
利用DNA 条形码技术对茶黑刺粉虱的鉴定与分析
2022-09-16 06:05张金峰王威锐孟泽洪
湖南农业科学 2022年8期
张 莉,李 帅,张金峰,王威锐,3,孟泽洪
(1.贵州省农业科学院草业研究所,贵州 贵阳 550006;2.贵州省农业科学院茶叶研究所,贵州 贵阳 550006;3.贵州大学茶学院,贵州 贵阳 550025)
茶黑刺粉虱(Aleurocanthus camelliaeKanmiya &Kasai)隶属于半翅目(Hemiptera)粉虱科(Aleyrodidae)粉虱亚科(Aleyrodinae)刺粉虱属(Aleurocanthus),是茶园里最常见的粉虱类害虫[1]。茶黑刺粉虱主要通过吸食叶片汁液、诱发煤烟病造成危害,从而影响树势,同时生产季节成虫的活动也会干扰茶农的劳作[2]。寄主为茶的茶黑刺粉虱(A. camelliae)与寄主为柑橘等植物的橘黑刺粉虱(A. spiniferus)的分类尚存在一定的争论。早些年,一般认为为害茶树的黑刺粉虱即橘黑刺粉虱[3-4];但基于生态学、形态学和分子进化与系统发育学的研究结果表明,茶黑刺粉虱应为独立的物种[5]。目前,茶黑刺粉虱和橘黑刺粉虱的线粒体基因组均已公布[6-7]。但由于茶黑刺粉虱在很长一段时间内被视为与橘黑刺粉虱同种,因此现已知的这2种粉虱的分子数据存在一定的混乱,需要进行全面细致的区分。
DNA 条形码技术(DNA Barcoding)是由加拿大的Hebert 博士首次提出的。该技术基于线粒体DNA 细胞色素C 氧化酶亚基I(mitochomdrail DNA cytochrome coxidase subunit I, mtDNACOI)基因进行核苷酸序列分析,可利用mtDNACOI中一段特定区域(约658 bp)鉴定不同的物种[8]。目前,该技术已在诸多昆虫中得以验证与应用[9-10]。
茶树主要害虫茶黑刺粉虱的mtDNACOI基因数据极少且均不是目前鉴定上通用的DNA 条形码特定标准区域序列片段[5],如此不便于种类间的横向比较,因此,笔者拟通过试验获取茶黑刺粉虱DNA 条形码特定标准区域(658 bp)的分子序列数据,并进行遗传距离及系统发育分析,以期为进一步掌握茶黑刺粉虱DNA 条形码以及分子鉴定提供基础数据。
1 材料与方法
1.1 供试材料
供试虫源采集信息如表1 所示,所有样本均采自茶叶寄主,将采集到的粉虱置于装有无水乙醇的2 mL 冻存管中,冷冻于-20℃冰箱中保存备用。
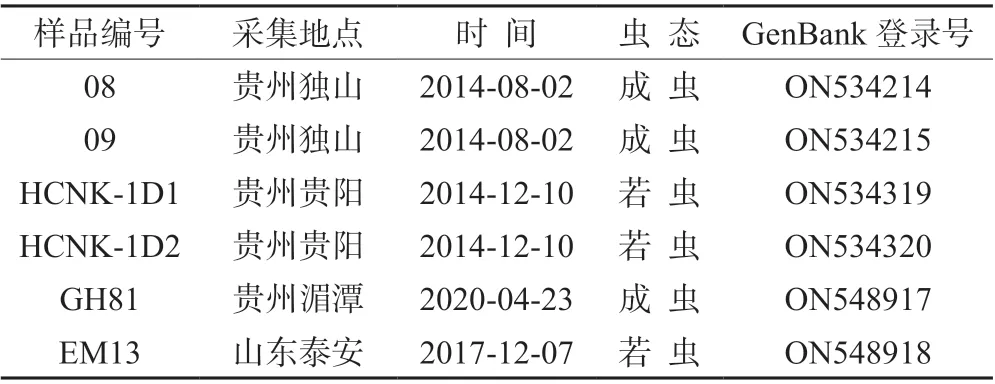
表1 试验样品信息
1.2 DNA 提取及PCR 扩增
采用Qiagen DNEasy Tissue kit 试剂盒提取单头虫体的DNA。虫体经研磨破碎后,按照说明书进行操作,提取的DNA 用1%琼脂糖电泳进行检测,再采用通用引物LCO1490/ HCO2198[11]对其mtDNACOI基因序列特定区域片段[8]进行PCR 扩增。PCR 扩增体系总体积为50 μL,包括10×PCR 缓冲液5 μL,25 mmol/L MgCl23 μL,2.5 mmol/L 的dNTP 混合液4 μL,10 μmol/L 的上下游引物各1.5 μL,5 U/μL 的TaKaRa TaqDNA 聚合酶0.3 μL 和DNA 模板2 μL,加ddH2O至50 μL。PCR 反应程序为:94℃预变性4 min;94℃30 s,Tm 40 s,72℃ 30 s,34 个循环;72℃ 10 min;4℃保存。PCR 产物经电泳检测后送上海立菲生物技术有限公司及北京诺赛基因组研究中心有限公司进行双向测序。
1.3 数据分析
序列的拼接使用DNAMAN 软件进行,结合测序峰值图进行正反链的拼接校对,将拼接好的序列在NCBI 网站上利用BLAST 程序进行序列同源性搜索,采用CLUSTALX2.0 进行序列比对。比对后的结果采用Mega6 软件[12]进行序列组成分析、遗传距离比较和NJ 树构建。下载已知的黑刺粉虱线粒体基因组数据KJ437166 和KU761949,并利用BIOEDIT 软件截取获得的同源DNA 条形码特定区域658 bp 片段序列进行聚类分析。
2 结果与分析
2.1 DNA 条形码特定区域片段的序列组成分析
如表1 所示,序列拼接后去掉两端引物序列,长度为658 bp,GeneBank 登录号分别为ON534214、ON534215、ON534319、ON534320、ON548918、ON548917。序列碱基组成特征为:T(42.8%)、C(13.7%)、A(22.7%)、G(20.8%),A+T 含量明显高于G+C 的含量。将获得的序列在NCBI 数据库中进行BLAST 同源性比对,结果显示,贵州湄潭(GH81)、独山(08、09)以及山东泰安(EM13)的样本序列完全一致,与KJ437166 序列对应片段高度吻合,仅在1 个碱基位点上有所差异,序列相似度为99.85%;贵州贵阳(HCNK-1D1、HCNK-1D2)的茶黑刺粉虱样本序列仅为一种单倍型,该序列同KU761949 对应片段的序列高度吻合,仅在2 个碱基位点上有所差异,序列相似度为99.70%。
2.2 DNA 条形码特定区域片段的比较分析
将试验获得的茶黑刺粉虱DNA 条形码特定区域片段序列与在GenBank 上下载的已知黑刺粉虱对应的同源片段序列KJ437166 和KU761949 基于Kimura 2-parameter 参数进行遗传距离分析,选择与茶黑刺粉虱近缘的刺桐粉虱Tetraleurodes acaciae(Quaintance)(GenBank 登录号:AY521262)[6]作为外群,结果见表2,茶黑刺粉虱DNA 条形码特定标准区域的序列遗传距离为0~8.7%,与外群刺桐粉虱遗传距离为28.9%~29.8%,与外群种的遗传距离远大于茶黑刺粉虱种内遗传距离。利用邻接法(Neighbor joining method,NJ 法)构建系统发育树,系统树各分支的置信度(boostrap)均进行1 000 次的重复检验,结果如图1 所示,茶黑刺粉虱与外群种刺桐粉虱明显属于不同的分支,同时6 个茶黑刺粉虱样本明显分出2 大分支,其中贵州湄潭(GH81)、独山(08、09)和山东泰安(EM13)以及KJ437166 聚为一分支,贵州贵阳样本(HCNK-1D1、HCNK-1D2)和KU761949 聚为另一分支。
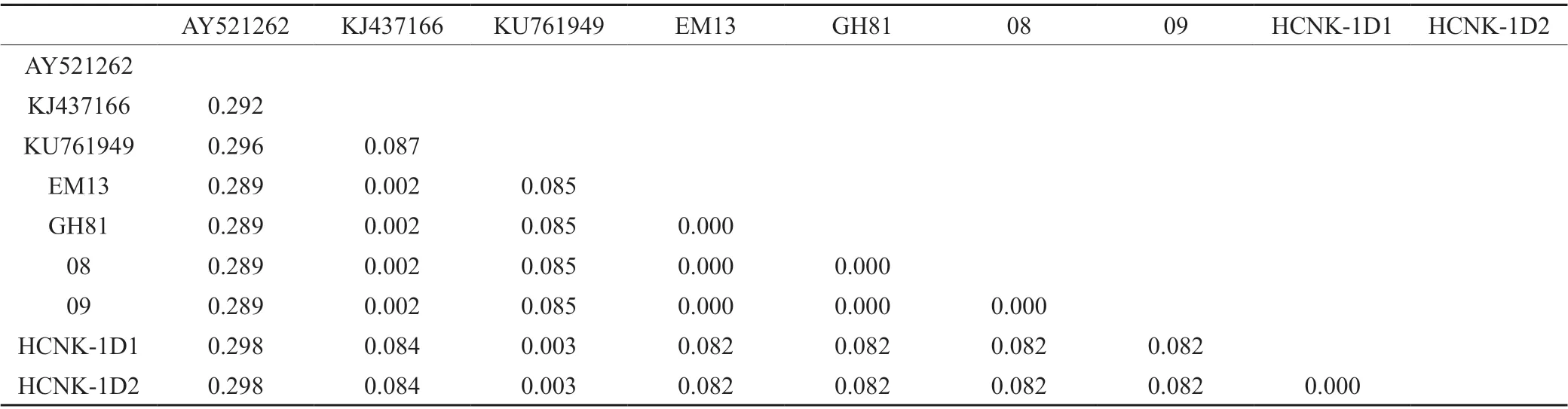
表2 遗传距离分析
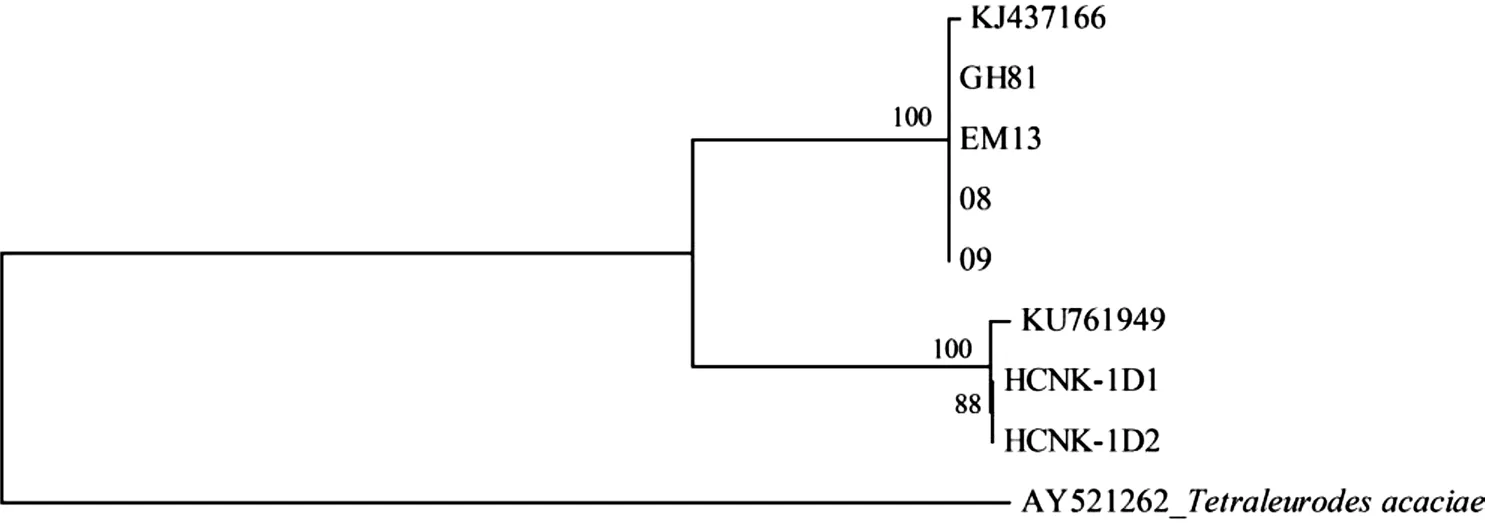
图1 基于mtDNA COI 基因DNA 条形码序列构建的茶黑刺粉虱NJ 树
3 讨 论
3.1 茶黑刺粉虱的COI 基因分化
COI基因在昆虫物种鉴定分类上应用极为广泛,其中关于茶黑刺粉虱A. camelliae COI基因的研究也有相关报道[5,13-15],例如Chen 等[6]测定的线粒体基因组全长序列KU761949。但不同研究者间分析所用的COI序列片段不尽相同,并且有些研究未将序列信息上传至GenBank[13,15]。目前,GenBank 数据库中除去线粒体基因组全长序列外,其他已知的COI基因序列仅11 条,并且与DNA 条形码标准序列片段重合区域极窄,不利于各粉虱种类间的横向比较。该研究提供了6 个茶黑刺粉虱mtDNACOI基因特定的DNA 条形码标准区域片段序列,并将序列信息上传至GenBank,登录号分别为ON534214、ON534215、ON534319、ON534320、ON548917、ON548918。
学者们关于茶黑刺粉虱COI基因分化程度的研究结果也有所不同,这与研究限定的地理范围有一定的关系。Kanmiya 等[5]选取了COI基因中一段759 bp的序列进行茶黑刺粉虱遗传分化研究,结果发现茶黑刺粉虱在日本7 个地理种群间序列相似度达99.74%,虽具有2 种单倍型,但单倍型间仅相距2 个碱基差异。胡雅辉等[13]选取了COI基因660 bp 的序列片段进行茶园黑刺粉虱遗传分化研究,发现湖南茶园内不同地理种群黑刺粉虱遗传距离为0~5.3%。Uesugi 等[14]对来自中国和日本的茶黑刺粉虱进行分子序列分析,发现2 国的茶黑刺粉虱已经明显分化为3 个分支。该研究基于COI基因特定的DNA 条形码标准区域658 bp的片段序列对6 种不同来源的茶黑刺粉虱进行的遗传距离分析,发现它们之间的遗传距离为0~8.7%,并且分化为2 大分支,其中贵州湄潭(GH81)、独山(08、09)和山东泰安(EM13)以及KJ437166 聚为一分支,贵州贵阳样本(HCNK-1D1、HCNK-1D2)和KU761949 聚为另一分支。
3.2 茶黑刺粉虱和橘黑刺粉虱的分类地位
Kanmiya 等[5]基于生态学、形态学和分子系统发育研究分析提出茶黑刺粉虱A. camelliae应为单独成立的物种。胡雅辉等[13]研究发现,柑橘黑刺粉虱种群和茶园黑刺粉虱种群的遗传距离明显高于柑橘黑刺粉虱不同地理种群间遗传距离和茶园黑刺粉虱不同地理种群间的遗传距离,并推测柑橘黑刺粉虱和茶园黑刺粉虱可能属于黑刺粉虱的不同生物型或不同种。陈瑶等[15]结合分子鉴定和形态观察发现,贵州省12 个茶园的黑刺粉虱为茶黑刺粉虱A. camelliae,并与橘黑刺粉虱A. spiniferus属于不同分支(种)。Uesugi 等[14]的研究也表明,在中国和日本,以柑橘为寄主的黑刺粉虱和以茶为寄主的黑刺粉虱应为不同种。该研究基于COI基因中DNA 条形码区域片段构建了系统树,发现茶黑刺粉虱不同地理种群可单独作为一分支,与外群刺桐粉虱分属不同类群。
Uesugi 等[14]利用COI基因674 bp 序列片段分析得出,茶黑刺粉虱分化为单倍型B1、B2 和单倍型群B3,采自我国安徽(宣城)、浙江(金华、宁德、温州)、福建(福安)、广东(英德)、江西(青原)、湖南(长沙)、四川(成都)的茶黑刺粉虱均为B1 单倍型,来自重庆永川的种群均为B2 单倍型,而检疫截获的来自中国山茶科植物的茶黑刺粉虱既有B1 单倍型,也有B3单倍型;采自日本的茶黑刺粉虱均为B1 单倍型,这一结果同Kanmiya 等[5]建立茶黑刺粉虱A. camelliae这一新种时的数据一致。Chen 等[6]发布了采自重庆永川的茶黑刺粉虱的线粒体基因组序列(登录号为KU761949),比对后发现,其与Uesugi 等[14]提出的B2 单倍型结果一致。Chen 等[7]发布了采自浙江杭州的黑刺粉虱线粒体基因组序列(登录号为KJ437166),但研究中并未提及其寄主,只是给出了鉴定种名为橘黑刺粉虱A. spiniferus;将其与Uesugi 等[14]提出的3 种单倍型数据比对后发现,该序列与B1 单倍型结果一致。而笔者的研究也发现,KJ437166 种群可能是来源于茶叶寄主,应该鉴定为茶黑刺粉虱B1 单倍型,而不是橘黑刺粉虱A. spiniferus,陈瑶等[15]也得出了类似的结论。然而,Uesugi 等[14]又表示,从系统发育树的结果来看,不同的单倍型(群)分支间可能已经达到不同种的级别。在该研究中,贵州湄潭(GH81)、独山(08、09)和山东泰安(EM13)的样本属于B1 单倍型(群),贵州贵阳样本(HCNK-1D1、HCNK-1D2)则属于B2 单倍型(群),这是除茶黑刺粉虱重庆永川种群之外在其他地区初次发现B2 单倍型,2 种单倍型(群)分化明显(遗传距离8.2%~8.7%),但是否已经达到不同种的水平,尚需要结合生态学或更多的序列数据进一步阐明。
4 结 论
研究利用DNA 条形码技术成功获取了来源于贵州独山、贵阳、湄潭以及山东泰安茶园的6 个茶黑 刺 粉 虱 样 本 的mt DNACOI基 因DNA 条 形 码 特定的标准区域序列片段,登录号分别为ON534214、ON534215、ON534319、ON534320、ON548917、ON548918,通过序列比对发现,茶黑刺粉虱个体间的遗传距离(0~8.7%)远小于与外群刺桐粉虱遗传距离(28.9%~29.8%),证明DNA 条形码技术应用有效;系统发育树结果显示,来源于不同地区的茶黑刺粉虱明显分化为2 大分支,分别为贵州湄潭、独山和山东泰安分支以及贵州贵阳分支,其结果可支持茶黑刺粉虱的单系性结论,同时2 大分支分化明显,尚可能存在隐存种。
猜你喜欢
今日农业(2022年15期)2022-09-20
黑龙江大学自然科学学报(2022年1期)2022-03-29
计算机系统应用(2021年10期)2022-01-06
少年文艺·开心阅读作文(2021年8期)2021-09-05
上海师范大学学报·自然科学版(2018年3期)2018-05-14
中学生物学(2018年8期)2018-03-01
数学大王·中高年级(2018年12期)2018-01-10
学苑创造·B版(2017年10期)2017-12-21
学生天地·初中(2014年1期)2014-02-17
疯狂英语·口语版(2013年1期)2013-01-31
