
LncRNA NONSUST004358.1 在猪脂肪细胞分化中的作用研究
2022-09-14 02:36吴小敏张双翔朱二鹏许厚强赵佳福程振涛代兴红陈秀华袁超
中国畜牧杂志 2022年9期
吴小敏,张双翔*,朱二鹏,许厚强,赵佳福,程振涛,代兴红,陈秀华,袁超
(1.贵州省草地技术试验推广站,贵州贵阳 550025;2.贵州大学动物科学学院,贵州贵阳 550025;3.贵州省农业科学院畜牧兽医研究所,贵州贵阳 550002)
脂肪细胞起源于胚胎中胚层,由前体脂肪细胞分化形成,过氧化物酶体增殖物激活受体(Peroxisome Proliferator-Activated Receptor,PPAR)、脂肪酸合酶(Fatty Acid Synthase,FAS)和激素敏感性脂肪酶(Hormone-Sensitive Lipase,HSL)是其分化与沉积中最重要的转录因子和关键酶基因。近年来一系列研究发现,长链非编码RNA(Long noncoding RNA,lncRNA)在脂肪沉积中发挥重要的调控作用。大部分lncRNA 由RNA 聚合酶II 转录,具有与mRNA 相似的5帽子结构和3polyA 尾巴,在胚胎发育、脂肪沉积和肌肉生长中发挥重要调控作用。Liu 等通过lncRNA 微阵列分析发现,lncRNA Gm15290 能促进脂质沉积增加,调控脂肪形成基因和等表达。过表达lncRNA AK043773 可显著抑制小鼠骨髓基质细胞中脂肪细胞分化和脂滴的形成。本课题组前期研究发现,AMP 活化蛋白激酶1(Protein Kinase,AMP Activated Alpha 1,)是影响从江香猪肉品质的重要候选基因。根据PRKAA1 筛选的lncRNA NONSUST004358.1 在从江香猪皮下脂肪和背最长肌组织中有较高表达,提示lncRNA NONSUST004358.1可能在从江香猪脂肪沉积中发挥重要调控作用。本实验通过分离培养从江香猪皮下和肌内前体脂肪细胞,分析lncRNA NONSUST004358.1 在前体脂肪细胞分化过程中的表达规律,构建pEGFP-C1-lncRNA NONSUST004358.1 过表达载体,分析转染后前体脂肪细胞中PRKAA1、FAS、HSL、PPAR的表达变化,以及对前体脂肪细胞中甘油三酯和胆固醇含量的影响,探究lncRNA NONSUST004358.1 在前体脂肪细胞分化中的作用。
1 材料与方法
1.1 组织来源 5日龄从江香猪背最长肌和皮下脂肪组织。
1.2 实验材料 T4 DNA 连接酶、限制性内切酶I、I 和逆转录试剂盒为美国Thermo Fisher 公司的产品;胶回收试剂盒、质粒提取试剂盒和2×Taq PCR Master Mix 为美国AxyGen 公司产品;0.1% I 型胶原酶、细胞培养瓶、细胞培养板、巴氏吸管为无锡耐斯生物科技有限公司产品;含0.25% EDTA 的胰蛋白酶为赛澳美细胞技术(北京)有限公司产品;胰岛素、地塞米松、3-异丁基-1-甲基黄嘌呤为Sigma 公司产品;胎牛血清、基础培养基和Opti-MEM 培养基为美国Gibco 公司产品;FuGENEHD Transfection Reagent 为Promega公司产品;细胞RNA 提取试剂盒为美国OMEGA 公司产品;SsoFast EvaGreen Supermix 为宝生物工程(大连)有限公司产品;油红O 染色试剂盒为北京索莱宝科技有限公司产品;甘油三酯和胆固醇检测试剂盒为南京建成生物工程研究所产品。
1.3 实验方法
1.3.1 细胞的分离培养与鉴定 参照屈长青等研究方法,在无菌环境下分离从江香猪背最长肌和皮下脂肪组织,剪掉血管与筋膜后剪成1~2 mm,利用0.1% 的I型胶原酶在37℃条件对其进行消化,皮下脂肪和背最长肌组织分别消化1 h 和2 h,将消化混合液分别经200目、400 目细胞筛进行过滤,滤液在1 500 ×g 条件下离心5 min,弃上清,沉淀用DMEM/F-12(不含胎牛血清)培养基吹打清洗,重复3 次,最后用含10% 胎牛血清的DMEM/F-12 完全培养基重悬细胞沉淀并接种于细胞培养瓶内,运用差速贴壁法(4 h)对肌内前体脂肪细胞进一步纯化,皮下前体脂肪细胞在24 h 后进行换液。在皮下和肌内前体脂肪细胞培生长达到抑制后,加入含10% FBS+1 μmol/L 地塞米松+5 μg/mL 胰岛素+
0.5 mmol/L 3-异丁基-1 甲基黄嘌呤的脂肪细胞分化诱导培养基,培养48 h 后,换成含10% FBS+1 μg/m L胰岛素的脂肪细胞分化维持培养基,2 d 更换培养液1次,直到细胞脂滴达到80%时按照油红O 染色说明书对诱导分化的细胞进行染色鉴定。
1.3.2 引物设计 根据前期筛选的猪lncRNA NONSUST 004358.1 序列及GenBank 数据库登录的猪(登录号:XM_021076521.1)、(登录号:NM_213839.1)、(登录号:NM_214315.3)和(登录号:NM_214379.1)基因序列,利用Primer Premier 5.0 软件设计lncRNA NONSUST004358.1 扩增引物与qRTPCR 引物(表1),引物由生工生物工程(上海)股份有限公司合成。

表1 qRT-PCR 引物信息
1.3.3 pEGFP-C1-lncRNA NONSUST004358.1 重组真核表达载体的构建 以前期构建的T 克隆载体pMDlncRNA NONSUST004358.1 为模板,根据设计加了酶切位点的引物进行PCR 扩增。PCR 反应体系30 μL:Taq DNA 聚合酶15 μL,上游引物(10 pmol/L)1 μL,下游引物(10 pmol/L)1 μL,100 μg/μL cDNA模板1 μL,ddHO 12 μL。PCR 反应条件:94℃变性4 min,94℃变性40 s,60℃退火50 s,72℃延伸10 min,共30 个循环,4℃保存。PCR 产物用1%的琼脂糖凝胶电泳检测,利用DNA 胶回收试剂盒对扩增产物进行回收纯化,并用限制性内切酶I 和I 对回收产物和pEGFP-C1 进行双酶切,16℃条件下经T4 DNA 连接酶连接过夜,将连接产物转化至大肠杆菌DH5,37℃培养12 h 后,挑取单菌落并进行菌液PCR 鉴定,对PCR 鉴定为阳性的菌液用质粒提取试剂盒进行质粒提取,质粒进一步双酶切鉴定,最后将酶切鉴定正确的重组质粒装样送上海英潍捷基有限公司测序。
1.3.4 qRT-PCR 方法的建立 分别在肌内和皮下前体脂肪细胞诱导分化的0、24、36、48、60、72、96 h和120 h 时提取细胞RNA 并逆转录。将构建的重组质粒pEGFP-C1-lncRNA NONSUST004358.1 转染前体脂肪细胞,36 h 后进行RNA 的提取并逆转录。以逆转录的cDNA 为模板,利用qRT-PCR 方法检测lncRNA NONSUST004358.1 和PRKAA1 在2 种前体脂肪细胞不同诱导时间中和细胞分化相关基因和的相对表达量。荧光定量PCR 体系(10 μL):2×SsoFast EvaGreen Supermix(High ROX)5 μL,上下游引物(10 µmol/L)各0.5 μL,100 μg/μL cDNA 模板1 μL,ddHO 3 μL。荧光定量PCR 反应条件:95℃预变性3 min、95℃变性30 s、56℃/60℃退火30 s、40 个循环后进行熔解曲线分析、延伸30 s、95℃终延伸50 s,每个样品3 个重复。
1.3.5 Western blot 检测细胞分化相关基因蛋白的表达量将重组质粒pEGFP-C1-LncRNA NONSUST004358.1 转染48 h 的前体脂肪细胞,利用1×RIPA 细胞裂解液进行蛋白提取,蛋白定量后加入适量的蛋白上样缓冲液在100 ℃条件下进行蛋白变性,-80℃保存备用。加入按比例稀释后的PRKAA1、FAS、HSL、PPAR一抗(山羊抗兔1:1 500)4℃过夜,TBST 洗3 次,加入二抗(山羊抗兔1:2 000)37℃孵育2 h,用-actin(鼠多抗1:1 500)作为内参标准化,利用ECL 化学发光试剂盒进行显色。
1.3.6 甘油三酯和胆固醇含量的检测 将重组质粒pEGFP-C1-lncRNA NONSUST004358.1 转染前体脂肪细胞6 h 后,加入配制的细胞诱导分化液和细胞维持液对细胞进行诱导分化,待脂滴达到饱和后,弃细胞培养液,利用1×RIPA 细胞裂解液裂解细胞并离心,取上清。按照甘油三酯和胆固醇使用说明书对其进行检测。
1.3.7 数据处理 与分析qRT-PCR 数据运用2方法进行计算分析,同时利用软件SPSS 18.0 对数据进行单因素方差分析;Western blot 灰度值运用Image Lab 软件进行计算,<0.01 表示差异极显著,<0.05 表示差异显著。
2 结果与分析
2.1 脂肪细胞的鉴定 待细胞长至85%时,利用分化诱导培养基和维持培养基对2 种细胞进行诱导。如图1 所示,在肌内前体脂肪细胞中,诱导培养2 d 时细胞形态由梭形开始向椭圆形变化;诱导培养4 d 时椭圆形细胞数量增多,出现少量脂滴;诱导培养6 d 后细胞脂滴趋于明显;诱导培养8 d,细胞出现大量脂滴。待前体脂肪细胞诱导成熟后按照油红O 染色的说明书进行鉴定。皮下前体脂肪细胞诱导分化及油红O 染色与肌内前体脂肪细胞基本一致。

图1 前体脂肪细胞油红O 鉴定
2.2 lncRNA NONSUST004358.1 和PRKAA1 在前体脂肪细胞诱导分化过程中的表达规律 qRT-PCR 结果显示,肌内前体脂肪细胞中,lncRNA NONSUST004358.1在诱导72 h 表达量最高,其次是0、48、60 h 和108 h,24 h 表达量最低,PRKAA1 在60 h 表达量最高,其次是72、48、96 h 和36 h 表达较高,108 h 表达量最低;皮下前体脂肪细胞中,lncRNA NONSUST004358.1 在诱导0 h 表达量最高,其次是72 h,在36 h 表达最低,PRKAA1 在48 h 表量最高,其次是60 h,108 h 表达量最低(图2)。两基因在2 种前体脂肪细胞中的表达基本一致,在48~96 h 表达量较高。
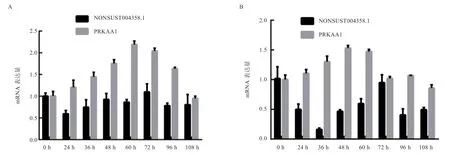
图2 lncRNA NONSUST004358.1 和PRKAA1 在前体脂肪细胞中的表达
2.3 重组质粒pEGFP-C1-lncRNA NONSUST004358.1的鉴定 酶切结果显示,在1 227 bp 和4 700 bp 左右出现单一亮带(图3),将鉴定的重组质粒送上海英潍捷基有限公司进行测序,测序结果分析发现未发生基因突变(图4),表明重组质粒pEGFP-C1-lncRNA NONSUST004358.1 构建成功。

图3 pEGFP-C1-lncRNA NONSUST004358.1 重组质粒双酶切

图4 pEGFP-C1-lncRNA NONSUST004358.1 重组质粒测序比对结果图
2.4 过表达对脂肪细胞分化相关基因表达的影响 qRTPCR 研究结果如图5 所示,肌内前体脂肪细胞中,pEGFP-C1-lncRNA NONSUST004358.1 转染后和的相对表达量高于对照组(<0.01);皮下前体脂肪细胞中,pEGFP-C1-lncRNA NONSUST0 04358.1 转染后表达量高于对照组(<0.05),低于对照组(<0.01),(<0.01)和(<0.05)高于对照组。对脂肪分化相关蛋白表达的检测结果显示,肌内前体脂肪细胞中,和表达量均高于对照组(<0.01),表达量低于对照组(<0.01);皮下前体脂肪细胞中,和表达量均低于对照组(<0.01),表达量高于对照组(<0.01)。

图5 pEGFP-C1-lncRNA NONSUST004358.1 转染后对脂肪细胞分化相关基因表达的影响
2.6 甘油三酯和胆固醇含量检测 如图6 所示,过表达lncRNA NONSUST004358.1 后,肌内前体脂肪细胞中,胆固醇含量高于对照组(<0.01),皮下前体脂肪细胞差异不显著;在皮下和肌内前体脂肪细胞中甘油三酯含量差异不显著。
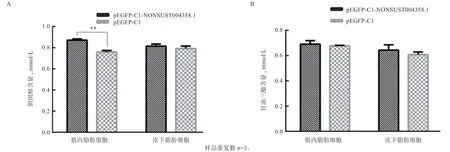
图6 甘油三酯(A)和胆固醇(B)含量检测
3 讨 论
脂肪是动物体内重要的储能物质,与机体能量平衡密切相关,在畜牧动物体内,脂肪的分布和含量影响畜产品的肉品质和风味。脂肪是由中胚层的特化细胞群分化而来,其分化是一个复杂的调控过程,伴随着细胞结构和功能的改变,受到、等多种转录因子和PKA(Protein Kinase A System)和MAKP(Mitogen-Activated Protein Kinase)等 信号通路的调控。近年的研究发现,lncRNA 可通过调控脂肪相关基因表达从而参与脂肪细胞的分化与形成。Chen 等研究发现,lnc-U90926 过表达会抑制3T3-L1 脂肪细胞的分化,抑制脂质蓄积,降低过氧化物酶体增殖物激活受体2(Peroxisome Proliferator-Activated Receptor-2,PPAR2)、脂肪酸结合蛋白4(Fatty Acid Binding Protein,4FABP4)和脂联素(Adiponectin,AdipoQ)的表达。Xiong 等研究发现,敲除lncRNA AK079912 可抑制褐色前脂肪细胞的分化,导致脂肪沉积的减少和脂肪代谢相关基因(脂蛋白脂肪酶:;;甘油三酸酯脂肪酶:)的下调;猪前体脂肪细胞分化过程中,干扰lncRNA 00046716的表达导致FASN mRNA 和蛋白表达的下降,抑制了脂肪细胞分化,与miRNA-149 结合后会影响FASN 的表达,调控猪前体脂肪细胞分化与甘油三酯在成熟脂肪细胞中的积累。
本课题组前期通过PRKAA1 筛选出了相关lncRNA NONSUST004358.1,其在从江香猪的肝脏、脂肪和背最长肌组织中具有较高表达且高于大白猪,暗示其可能参与从江香猪的脂肪代谢与沉积。本实验分离培养从江香猪原代皮下和肌内前体脂肪细胞,利用qRT-PCR检 测lncRNA NONSUST004358.1 和PRKAA1 在前体脂肪细胞分化过程中的表达,分析其表达规律。qRTPCR 结果显示,在肌内和皮下前体脂肪细胞分化过程中,PRKAA1 与lncRNA NONSUST004358.1 均呈先上升后下降的表达模式,这与lncRNA 00046716 在猪前体脂肪细胞分化过程中的表达一致,通过与从江香猪前体脂肪细胞分化过程中脂肪相关的基因的表达分析发现,lncRNA NONSUST004358.1 与PPAR、FAS 和ACC等在肌内前体脂肪细胞分化过程中具有相同的表达趋势,暗示其可能在肌内前体脂肪细胞分化过程中发挥重要调控作用;皮下前体脂肪细胞中,NONSUST004358.1 虽在分化前期(0~36 h)与和的表达不同,但在36 h 之后却表现出相同的表达规律,表明lncRNA NONSUST0043 58.1 可能在皮下前体脂肪细胞分化后期对脂肪细胞的分化具有调控作用。lncRNA NONSUST004358.1 在肌内与皮下前体脂肪细胞分化过程中表现出不同的表达规律,表明其在皮下和肌内前体脂肪细胞分化过程中具有不同的功能。通过检测前体脂肪细胞分化过程中lncRNA NONSUST004358.1 和的表达量,分析其表达规律,为后续研究其在脂肪分化过程中的作用机制提供试验数据。
为探究lncRNA NONSUST004358.1 在前体脂肪细胞中对脂肪分化相关基因的影响,本实验验通过构建重组真核表达载体pEGFP-C1-lncRNA NONSUST004358.1,转染肌内和皮下前体脂肪细胞,利用qRT-PCR 和Western blot 检测脂肪代谢相关基因的表达,qRT-PCR结果显示,肌内前体脂肪细胞中,和的表达量均上调;皮下前体脂肪细胞中,和表达量上调,表达下调。表明肌内和皮下前体脂肪细胞中lncRNA NONSUST00 4358.1 在转录水平对和基因具有正向调控作用,对FAS 在皮下前体脂肪细胞中具有负调控。Western blot 结果显示,肌内前体脂肪细胞中,过表达lncRNA NONSUST004358.1 上调了PRKAA1、FAS 和PPAR蛋白表达 量,HSL表达下调;皮下前体脂肪细胞中,FAS 和PPAR蛋白表达量上调,PRKAA1 和HSL 蛋白表达量下调。说明lncRNA NONSUST004358.1 在肌内和皮下前体脂肪细胞中对脂肪合成基因FAS 和PPAR的蛋白具有正向调控,暗示其参与脂肪细胞的分化与脂肪的形成。FAS和PPAR作为作为脂类代谢的重要调节因子,其活性的高低对动物体脂沉积具有重要意义,过表达lncRNA NONSUST004358.1 均促进肌内与皮下前体脂肪细胞中FAS、PPARmRNA 与蛋白的表达,表明lncRNA NONSUST004358.1 是影响猪体脂沉积的重要候选基因。甘油三酯和胆固醇作为脂肪细胞的主要代谢产物,脂肪细胞分化程度影响甘油三酯和胆固醇含量。本实验通过测定转染pEGFP-C1-lncRNA NONSUST004358.1 后肌内和皮下前体脂肪细胞中甘油三酯和胆固醇含量发现,含量均上调。表明lncRNA NONSUST004358.1 在转录水平和蛋白水平调控PRKAA1、FAS、PPAR和HSL 的表达,促进猪皮下和肌内前体脂肪细胞的分化与脂肪的形成。lncRNA 参与脂肪代谢的主要调控机制是直接与靶基因结合、招募转录物、作为辅助因子或通过与 miRNAs 竞争性结合调控靶基因的表达,而lncRNA NONSUST004358.1对PRKAA1、FAS、PPAR和HSL 的具体调控机制还需进一步研究。
4 结 论
本实验中,lncRNA NONSUST004358.1 在肌内和皮下前体脂肪细胞分化过程中的表达均呈现下降、上升、再下降的表达模式,能调控脂肪沉积相关基因的表达,影响脂肪细胞中油三酯和胆固醇含量,研究结果为后续探究lncRNA NONSUST004358.1 在脂肪细胞分化中的作用机制提供了理论支撑。
猜你喜欢
中学生物学(2022年8期)2022-10-13
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
中国畜牧杂志(2022年6期)2022-06-13
中国典型病例大全(2022年7期)2022-04-22
科学大观园(2020年22期)2020-11-30
三农资讯半月报(2020年11期)2020-06-21
江苏农业学报(2019年1期)2019-09-10
航空兵器(2018年6期)2018-02-26
中外医疗(2016年29期)2016-11-30
中国现代医生(2016年25期)2016-11-19
