
湖羊瘤胃微生物葡聚糖酶基因IDSGluc5-31 的克隆、表达与性质研究
2022-09-14 02:36高德英曹佳雯孙小宝何波王佳堃王谦
中国畜牧杂志 2022年9期
李 锘,高德英,曹佳雯,孙小宝,何波,3,王佳堃,王谦*
(1.浙江大学动物科学学院奶业科学研究所,浙江杭州 310058;2.浙江万里学院生物与环境学院,浙江宁波 315100;3.上海交通大学生命科学技术学院微生物代谢国家重点实验室,上海 200240)
-葡聚糖是一类由--葡萄糖残基通过-1,3 和-1,4-糖苷键连接形成的线性多聚糖。作为一类非淀粉多糖(Non-Starch Polysaccharide,NSP),-葡聚糖是植物性饲料中一种重要的抗营养因子,它粘稠性高且难降解,难以被单胃动物消化吸收利用,导致动物生产性能降低及饲料利用率下降。-葡聚糖酶是指能降解-葡聚糖的一大类糖苷水解酶(Glycoside Hydrolases,GHs),可作为饲料添加剂、多糖改性增溶剂等应用于饲料和食品工业。-葡聚糖酶能够消除-葡聚糖的抗营养作用,同时,-葡聚糖的降解产物主要是聚合度为2~5 的低聚纤维寡糖,这些功能性寡糖能作为益生元促进消化道有益微生物的增殖,维持动物和人体健康。
目前,-葡聚糖酶主要从环境微生物中分离纯化或克隆后异源表达获取。瘤胃微生物能够产生包括-葡聚糖酶在内的碳水化合物活性酶(Carbohydrate-Active Enzymes,CAZymes)。由于长期适应于瘤胃特殊的生长环境,绝大多数瘤胃微生物不能被纯培养,因此采用常规方法难以直接从中获取-葡聚糖酶。课题组前期采用宏转录组学技术从湖羊瘤胃微生物中发现436 个GH5 家族编码基因。本研究进一步扩增并异源表达-葡聚糖酶基因,分析重组蛋白的酶学性质、动力学参数和水解产物,为高效降解植物性饲料和饲用酶制剂研发建立基础。
1 材料与方法
1.1 实验材料 表达载体pET-28a(+)购自Novagen 公司;Super Pfx 聚合酶购自康为世纪公司;ClonExpressII One step Cloning Kit 购自Vazyme 公司;质粒提取试剂盒购自Omega 公司;PCR 产物纯化试剂盒、割胶回收试剂盒购自上海生工公司;大麦葡聚糖、苔藓地衣多糖和纤维寡糖购自Megazyme 公司;核酸与蛋白Marker、Ni-NTA agarose、卡那霉素(kanamycin,Kan)和异丙基---硫代吡喃半乳糖苷(IPTG)购自上海生工公司;湖羊瘤胃内容物样品和原核表达宿主Escherichia coli BL21(DE3)为本实验室保存。
1.2 重组工程菌构建与蛋白表达 参照何波的方法提取湖羊瘤胃内容物总RNA 并反转录成cDNA。利用IDSGluc5-31-HI(5'ACAGCAAATGGGTCGCGGAT CCATGCAGCGTATCAAAATCTTAGCG3')和IDSGluc5-31-I(5'CTTGTCGACGGAGCTCGAATTCTTAT GCCGCAGGCGCTTCC3')引物(下划线为表达载体同源臂序列)和Super Pfx 聚合酶扩增cDNA 样品。纯化后的PCR 产品与经H I 和RI 双酶切的pET-28a(+)充分混合 后,利用ClonExpressII One step Cloning Kit 进行同源重组,BL21(DE3)感受态细胞用于反应产物的热激转化,转化完成后涂布含50 μg/mL Kan 的LB 平板。37℃培养16~20 h 后,挑取单菌落进行菌落PCR 扩增,并提取质粒送至上海生工公司测序。
利用1 mmol/L IPTG 诱导工程菌BL21/pET28a/表达重组葡聚糖酶,在16℃,100 r/min下孵育16 h 后,收集菌体。按本课题组前期建立的方法,进行均质破碎获得粗酶液。
1.3 蛋白纯化与电泳分析 将亲和Ni-NTA agarose 填料按1:100 与上述粗酶液混合,加入终浓度为20 mmol/L的咪唑,冰上缓慢震荡孵育1 h。接着利用含20~500 mmol/L 咪唑的磷酸盐缓冲液梯度洗脱目标蛋白。之后,利用Millipore AmiconUltra 超滤柱(3 ku)去除残余咪唑获得纯化蛋白,用于SDS-PAGE 电泳和酶谱分析。
酶谱分析是在聚丙烯酰胺凝胶中加入终浓度为0.5% 的葡聚糖或地衣多糖底物,在最适反应条件下测 定IDSGLUC5-31 对底物的 降解活性。将20 μL IDSGLUC5-31 纯化酶液与5 μL 5×SDS 上样缓冲液充分混匀,于4℃下放置20 min。蛋白上样量为10 μL,以BSA 为阴性对照。90 V 恒压电泳40 min,120 V 恒压电泳2 h。电泳结束后,将凝胶置于25%异丙醇中复性3 次,每次10 min。复性后的凝胶置于1×PBS 缓冲液中,37℃反应过夜。用0.1%刚果红染色20 min,之后用1 mol/L NaCl 溶液中至脱色完全。
1.4 酶学性质分析 采用3,5 二硝基水杨酸(3,5-Dinitrosalicylic acid,DNS)法测定葡聚糖酶活性,采用Bradford 法测定蛋白浓度,操作方法:将20 μL IDSGLUC5-31 纯化酶液(约13 μg,下同)与50 μL 0.5%葡聚糖底物充分混合,在相应温度或pH 条件下反应15 min 后,加入70 μL DNS 试剂,99℃保温10 min。冷却至室温后,使用SpectraMax M3 酶标仪(Molecular Devices)测定OD,同时设置热处理失活的酶为对照和3 组平行试验。
1.4.1 最适温度和热稳定性 测定IDSGLUC5-31 在pH6.0,20~90℃范围内的反应活性,以最高活性时的温度为最适温度,计算各温度条件下相对活性。热稳定性分析是在pH6.0 条件下,将酶液分别在40~60℃下保 温60 min。在0、5、10、20、30、40 和60 min 取样,pH6.0,40℃下测定葡聚糖酶活性。以反应起始点为100%,计算各试验组的残余酶活。
1.4.2 最适pH 和pH 稳定性 测定IDSGLUC5-31 在40℃,pH 2.2~10.6 缓冲液(柠檬酸/磷酸缓冲液pH 2.2~8.0;碳酸钠/ 碳酸氢钠缓冲液pH 8.9~10.6)范围内的反应活性,以最高活性时的pH 为最适pH,计算各pH 条件下相对活性。pH 稳定性分析是在40℃下,将酶液分别在4℃ pH 2.2~10.6 缓冲液中孵育30 min,在pH 6.0,40℃下测定葡聚糖酶活性。以反应起始点为100%,计算各实验组的残余酶活。
1.4.3 金属离子及化学试剂耐受能力 将IDSGLUC5-31 与0.1~10 mmol/L 的金属离子(Na、Ca、Mg或Cu)或其他化学试剂(SDS 或EDTA)在4℃下保温30 min,在pH 6.0,40℃下测定葡聚糖酶活性。以反应起始点为100%,计算残余酶活。
1.4.4 动力学参数 将IDSGLUC5-31 分别与0.5~10 mg/mL的葡聚糖、地衣多糖、羧甲基纤维素、微晶纤维素、磷酸溶胀纤维素、糊精或木聚糖底物充分混合,在pH 6.0,40℃下测定葡聚糖酶活性。利用GraphPad Prism 8 软件(San Diego,CA)计算和。
1.5 底物水解分析
1.5.1 薄层色谱法(Thin-Layer Chromatography,TLC)采用TLC 法分析IDSGLUC5-31 催化葡聚糖底物的水解产物。将5 mg/mL 的大麦葡聚糖或地衣多糖底物置于40℃下预热5 min,加入1 µg IDSGLUC5-31,40℃,pH 6.0 反应0.5 h 和20 h 后分别取样1 mL,95℃孵育30 min 终止反应。4℃,13 000 r/min 离心30 min,取0.3 µL 反应液或1.0 mg/mL 标准品混合液(含葡萄糖、纤维二糖、纤维三糖、纤维四糖和纤维五糖)轻轻点在薄层板(Merck 公司)上,重复上样5 次。流动相为正丁醇:乙酸:水体积比为2:1:1,展开到距薄层板顶端1 cm 处取出,置于通风橱待平板充分干燥后,置入展层液中二次展开,再干燥后放入显色液(硫酸:甲醇=5:95,v/v;0.5%-萘酚,w/v)反应2 min,100℃ 10 min高温显色。
1.5.2 高效液相色谱法(High Performance Liquid Chrom atography,HPLC) 进一步采用HPLC法分析IDSGLUC5-31催化葡聚糖底物的水解产物。取2 mL IDSGLUC5-31纯化酶液(约120 μg)分别与5 mL 0.5%葡聚糖或0.5%地衣多糖底物混合,在最适反应条件下孵育20 h,分别于8 h 和20 h 取样。于95℃保温30 min,13 000 r/min离心15 min,取上清,设置3 组重复试验。利用Asahipak NH2P-50 4E 色谱柱(Shodex 公司)和LC-1200 高效液相色谱仪(Agilent 公司),流动相为乙腈:水=65:35,柱温40℃,流速0.8 mL/min,采用RID-20A 时差折光检测器分析底物水解样品。
1.6 统计分析 图片绘制和动力学分析用Graphpad Prism 8 完成,数据统计采用Student's-test。* 表示差异显著(<0.05);**表示差异极显著(<0.01)。
2 结果与分析
2.1 葡聚糖酶序列分析 本试验成功地从湖羊瘤胃未培养微生物cDNA 中扩增得到1 个葡聚糖酶基因,其开放阅读框长度为1 245 bp,编码415 个氨基酸。该蛋白理论等电点为4.89,理论分子量为46.6 ku(https://web.expasy.org/compute_pi/)。SingalP 5.1和Pfam 分析显示,IDSGLUC5-31 蛋白包含一段24个氨基酸的信号肽(http://www.cbs.dtu.dk/services/SignalP/),成熟肽中包含1 个GH5 家族催化结构域(http://pfam.xfam.org/)。同源建模分析显示(图1),该蛋白包含(/)桶状催化结构域,呈现GH5 家族的典型结构特征(https://www.swissmodel.expasy.org/interactive)。

图1 IDSGLUC5-31 蛋白三维结构预测
系统发育进化树和BLAST 分析表明,IDSGLUC5-31与梭菌目细菌(GenBank No:SNT95308.1)来源的GH5 家族蛋白聚成一簇(图2),氨基酸序列相似性分别为78.27%,该蛋白的功能与性质未被研究。通过序列比对分析IDSGLUC5-31 的成熟肽与QTE70932.1、MEB6847130.1、MBD5081498.1、WP051109229.1 等具有保守的氨基酸序列,其中保守催化残基区域E198和E332 与已报道的GH5 家族内切葡聚糖酶的保守序列的规律一致(图3)。

图2 IDSGLUC5-31 系统进化树分析

图3 葡聚糖酶氨基酸序列比对
2.2 葡聚糖酶IDSGLUC5-31 异源表达与电泳分析 利用IPTG 诱导表达重组工程菌BL21/pET28a/,粗蛋白质经Ni-NTA agarose 亲和层析后,获得分子量约为48 ku 的IDSGLUC5-31 蛋白(图4-A)。进一步通过酶谱分析发现,IDSGLUC5-31 能有效降解大麦葡聚糖和苔藓地衣多糖(图4-B 和4-C)。
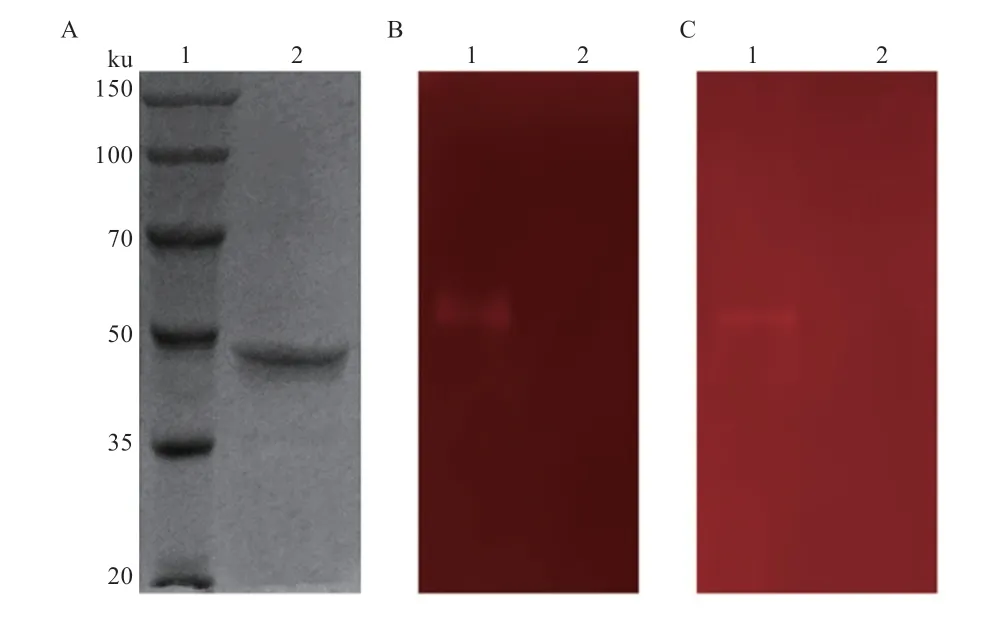
图4 IDSGLUC5-31 SDS-PAGE 和酶谱分析
2.3 IDSGLUC5-31 的酶学性质 酶学性质分析显示,IDSGLUC5-31 的最适反应温度和pH 分别为40℃和6.0(图5-A 和5-B)。在40℃以下较为稳定,处理1 h 后,仍能保持79.95%的残余活性。当温度上升至50℃以上,该蛋白迅速失活。50℃下处理5 min,蛋白几乎完全失活(图5-C)。IDSGLUC5-31 在pH 5.0~7.0 较为稳定,处理30 min 后仍能保持70%以上活性(图5-D)。

图5 IDSGLUC5-31 的酶学性质
2.4 IDSGLUC5-31 的动力学参数 活性底物谱分析显示,IDSGLUC5-31 能高效催化大麦葡聚糖和苔藓地衣多 糖,分别为508.7 和240.1 μmol/(min·mg)。且底物亲和力较强,分别为1.725 和2.628 mg/mL(表1)。然而,该蛋白不能降解羧甲基纤维素钠、微晶纤维素、磷酸溶胀纤维素、糊精或木聚糖。

表1 IDSGLUC5-31 动力学参数
2.5 金属离子和化学试剂对IDSGLUC5-31 活性的影响 不同类型和不同浓度金属离子和化学试剂对IDSGLUC5-31 活性的影响如图6 所示,该蛋白对Na的耐受能力较强,经0.1~10 mmol/L Na处理30 min后,活性几乎没有改变;低浓度(0.1 mmol/L)Ca、Mg或SDS 不影响IDSGLUC5-31 的催化活性。当浓度提高至10 mmol/L 时,蛋白活性下降(<0.01);IDSGLUC5-31 在EDTA 和Cu存在的环境中稳定性较差,经孵育30 min 后活性迅速下降,甚至完全失活(<0.01),且伴有蛋白析出。
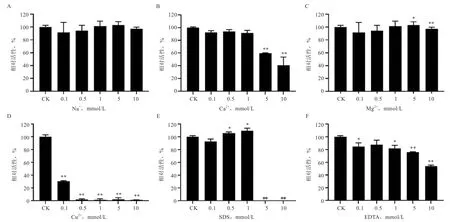
图6 金属离子和化学试剂对IDSGLUC5-31 活性的影响
2.6 IDSGLUC5-31 水解多聚糖的产物分析 采用TLC法分析IDSGLUC5-31 水解地衣多糖和大麦葡聚糖的产物可以发现,IDSGLUC5-31 水解地衣多糖和大麦葡聚糖的主要产物为纤维二糖至纤维五糖,其中纤维三糖含量最高(图7-A)。进一步利用HPLC 分析IDSGLUC5-31 催化大麦葡聚糖和苔藓地衣多糖的水解产物发现:经IDSGLUC5-31 处理20 h 后,大麦葡聚糖的水解产物为纤维二糖、纤维三糖、纤维四糖和纤维五糖,产量分别为0.912、0.94、0.56 和0.24 mg/mL(图7-B);苔藓地衣多糖的水解产物也为纤维二糖、纤维三糖、纤维四糖和纤维五糖,产量分别为0.10、0.87、0.23 和0.1 mg/mL(图7-C)。2 种底物的主要水解产物均为纤维三糖和纤维四糖,推测IDSGLUC5-31 是一种内切-葡聚糖酶。
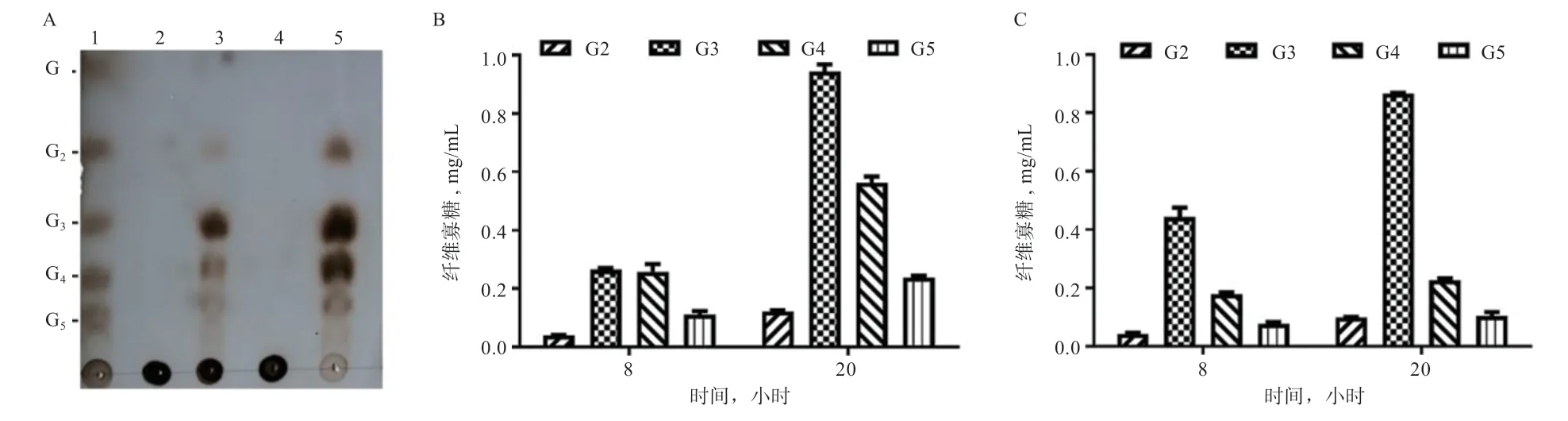
图7 IDSGLUC5-31 水解产物分析
3 讨 论
微生物纯培养与无菌技术长期以来是微生物学研究的基石,但由于难以满足来自瘤胃等极端环境微生物的生长需求,瘤胃微生物的纯培养和相关功能基因的研究较为困难。宏基因组学和宏转录学技术改变了传统纯培养的理念,以样本总DNA 或RNA 为研究对象,分析总微生物的功能基因与蛋白信息。Wang 等利用宏基因组学技术构建了娟珊牛瘤胃微生物的14 000 个fosmid 文库,获得120 个对碳水化合物底物具有水解活性的克隆,其中降解纤维素、木聚糖、淀粉和七叶苷底物的fosmid 克隆分别有34、52、1 和33 个,共编码575 个潜在CAZymes 或与底物转录调控、转运及信号传导相关的功能基因。He 等利用宏转录组学技术从湖羊瘤胃微生物中发现的238 万多个unigenes 中2.65%为糖苷水解酶,其中与纤维素降解相关的功能基因主要分布于GH3、5 和9 家族。这些研究表明,瘤胃微生物基因资源非常丰富,利用多组学技术能有效筛选、获取CAZymes。
本研究从湖羊瘤胃微生物cDNA 中获得的葡聚糖酶IDSGLUC5-31 的底物特异性好,对大麦葡聚糖酶、地衣多糖的催化活性和底物亲和力均较高,然而,该蛋白的热稳定性不佳,在40˚C 以上孵育后迅速失活。由于反刍动物瘤胃温度通常为38~41˚C,推测IDSGLUC5-31 的反应温度可能是与消化道环境温度相适应的结果。为便于运输、提高适口性和饲料转化率,饲用酶制剂在工业化生产中需要进行饲料制粒。根据制粒对象和工艺的不同,制粒温度需达到80~90˚C 甚至130˚C 以上。而IDSGLUC5-31 在50℃下处理后就快速失活,不能满足工业化生产的要求。因此,后续可以通过定向进化、分子环化等酶工程手段进一步提升其热稳定性。
植物性饲料经糖苷水解酶处理后,可以降解细胞壁中的NSPs,消除其抗营养作用,对肉鸡、猪等单胃动物甚至奶牛、绵羊等反刍动物均具有促生长或提高生产性能的作用。本研究中,IDSGLUC5-31 可以催化大麦葡聚糖和苔藓地衣多糖生成纤维三糖和纤维四糖,提示是一种内切-葡聚糖酶。IDSGLUC5-31 一方面消除植物性饲料中葡聚糖的抗营养作用,另一方面产生的低聚寡糖可以作为益生元促进动物消化道中双歧杆菌和乳酸菌等有益菌的定植,进而维持动物机体健康。
4 结 论
本研究将湖羊瘤胃微生物来源的葡聚糖酶基因进行异源表达获得重组蛋白IDSGLUC5-31。该蛋白的最适反应温度和pH 分别为40℃和6.0,且在40℃以下或pH5.0~7.0 内较为稳定,对0.1~10 mmol/L Na、0.1~1 mmol/L Ca或Mg表现较强耐受能力。此外,IDSGLUC5-31 能催化大麦葡聚糖或苔藓地衣多糖生成纤维三糖和纤维四糖。本研究的结果为从瘤胃微生物中挖掘新型CAZymes 提供参考,并为研发饲用酶制剂和制备功能性寡糖建立基础。
猜你喜欢
教育周报·教育论坛(2020年3期)2020-10-21
老年博览·上半月(2019年1期)2019-09-10
医学食疗与健康(2019年9期)2019-09-10
农民致富之友(2019年25期)2019-09-03
新农业(2019年7期)2019-07-27
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
安徽农业科学(2018年18期)2018-05-14
科技资讯(2017年12期)2017-06-09
食品安全导刊(2014年7期)2014-10-21
