚珠优先发育,但并没有在顶端形成较高的结籽率,而是在中部。究其原因可能与胚珠间较长的受精间隔有关。我们对同为豆科植物,授粉后基部胚珠优先发育,最终在基部形成1—2个种子的黄花草木樨(Melilotusofficinalis)进行分析发现[25]:黄花草木樨授粉不存在传粉限制,花开放后当天闭合,这意味着子房内顶端和基部胚珠间的授精间隔较短,在顶端胚珠受精时间没有优势的情况下,资源分配的位置效应可能会对胚珠的发育起主导作用。基于这一原因,那些顶端胚珠发育较好的植物可能是与传粉受限,中下部胚珠不能被正常受精有关。
而刺叶锦鸡儿花期4—5d,尽管不存在传粉限制,绝大多数胚珠都能受精,但在开花当天,去雄后的柱头花粉数(12.11±5.87)粒并不能满足所有胚珠(14.00±0.14)粒的受精需求。晴朗天气自然状态柱头花粉数在第3天才达到最大值[25],所以,柱头花粉的沉积是一个不断累积的过程。这就意味着基部与顶部胚珠有一个较长的受精间隔。在资源受限的情况下,晚受精的胚珠在争夺资源方面会处于劣势;而顶部的胚珠因为受精早得到优先发育。但随着胚珠体积的增长,资源竞争的加剧,顶部胚珠在资源竞争中的劣势将逐步凸显。只有中部胚珠在受精时间上与顶端较为接近,而在资源竞争中具明显的位置优势,从而得到较好发育。这一推理在两种栽培玉米的控制授粉实验中得到了较好的验证[26]。这可能也是为什么同样受两种资源梯度的影响却表现出不同结籽格局的原因。所以,探究受精后影响胚珠发育的因素,除了考虑资源分配状况及胚珠的位置影响外,还应该考虑前期胚珠间受精时间的长短及花粉限制的影响。
受精是胚珠可持续发育的前提,而受精后胚珠的发育对资源的需求是一个逐步增加的过程。所以,选择性种子败育通常表现为花粉限制多作用于果实、种子形成的早期,而资源竞争在后期尤为关键[12]。在这种作用模式下,如果植物不存在传粉限制,而存在资源限制,就会表现为在结实早期大量胚珠膨大,而后又败育的现象,这在很多果树中比较明显,刺叶锦鸡儿的这一结籽格局也印证了上述现象。
3.2 花粉传递与种子形成
传粉是植物完成受精过程的必备阶段,胚珠的败育往往与柱头上花粉的来源和数目有关。在许多植物中,要产生1枚果实,柱头上的最少花粉粒数要达到一定的数目,即传粉强度[5, 27—28]。花粉源也影响种子的发育,在自交不亲和的植物中,自花粉的落置往往会影响异花粉的萌发,从而导致胚珠受精不足而发生败育[7,14—15]。刺叶锦鸡儿的单花花粉量较大,属于花粉释放中的瓣膜机制。这种机制以具有弹性的花结构为基础,通过传粉者对花瓣的作用力,将花粉多次呈现出来,可以有效的避免花粉浪费,提高雄性适合度。从刺叶锦鸡儿的花粉传递看,尽管昆虫的访花会导致部分自花粉的滞留,但自交不亲和性并没有使其自然结实率(86.00±4.96)%受到影响。在交配系统的检测中,去雄自然授粉和去雄人工异花授粉在结实率和结籽率方面与自然状态的均没有差异,说明刺叶锦鸡儿具较高的传粉强度,不存在传粉限制。而在去雄套袋(孤雌生殖)和人工自花授粉的处理中,结实率和结籽率均为零,没有发现荚果内胚珠有膨大后又败育的现象,说明该植物在胚珠发育过程中不存在假受精现象。
传粉只是受精过程的第一步,花粉在柱头上的萌发状况及花粉管生长的先后顺序对胚珠的发育至关重要。从刺叶锦鸡儿胚珠的发育动态可以看出,受精的先后顺序,对胚珠的发育有重要影响。也体现了在胚珠发育早期,传粉、受精过程中时间效应的主导地位。
3.3 资源分配与种子形成
种子的形成与资源的可获得性密切相关。在资源有限的情况下,投入到某一性状或部位的资源量的增加必然会降低它投入到其他性状或部位的资源量,即资源分配的“此长彼消”[12, 17],如居群水平或植株水平的间歇性大量结实、逆境胁迫下繁殖方式或繁殖力的改变等。刺叶锦鸡儿居群斑块状的开花现象体现了繁殖资源分配的差异性,尽管还不能确定导致这种差异性的原因,但通过资源调控后,显著增加了低开花植株荚果的结籽率,说明存在资源限制。
所调查居群位于年均降雨量不到200mm的干旱荒漠化的低山坡上,多样化的地形和早春较低的降水可能会成为植株营养生长的一个重要影响因素,斑块状的开花可能与植株所处的微地形有关。这一现象说明资源分配与权衡在居群水平、个体植株的年份间及不同生长阶段、荚果内胚珠间等广泛存在,并对繁殖产生影响。这反映了母本在资源分配与调控形式方面的多样性。这种母本通过败育部分花或胚珠的资源分配方式在一定程度上对提高母本的繁殖适合度或种子的后代适合度均具重要意义[29—30]。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
种子(2022年5期)2022-06-27
延边大学学报(自然科学版)(2021年1期)2021-04-27
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
课外生活(小学1-3年级)(2020年2期)2020-03-09
农民致富之友(2018年8期)2018-06-27
大科技·百科新说(2018年3期)2018-04-11
中国绿色画报(2016年9期)2017-05-31
作品(2015年5期)2015-11-18
中国蜂业(2015年1期)2015-01-24
