
木荷林窗微环境特征及幼苗更新研究
2022-10-17 09:56刘海英蒋仲龙姚任图孙碧霞葛晓改李正才周本智
西南林业大学学报 2022年5期
刘海英 蒋仲龙 姚任图 孙碧霞 王 刚 葛晓改 李正才 王 增 周本智
(1. 浙江省公益林和国有林场管理总站,浙江 杭州 310020;2. 国家林草局钱江源森林生态系统定位观测研究站,中国林业科学研究院亚热带林业研究所,浙江 杭州 311400)
森林生态系统易受自然或人为因素的干扰而形成林窗,由于林窗大小和森林类型的不同,使得林窗内微环境发生改变,形成了与周围区域不同的小气候条件,并影响着林窗内的生物、化学、物理和水文过程[1]。林窗大小影响着林窗内太阳辐射、土壤温度、土壤湿度和风速[1-2]。此外,林窗的形成在一定程度上影响着林分结构,并在维持林窗内植物多样性方面起着重要作用。
林窗内的小气候可促进林窗内种子萌发,林下草本植物和木本植物的生长速度也有所提高,然而,林窗内生物和非生物因素的变化取决于林窗的大小和所在林窗内的位置[3]。通过林窗进入的光线会进一步穿透林窗边缘森林的林下植被,形成林窗内和林下小气候的差异[4]。一般来说,森林林窗的干扰会影响可利用资源,如光照、水分和养分,而这些资源与幼苗的成活和生长密切相关[2,5]。Sefidi等[5]的研究表明,通过在山毛榉(Fagus orientalis)混交林中建立中小型林窗来模拟自然干扰状态,可以为山毛榉的更新提供适宜的条件。林窗的大小、形状的动态变化与林窗边缘树木的生长、死亡以及林窗内幼苗的生长有密切关系[6]。
冰雪灾害是森林极端干扰的最重要形式之一,在森林群落演替和森林生态系统发展方面发挥着重要作用[7]。在我国亚热带地区,冰雪灾后的林窗研究很少受到关注,鉴于此,本研究以2008年冰雪灾害形成的木荷林(Schima superba)林窗为研究对象,开展冰雪灾害后林窗微环境及其对幼苗更新和生长的研究,为评价极端气候干扰对林窗微环境的影响,以及冰雪灾害后形成的林窗微环境与幼苗更新之间关系提供理论基础,也为研究灾后木荷群落更新及恢复提供参考依据。
1 研究区概况
研究区位于浙江省江山市江郎山(118°33′45″E,28°32′00″N),气候温暖湿润,四季分明,年平均 降 水 量1 650~2 200 mm,相 对 湿 度85%~95%,无霜期250~290 d,年均气温14 ℃,土壤为黄壤,植被覆盖率为85%。区内自然植被类型为常绿阔叶林、针阔混交林、针叶林和灌木林等,优势树种包括木荷、马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)、毛 竹(Phyllostachys pubescens)。江郎山植被垂直分布具有一定的规律性,山体中下部为常绿阔叶林(其中木荷林分布较广),中上部为常绿落叶阔叶混交林,岩壁为苔藓植被、岩生草甸和次生灌丛。本研究的木荷林主要位于海拔400~600 m。
2 研究方法
2.1 样地设置与调查方法
在受冰雪灾害影响的木荷林内,通过前期踏查和所设立的调查林窗,最终选择了海拔、坡度、坡向和土壤等立地条件类型大体一致的44个具有代表性的林窗,保证试验研究具有一定的可比性,并测量其宽度和长度,根据林窗大小和分布情况,将林窗划分为小林窗( < 50 m2)、中林窗(50~100 m2)和大林窗(≥100 m2)3种类型,其中小林窗26个,中林窗16个,大林窗2个。
在林窗内及距林窗10 m处的非林窗林分内设置5个4 m × 4 m的样方,调查各样方内树种、乔木数、胸径(DBH)、树高(H)等特征,选择3个2 m × 2 m小样方和4个1 m × 1 m小样方进行灌木和草本调查,林窗内的更新苗木划分幼苗(H<1 m)和幼树(H> 1 m,DBH > 5 cm)2种[8],调查记录样地内物种的数量、树高和基茎等因子。
2.2 因子测定
在各林窗内,分别林窗和非林窗测定光照、温度、相对湿度和土壤温度等环境因子。于2010年7—9月每月选择晴好天气3~5 d,从8:00至18:00每隔1 h,用TES-1336照度计沿水平梯度测量,即在林窗中心区(冠林窗)、林窗近心区(冠林窗与扩展林窗的过渡区)、林窗边缘区(扩展林窗与非林窗的过渡区)[8-9]及非林窗(林下)分别测量光照强度(为了便于比较,林窗内光照日变化取各林窗中心区光照强度);在3个林窗中心区及林下,用TES-1360A温湿度表在距离地表1 m处测气温和空气相对湿度。
2.3 土壤样品采集和测定
在每个林窗的南北轴及东西轴上,分别间隔2~3 m各设立5个土壤采集样点,清除地表枯落物后,采用内径为5 cm的土钻,分层采集0~5、5~10、10~20、20~40 cm土壤样品,同一林窗同一土层的土壤样品均匀混合。同时在离开该林窗10 m处的非林窗的郁闭林分中,清除去除地表枯落物后,采用内径为5 cm的土钻,分层采集0~5、5~10、10~20、20~40 cm土壤样品,同一土层的土壤样品均匀混合,带回实验室风干,供土壤分析和测定。
土壤pH值采用电位测定法测定;土壤有机质含量采用重铬酸钾容量法测定;土壤全氮凯氏定氮法测定。土壤有效氮采用碱解扩散法测定。土壤全磷和有效磷采用钼锑抗比色法方法测定[10]。
如某风险带来损失的概率相对低,且最大损失也不会对企业造成重大影响,就可主动选择风险自留,这将是比其他风险处理技术更经济有效的处理方式。
2.4 分析方法
采用Excel 2007进行数据处理,所有统计分析使用SPSS 16.0软件包进行。采用Canoco分析法分析幼苗密度、幼苗生长特性与环境因子之间的相关关系。
3 结果和分析
3.1 林窗微环境特征
由图1可知,不同大小林窗和非林窗的光照强度日变化均为单峰型,早晚光照较弱,中午11:00—13:00前后达到峰值。林窗内光照强度随林窗大小显著增加,同一时间点不同大小林窗光照强度表现为大林窗 > 中林窗 > 小林窗 > 非林窗,大林窗和中等林窗最大光照强度分别比小林窗高117.3%和39.1%,不同大小林窗和非林窗光照强度的时间变化差异显著(F=5.88,P=0.002),林窗面积越大,光照强度越强,大林窗的日变化最显著,最大为3 663 lx。
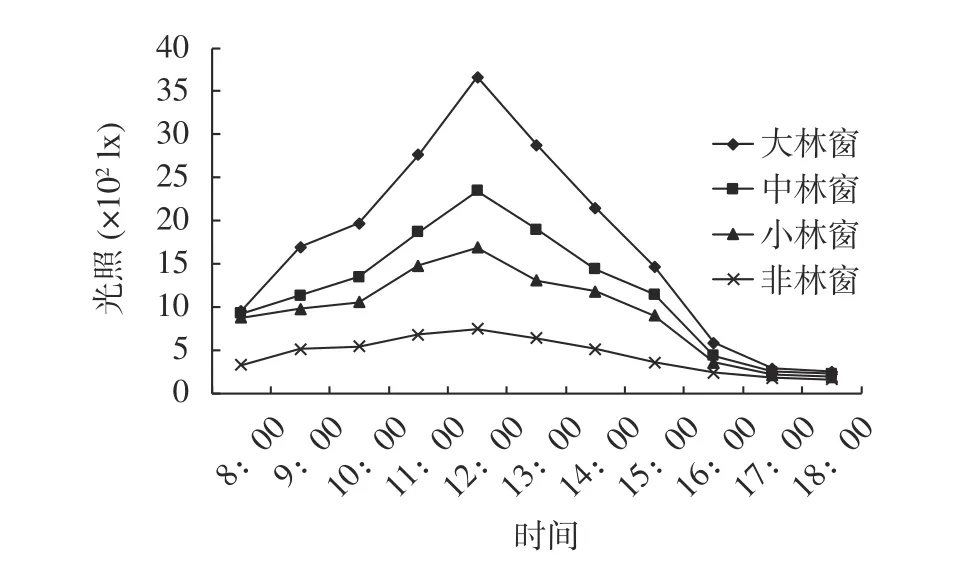
图 1 不同大小木荷林窗与非林窗的光照强度日均变化Fig. 1 The diurnal variation of average illumination intensity of different size forest gaps and non-forest gap in S. superba stand
不同大小林窗和非林窗的气温日变化均也呈单峰变化,在约12:00之前逐渐上升,并达到峰值(图2),林窗内温度随林窗的增大而增大,但是差异均不显著(F=1.095,P=0.362)。小、中、大林窗和非林窗的日变化幅度分别为7.8、9.6、10.8 ℃和6.1 ℃,日最高气温与光照强度具有相同的时间动态,林窗昼夜温差大于非林窗昼夜温差。
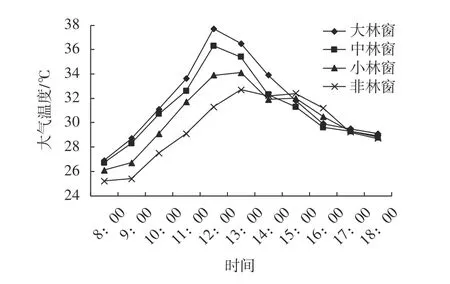
图 2 木荷不同大小的林窗和非林窗的气温日变化Fig. 2 The diurnal variation of air temperature in different size forest gaps and non-forest gaps in S. superba stand
林窗和非林窗空气相对湿度的日变化均表现为早晚较高,中午较低,变化曲线呈“V”型,最小值出现在13:00左右(图3)。中、小林窗相对空气湿度大于大林窗,但是差异均不显著(F=0.915,P=0.442)。林窗空气相对湿度与温度、光照强度密切相关,温度越低,相对湿度越大,光照强度越大,相对湿度越小。
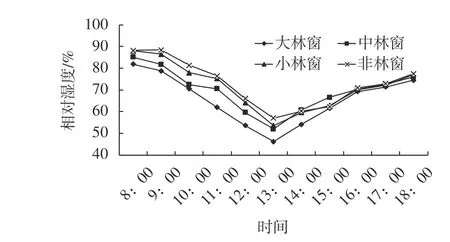
图 3 木荷不同大小林窗和非林窗空气相对湿度的日变化Fig. 3 The diurnal variation of air humidity in different size forest gaps and non-forest gap in S. superba stand
3.2 林窗和非林窗土壤养分特征
除土壤全磷含量外,林窗和非林窗土壤养分含量在垂直方向上均存在显著差异(图4)。林窗和非林窗土壤有机质、全氮、水解氮和速效磷的变化规律相似,均随土壤深度的增加而减小。土壤全磷含量随土层深度的增加而降低,但各土层间差异不显著;土壤pH值随土层深度的增加而逐渐增加,但差异不显著。
林窗与非林窗0~5 cm土壤养分含量除全磷外均存在显著差异,非林窗土壤养分含量均高于林窗(图4)。0~5 cm土层土壤有机质、全氮、水解氮和速效磷的变化,林窗分别比非林窗低42.3%、21.1%、16.9%和24.4%。林窗内各土层pH值均高于非林窗,但仅表层土壤pH值存在显著差异(F=18.85,P=0.012)。
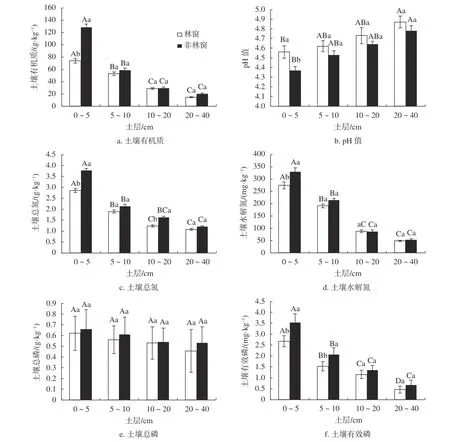
图 4 木荷林窗和非林窗主要土壤养分和pH值动态Fig. 4 Dynamics of main soil nutrients and pH value of forest gap and non-forest gap in S. superba stand
3.3 林窗和非林窗的幼苗更新特征
不同大小林窗与非林窗的更新特征存在显著差异(P< 0.05)(表1)。木荷的更新密度在中等林窗最高,其次是小林窗,大林窗最低,中等林窗幼苗更新密度分别比小林窗、大林窗和非林窗高41.20%、57.15%和49.66%。
林窗内更新幼树树高和基径都显著高于非林窗更新幼树树高和基径,而不同大小林窗之间更新幼树树高和基径无显著差异。林窗内更新幼苗基径显著高于非林窗更新幼苗基径,中等林窗内更新幼苗高度显著高于非林窗幼苗高度,其他大小林窗与非林窗无显著(表1)。不同大小林窗幼苗高度,中等林窗幼苗高度为39.65 cm,显著高于小林窗幼苗高度(F=4.893,P=0.007)和大林窗幼苗高度(F=5.203,P=0.004)。
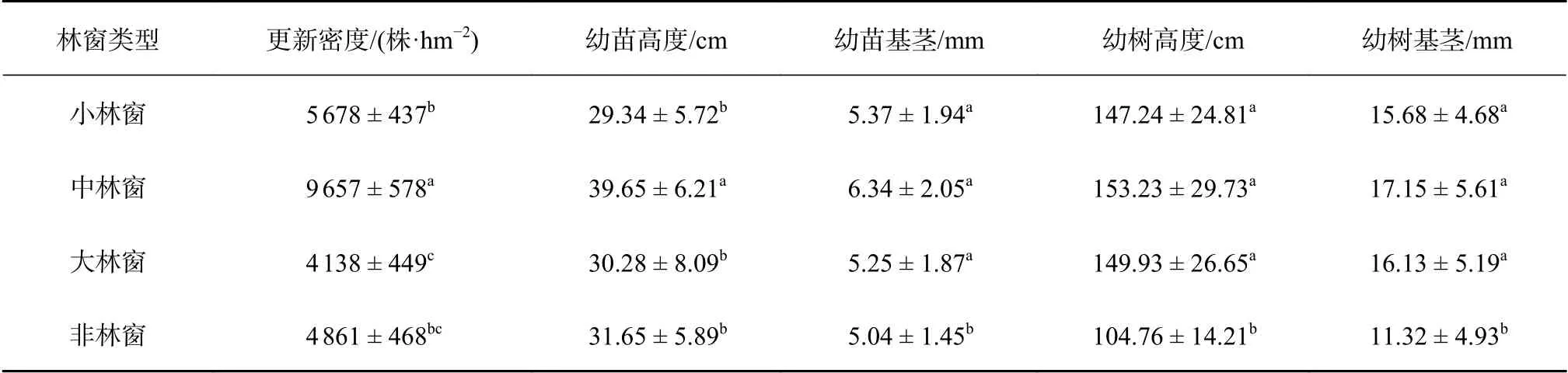
表 1 林窗和非林窗木荷幼苗和幼树的更新密度和生长特性Table 1 The regeneration density and growth character of S. superba seedlings and saplings in gaps and non-forest gaps
3.4 微环境对林窗和非林窗更新的影响
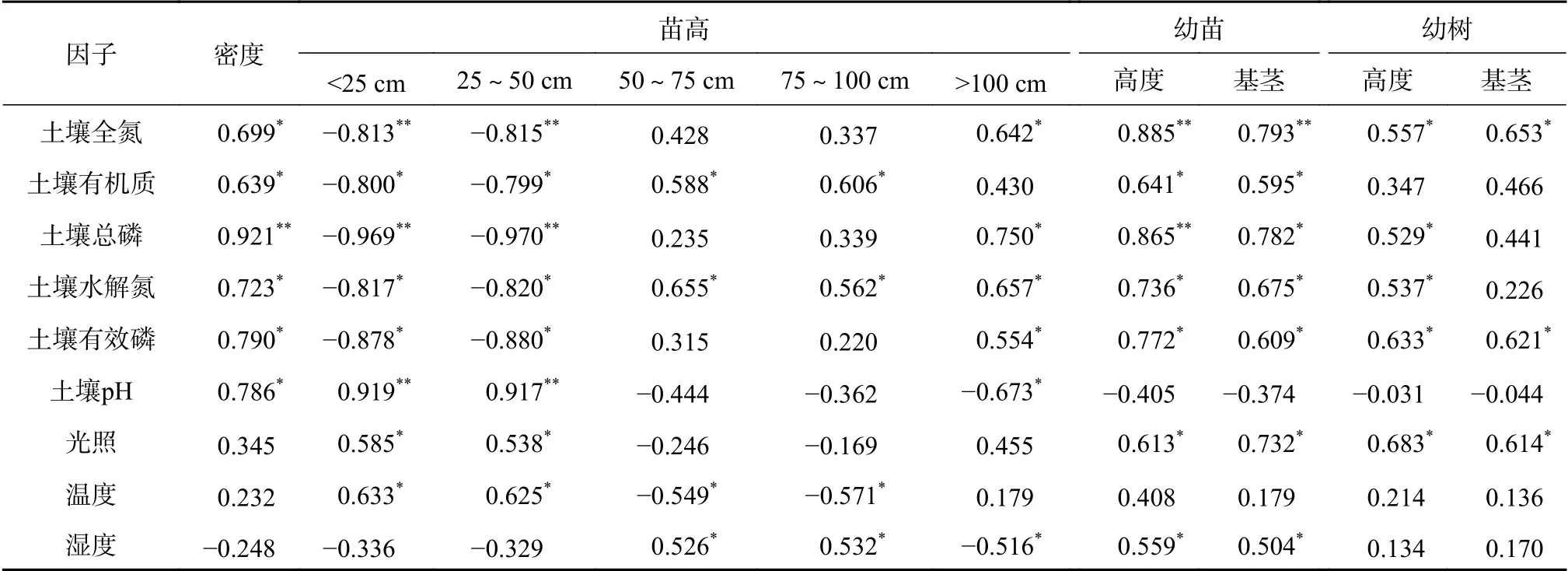
表 2 苗木生长特征与土壤因子的相关性Table 2 Relationship between seedling growth characteristics and soil factors
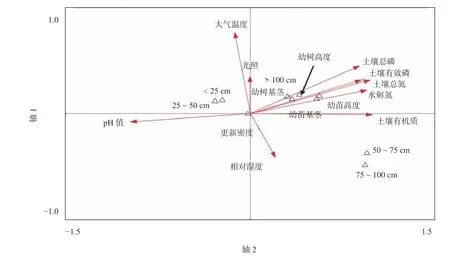
图 5 微环境与林窗内外幼苗更新之间的RDA二维图Fig. 5 Two-dimensional diagram of RDA between microenvironment to within and outside of forest gaps regeneration
4 结论与讨论
4.1 讨论
4.1.1 林窗微气象环境
根据林窗大小不同,林窗内光照、水分和营养等资源相对于干扰前可能会显著增加[11],林窗内的光照条件优于林内任意地点[12-13]。本研究发现,林窗内的光照强度随着林窗大小的增大而显著增强,大林窗和中等林窗的光照强度分别比小林窗高117.3%和39.1%。,这一研究结果与陈梅等[14]的研究结果一致,表明不同林窗大小和形状的光照强度及其分布是不同的。林窗引起光照强度的增加,从而促进了林窗内幼苗和幼树的生长。Beaudet等[15]的研究表明,冰雪灾害后,林分水平的平均光合光子通量密度在地面上0.3~4.0 m范围内增加了4~5倍。林窗内的光照、土壤和空气温度以及土壤湿度的空间异质性取决于微地形的变化、更新树木和边缘树木的生长导致的遮荫效果等[4]。林窗形成后,林窗内及林窗周围的小气候立即发生变化,光照强度大幅增加,从而提高了空气温度,降低了空气湿度[16]。本研究中,大林窗和中林窗的光照水平相似,而小林窗的光照强度更接近于林下,中林窗的土壤温度和光合有效辐射透过率较高,土壤湿度较低,而小林窗的光照强度小于大林窗,这说明土壤质地的局部变化会影响土壤湿度和温度[17]。
朱教君等[18]的研究表明,在不同大小的林窗内,最高温度出现的时间不同,大林窗比小林窗更早,他们还观察到,与小林窗相比,中等林窗的土壤湿度更低。本研究也表明,林窗内的温度随林窗的大小而逐渐增加,小、中和大林窗温度日较差分别为7.8、9.6 ℃和10.8 ℃(图2),这表明林窗大小对植物更新微环境具有重要影响。
4.1.2 林窗大小与幼苗更新
与森林林下植被相比,林窗内形成小气候可以促进种子萌发,提高草本植物和木本物种的生长速率[3]。在本研究中,中等林窗的幼苗更新密度最大,说明中等林窗湿度和光照条件等环境因子更加适合幼苗更新,研究结果与刘庆[19]的研究结果一致,即:在大小不同的林窗中,幼苗的高度和基径大小如下:中 > 小 > 大 > 林下,表明林窗内草本植物的光照比小林窗更充足[20]。此外,林窗大小对种群更新密度等变量也有影响。在本研究中,中等林窗种群密度比小林窗和大林窗种群密度高41.20%~57.15%,说明林窗面积是幼苗更新的关键[9]。相反,Sefidi等[5]则认为林窗大小与树苗频度、多样性之间没有显著的相关性。
Kern等[21]的研究也表明,中等林窗(20 m直径)到大林窗(30 m和46 m直径)的幼树比小林窗的幼树要少。相比之下,闫兴富等[22]的研究表明,小林窗中心区域早期苗高和基茎高于林缘。鲜骏仁等[23]的研究表明,亚高山天然针叶林林窗与林冠之间冷杉幼苗平均高度存在显著差异,而平均基径和生物量没有显著差异。在本研究中,幼树的基径也有类似的结果。因此,中等林窗可能更加适宜森林更新,幼苗生长更快[19],说明中等林窗优越的水分和温度条件可以促进养分循环,促进幼苗和幼树的生长。
4.1.3 林窗土壤养分动态
林窗对土壤养分的影响是一个复杂的过程,不同研究区域的林窗可能与原生土壤的肥力条件、凋落物分解速率和根系生物量的返还有关[24-25]。本研究表明,与非林窗土壤相比,林窗内土壤有机质含量下降了42.33%,全氮下降了21.14%,水解性氮下降了16.91%,有效磷含量下降了24.36%,可能的原因是森林凋落物是土壤有机质的主要来源,林窗形成后,林窗内凋落物大幅减少,从而导致林窗内土壤有机质含量降低,而土壤有机质是土壤有效养分的重要决定因素,土壤有机质的减少导致土壤有效养分的减少[26]。
林窗增加了光照时间和光照强度,林窗大小通过根系和微生物活动的竞争而间接改变了微环境[3]。在本研究中,林窗大小与土壤养分有显著的相关关系(图5)。不同大小林窗的幼苗和幼树,无论是树高还是基径,都受到土壤养分的显著影响。
林窗大小影响土壤微生物的活性,小林窗内土壤微生物的活性受到周围大乔木的影响,减弱了有效光照和降雨,阻碍了土壤温度、湿度等条件的改善;而大林窗内白天温度太高,并且昼夜温差大,也不利于土壤微生物活性的增加,因此,导致林窗内土壤微生物的活性随着林窗面积的增大呈现先升后降的趋势[27]。林窗大小影响土壤酶的活性,许建新[28]的研究表明,土壤脲酶、土壤磷酸酶和土壤过氧化氢酶的活性均随林窗面积的增大而下降。虽然林窗形成初期光照的增加有利于提高土壤酶活性,但随着时间推移,大林窗内太阳辐射直达地表,气温、土温都较高,林窗越大,土壤水分蒸发越激烈,因此,土壤酶的活性也随之下降。
综合上述分析,相对于大林窗和小林窗,中等林窗温度、湿度和光照条件三者均比较优越,所以导致中等林窗内的有机质分解比大林窗和小林窗内更快[17],释放更多的养分,从而更加有效地促进幼树和幼苗的生长,因此,本研究认为中林窗(50~100 m2)可能是最优的林窗,研究结果与刘庆等[8]的研究基本一致。
4.2 结论
林窗大小显著影响林窗内的光照强度、温度、湿度和土壤养分,进一步影响林窗的幼苗更新和生长。中等林窗的幼苗高度、幼树地径和幼树高度均高于小林窗和大林窗,表明中等林窗在湿度和光照强度方面更适合幼苗幼树的生长,林窗大小与土壤养分之间存在显著的交互作用。本项研究对2008年南方冰雪灾害后林窗土壤生产力的维持和提高以及灾后森林的快速恢复具有一定的指导意义。
猜你喜欢
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
小学生优秀作文(低年级)(2021年10期)2021-11-24
花卉(2021年2期)2021-01-21
林业与环境科学(2020年3期)2020-07-21
河北果树(2020年1期)2020-02-09
现代园艺(2017年11期)2017-06-28
作文大王·低年级(2017年1期)2017-02-16
土壤与作物(2015年3期)2015-12-08
爆笑show(2015年3期)2015-05-08
