
中国荻的群体遗传多样性研究
2022-09-30 01:50王宇飞赵禹贺李珊珊徐晓龙
广东蚕业 2022年8期
王宇飞 赵禹贺 李珊珊 刘 帅 徐晓龙
(1.德州学院 山东德州 253023;2.中国农业大学 北京 100193)
当前煤炭、石油、天然气等不可再生资源被过度消耗,世界面临着能源短缺问题,石油、天然气等资源消耗过程中产生的一氧化碳、二氧化碳等气体,又造成了环境污染和温室效应等问题。人类社会要走向可持续发展,必须寻求可再生的清洁能源。开发能源作物就是解决能源问题和环境问题的一个有效途径[1]。第一代能源作物包括玉米、高粱、甘蔗、大豆、油菜等粮食作物或经济作物,但这些作物大多是一年生,需年年耕地播种,大量灌溉和施肥,投入能量相对较大。另外这些经济作物往往与粮食作物争夺耕地,带来了一系列负面影响[2]。基于此,以芒草、柳枝稷、杨树、柳树等为代表的第二代能源作物应运而生,其特点是生物量高、管理成本低、适应恶劣的气候条件和贫瘠的土壤条件[3-4]。其中,芒属(MiscanthusAndersson s. l.)植物具有生物量高、耐受力强、多年生等优点,可用于与煤混合燃烧发电、发酵制造乙醇和环境修复等,在生物能源领域受到广泛关注[5-6]。荻(Miscanthus sacchariflorus)是芒属重要的能源物种,它主要分布在中国北方和中部地区,以及日本、韩国、朝鲜和俄罗斯,生长于山坡、路边、平原岗地和河岸湿地。荻作为一种C4植物,具有生物量高、抗寒、抗旱等优良特性。荻还是现在广泛推广的三倍体杂交种M.×giganteus的亲本,因此具有很重要的应用价值[7]。
遗传多样性反映了个体之间的遗传差异,正确评价现有栽培植物及其野生近缘物种的遗传多样性,是现代植物育种和改良的重要依据。目前针对芒属的群体遗传学研究大多针对芒属的另外几个物种,例如芒、南荻等[8-10],而针对荻的群体遗传学研究相对较少,而且大多研究没有运用群体的取样策略,不能在群体水平探讨遗传多样性的相关问题[11]。本研究利用微卫星分子标记分析中国分布的荻进行遗传多样性研究,旨在回答以下几个问题:(1)中国分布的荻是否存在种下等级的遗传分化;(2)其遗传多样性水平如何,遗传分化水平多高;(3)是否存在地理分布格局,地理因素是否是造成遗传分化的主要原因。
1 材料与方法
1.1 材料的采集
样品采集自中国东北三省、内蒙古自治区、山西、陕西、甘肃、河南、河北、山东等地。样品采集严格按照群体的取样策略进行,即选择足够大的群体,在群体中随机选择有代表性的个体。由于荻存在克隆繁殖的现象,为了避免取到相同遗传个体的不同克隆分株,应尽量做到每个个体样品间的距离大于10 m,每个群体取样个体数目大多为20个左右,不同群体间的距离大于200 km。选取植株的新鲜叶片置于密封袋中,用硅胶干燥,待完全干燥后放进-20 ℃冰箱保存。共选取在地理分布上有代表性的29个群体的541个个体,具体信息见表1。
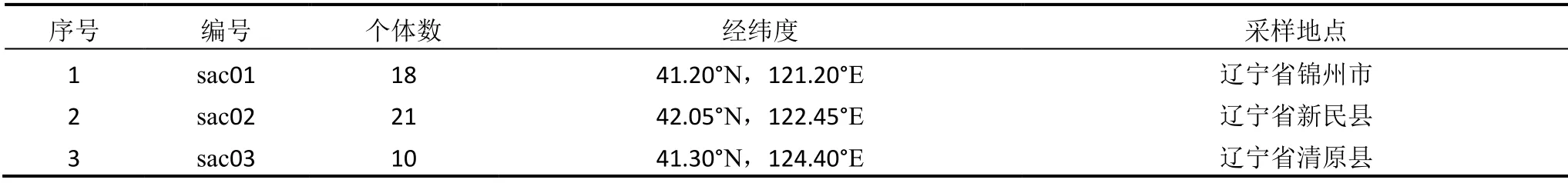
表1 荻的天然群体信息表
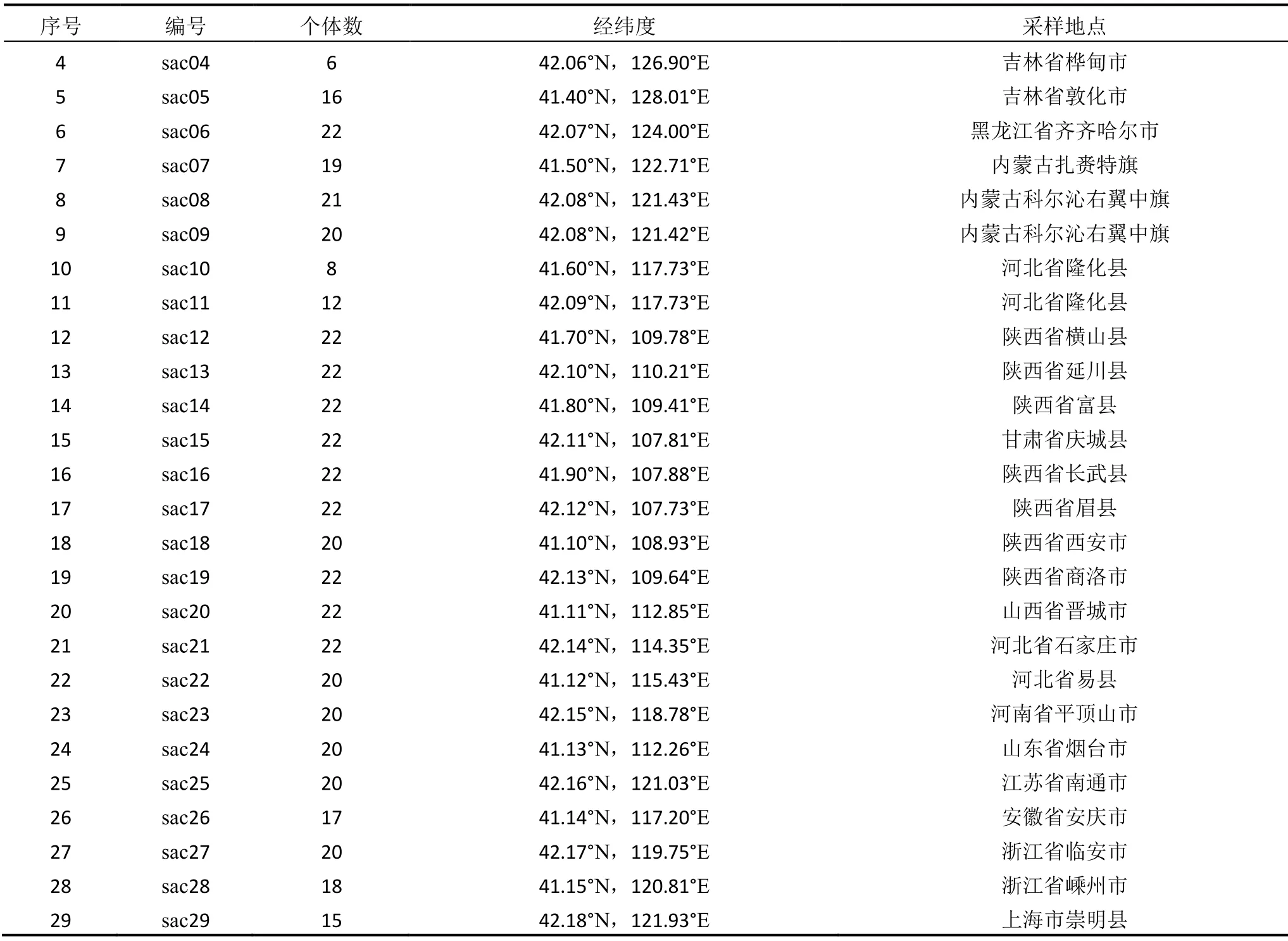
续表1 荻的天然群体信息表
1.2 DNA提取和微卫星标记的筛选
1.2.1 DNA的提取
干燥叶片用CTAB法提取DNA。DNA用1.5%琼脂糖凝胶电泳检测,于-80 ℃保存。
1.2.2 SSR引物选择及PCR扩增
在已发表的相关文章中[12-13]选取扩增条带清晰单一的20对引物,在引物5’端增加一个荧光标记(FAM蓝色,HEX绿色,TAMRA红色)。对所有的参试个体进行PCR扩增。将1 μL PCR产物、0.6 μL GS500LIZ内标和8.4 μL Hi-Di甲酰胺均匀混合95 ℃变性5 min后在ABI 3730测序仪上检测,根据GeneMapper 4.0显示的电泳峰图读取样品的等位基因。
1.3 数据分析
对每个群体每个位点,计算如下遗传多样性参数:位点平均等位基因数(mean number of alleles per locus, NA)、平均观察杂合度(mean observed heterozygosity, Ho)、哈迪-温伯格平衡下的平均期望杂合度(mean expects heterozygosity,He)、多态信息含量(polymorphic information content,PIC)。NA、Ho、He参数的计算均用PopGene 1.31软件包完成。PIC的计算用软件Powmarker 3.0完成。
在进行Structure分析时,选择混合模型对所有群体进行分析,burn-in为50万次,MCMC循环100万次,先设定K值范围为1~15,输入不同的随机数,每个K重复计算10次。用GeneAlex 6.2中的Mantel test来判断遗传变异的分布是否符合距离隔离模型,绘制各个类群的遗传距离与地理距离之间的散点图,判断遗传距离与地理距离之间的相关性。并用SPSS软件分析群体的遗传多样性水平(He)与纬度之间的相关性。
2 结果分析
2.1 标记多态性分析
如表2所示,20个位点整体上表现出较高水平的多样性,平均每个位点等位基因数为15,其中多样性较高的m11、M04、m18、D09、d02位点分别有25、40、23、26、29个等位基因。平均期望杂合度为0.523,其中最小的A39为0.038,最大的d02为0.801。观察杂合度普遍比期望杂合度略低,平均表现杂合度为0.389。平均多态信息含量为0.602,范围在0.015~0.930之间。零等位基因出现的频率是0.033,其中零等位基因频率较高的位点有d21、d02,其零等位基因的频率分别是0.061和0.060,其余位点零等位基因频率均小于0.06。
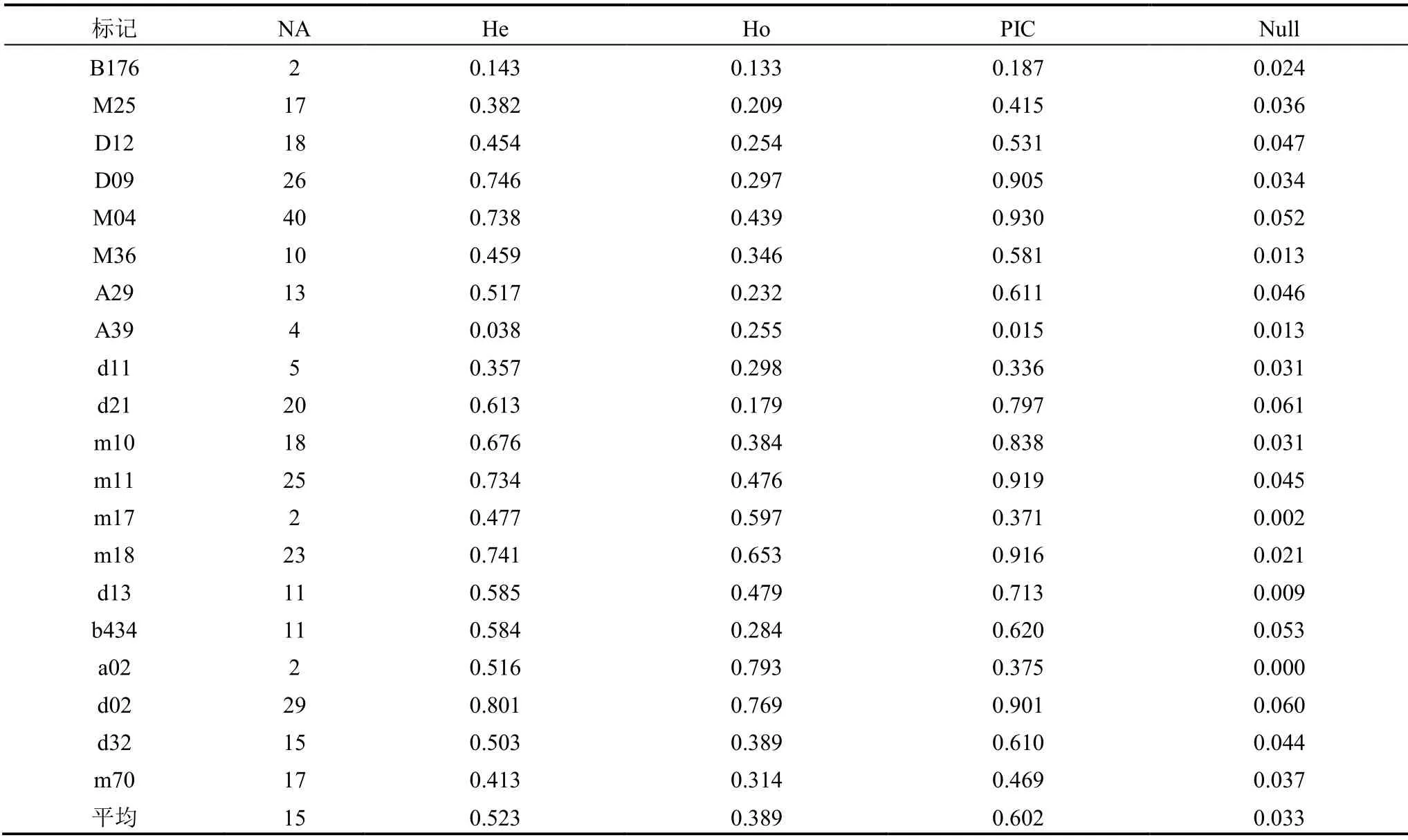
表2 标记多态性分析表
2.2 遗传结构分析
Structure分析中,最大似然值在K=1~15的范围内持续升高,没有明确的最大值。为了确定最优K值,计算了△K值,发现在K=3时,△K出现明显的峰值,这说明参试群体可以大致分为3部分。图1为K=3时的Structure分析图,横向为参试的荻的个体,竖直方向为Q值分布,每份个体竖向线条最长色条的颜色决定了该份材料所属类群。中国荻的类群可以划分为3个亚群,亚群1包括9个居群,共153个个体,主要分布在辽宁、吉林、黑龙江和内蒙古自治区;亚群2包括12个居群,共238个个体,主要分布在河北、陕西、甘肃、山西;亚群3包括8个居群,共150个个体,主要分布在河南、山东、江苏、安徽、浙江。类群的划分表现出强烈的地域性,3个亚群在地理分布上大致分属于东北地区、华北地区和华中地区。
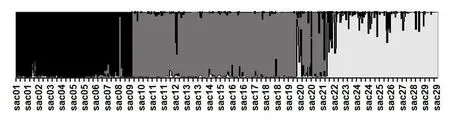
图1 Structure分析结果
2.3 群体的遗传多样性水平和遗传分化分析
如表3所示,无论是在群体水平还是在类群水平,荻的遗传多样性水平普遍较高,而且差异不大,群体水平上的期望杂合度He普遍低于类群水平。总体类群水平上He为0.71,Ho为0.53。3个亚类群中,华北地区荻的遗传多样性水平最高,He为0.58,其次是东北和华北地区,He分别为0.56和0.53。3个类群内部各个群体之间遗传分化水平最高的是东北地区的荻,遗传分化系数Fst是0.17,其次是华北和华中地区,Fst分别是0.09、0.04。Mantel test检验结果表明,荻的遗传距离与地理距离呈显著的正相关(r=0.32,P<0.01),这表明遗传结构符合地理隔离模型,即地理距离近的群体其遗传距离也较近,推测地理隔离因素是造成群体间分化的因素(见图2)。遗传多样性参数He与纬度的相关性分析结果显示(见图3),二者表现出显著的负相关关系(r=-0.24,P<0.01),即遗传多样性随纬度的升高而减小,推测低纬度地区更加稳定而适宜的气候条件为荻的遗传分化提供了良好的环境。
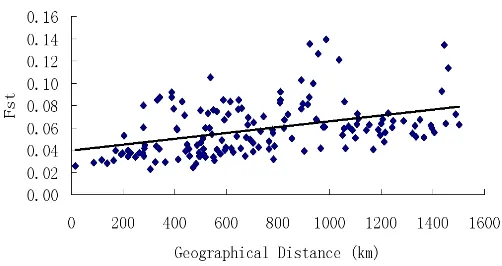
图2 Mental test分析图
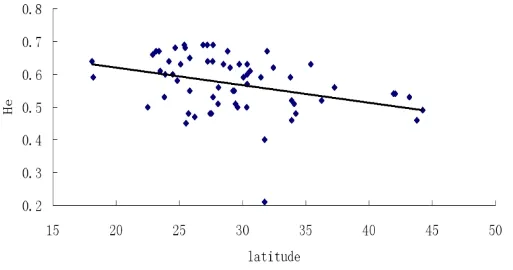
图3 遗传多样性参数He与纬度的相关性分析图
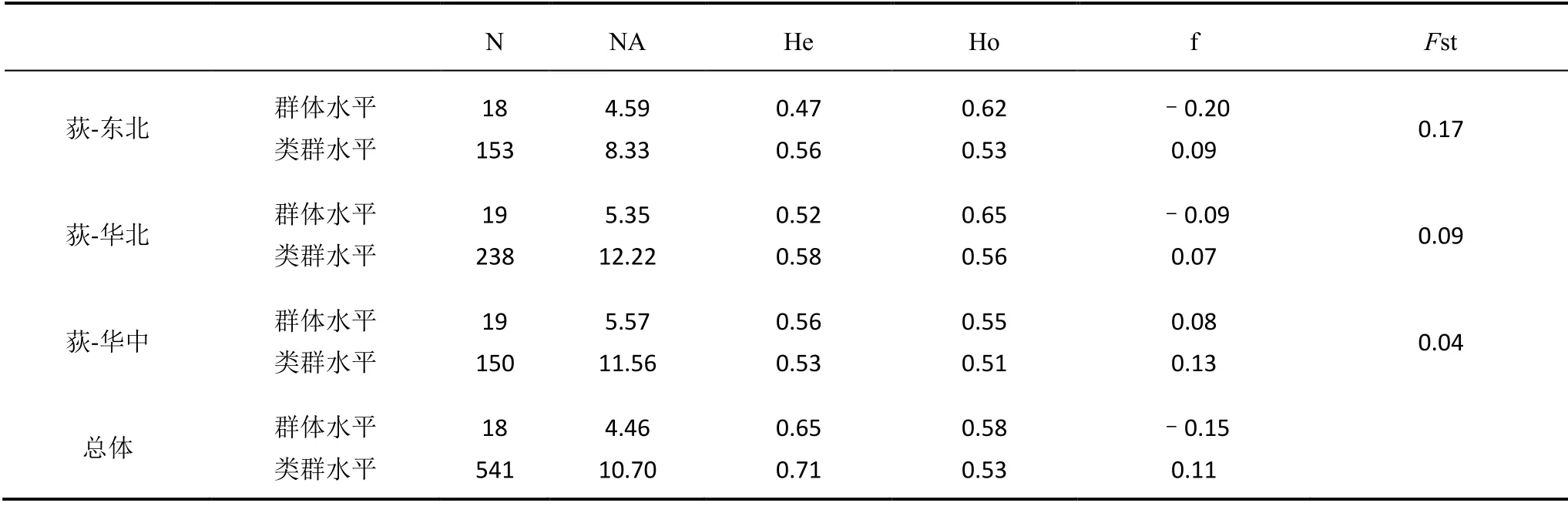
表3 群体的遗传多样性水平和遗传差异分析表
3 讨论
由于芒属植物作为能源植物的重要应用价值,近年来针对芒属植物的遗传多样性研究越来越多,早期针对芒属的群体遗传学研究多局限在某个局部地区,实验方法有AFLP、SSR、ITs等。近年来的群体遗传学研究的取样范围逐渐扩大,涵盖整个中国、日本和韩国等地区,随着测序技术和基因组学的飞速发展,简化基因组测序(RADseq)等组学手段被广泛采用[8-11,14]。但多数研究是针对芒属的芒(M. sinensis),针对荻的研究相对较少。Von Wü hlisch等(1994)用同工酶技术对芒、荻和三倍体杂交种M.×giganteu做遗传多样性分析,大部分同工酶都有很高的变异量,其中变异量最高的等位酶有9种变异式样[15]。Greef等(1997)用6个AFLP标记调查了2个荻个体、11个芒个体和31个M.×giganteus的遗传多样性水平和亲缘关系,在他的研究中荻的遗传多样性较低[16]。Clark等(2019)曾用RAD-seq技术对荻和相近物种进行群体遗传学分析,结果显示中国北方荻的遗传多样性指标SNP diversity(D)为0.139[11]。本研究针对20个SSR位点研究了中国荻29个群体的遗传多样性水平,群体水平遗传多样性参数(He)为0.65。造成各项研究之间荻的遗传多样性水平差异的原因,一方面来自取样范围的不同,本研究取样范围更广,基本上能覆盖其分布范围;另一方面来自分子标记分辨率的不同,SSR标记具有高多变性的特点。
SSR应用于禾本科其他重要作物野生种的群体遗传多样性研究中,水稻的野生近缘种Oryza rufipogon的He为0.787[17],大麦的野生近缘种Hordeum spontaneum的He为0.74[18],玉米的野生近缘种Zea maysssp.mexicana和Zea maysssp.parviglumis的He分别为0.85和0.89[19]。与同属于禾本科的其他重要作物的野生种相比,荻的遗传多样性水平略低,但仍旧保留了较高水平的遗传多样性和较高的群体间分化程度。中国荻的天然群体是育种及遗传改良最重要的基因库,可以为驯化出高产能、高抗性、高耐受性的芒草品种提供丰富的遗传资源。
本研究中荻不同群体之间表现出较高水平的遗传分化,而且呈现明显的地理分布格局,即所有群体可以大致分为3个亚群,地理分布上分别对应中国的东北地区、华北地区和华中地区。Mental test分析表明,地理距离与遗传分化之间呈现显著的相关性,进一步说明了地理距离是促进遗传分化的重要因素。芒属另一个重要物种芒(M.sinensis)的遗传分化也表现出类似的地理分布格局。Zhao等(2013)用SSR标记对中国范围内的芒进行遗传多样性和遗传结构分析,结果显示所有个体被划分成5个组分,5个组分的分布区存在明显重叠[9]。Clark等(2014)用RADseq和SSR对中国、韩国和日本的芒进行研究,所有个体被分为6个组分,其中4个组分来自亚洲大陆(中国东南部、秦岭-长江、四川盆地、韩国和中国北方),另外2个组分分布在日本的北部和南部[20]。Li等(2019)用SSR标记对中国芒属4个物种进行群体遗传学研究,其中芒可以划分为北方和南方两部分[14]。Clark等(2019)在对荻和相近物种进行群体遗传学分析中,荻表现出明显的多倍化现象,二倍体的荻根据地理分布可划分为两部分,一部分是中国东北地区、韩国和俄罗斯,另一部分分布在中国北方地区[11],与本研究中东北和华北两个亚群的地理分布大体一致。但该研究的取样范围没有包含河南、江苏、安徽、浙江一带的荻,因此不能与本研究第三个亚群(华中地区的荻)相互印证。芒属物种的遗传分化表现出明显的地理分布格局,这可能与其风媒传粉和较强的抗逆性有关。
猜你喜欢
中国免疫学杂志(2022年17期)2022-11-15
分子催化(2022年1期)2022-11-02
中国农业科学(2022年16期)2022-09-19
中国医药科学(2022年5期)2022-05-05
清华金融评论(2022年4期)2022-04-13
作物学报(2022年3期)2022-01-22
科学导报(2020年25期)2020-04-28
植物保护(2019年2期)2019-07-23
电脑知识与技术(2018年19期)2018-11-01
古代文明(2014年1期)2014-02-23
