
西沙群岛冷泉区中层鱼类群落结构初探
2022-10-25 08:33江艳娥陈作志徐姗楠朱江峰于文明
南方水产科学 2022年5期
田 翰 ,江艳娥, ,张 俊, ,陈作志, ,徐姗楠, ,朱江峰,于文明
1. 上海海洋大学 海洋科学学院,上海 201306
2. 中国水产科学研究院南海水产研究所/农业农村部外海渔业可持续利用重点实验室,广东 广州 510300
3. 南方海洋科学与工程广东省实验室 (广州),广东 广州 511458
海底冷泉通常指富含硫化氢、甲烷以及其他碳氢化合物以流体形式在海底的喷溢,一般与周边正常底层水体有着相同或相近的温度;深海冷泉活动区域能够为部分化能微生物提供丰富的养分,形成深海冷泉生态系统,一般是深海海底生命极度活跃的地方,被喻为“深海沙漠中的绿洲”[1-2]。目前,从水深几十米的浅海到数千米的深渊区均有冷泉生物群落的分布,国内外对其理化性质和底栖生物群落的研究逐步深入[3-5]。Macdonald等[6]在墨西哥湾冷泉区的表层海洋监测到与冷泉渗漏同源的碳氢化合物,推测冷泉能影响中上层海洋的鱼类群落。琼东南盆地位于南海北部陆缘海南岛的东南部(108°E—111°E、15°N—19°N),是我国南海海域冷泉最丰富的地区之一,不同发育区冷泉流体通量、时空演化、地形地貌、流体地球化学、沉积物特征、生态系统、水合物赋存等方面有着较大差异[7-8]。
中层鱼类 (Mesopelagic fish) 是指栖息于大陆斜坡和大洋中层 (200~1 000 m) 的小型鱼类群,广泛分布于世界各大洋海域,是海洋中现存生物量最大的脊椎动物类群[9-10]。近年研究显示,全球中层鱼类资源量高达19.5×109t,该数值仍因观测技术不完善存在被低估的可能[11]。昼夜垂直迁移 (Diel vertical migration, DVM) 发生在部分种类中层鱼类中,是一种以日周期进行的昼沉夜浮垂直方向的长距离运动,中层鱼类通过昼夜垂直迁移为深海带入碳源,加速海洋固碳进程[12-13];同时中层鱼类在海洋生态系统中扮演着重要角色,起到连接浮游动物和高营养级掠食生物的生态作用[14]。白天中层鱼类垂直分布在200 m以深海域的不同水层,其中深海散射层 (Deep Scattering Layers, DSL) 是中层鱼类分布最集中的区域;夜间部分种类中层鱼类经过垂直迁移后集中分布在近表层海域 (<150 m),其他种类继续停留在深海[15-16]。
由于研究技术和采样工具的局限,国外对冷泉区鱼类群落的研究极少[17-18]。迄今国内关于冷泉区域中层鱼类组成和多样性的研究尚未见报道,如冷泉生态系统对其区域内鱼类群落的影响,冷泉区域内外鱼类群落结构组成差异等尚不清楚。本研究采用中层拖网对西沙群岛冷泉生态系统内外区域的中层鱼类资源进行调查,探讨了冷泉区内外中层鱼类种类组成、优势种和物种多样性的特征及其差异,旨在为认识冷泉生态系统中层鱼类种类组成、物种多样性特征提供参考资料。
1 材料与方法
1.1 站位设置和鱼类样品采集
于2021年5月25—28日对位于琼东南盆地的西沙群岛冷泉区内外海域中层鱼类进行调查,冷泉内、外区域各设3个临近的采样站位,内部站位为 A1、A2、A3,经纬度范围为 110°31'E—110°33'E、16°40'N—16°43'N;外部站位为 B1、B2、B3,经纬度范围为 112°14'E—112°29'E、18°16'N—18°24'N(图1)。鉴于部分中层鱼类种类有昼夜垂直迁移的习性[9-10],在白天的中层鱼类调查未设置表层水域采样站点;故此次中层鱼类资源调查设置站位分为白天深层水域、夜间表层水域和夜间深层水域。其中,表层采样深度范围为50~75 m,深层采样深度范围为400~600 m,依据声学映像并结合实际海况展开。采样站位详细信息见表1。用ArcGis 10.2软件制图,水深数据来自通用大洋水深制图 (GEBCO)。
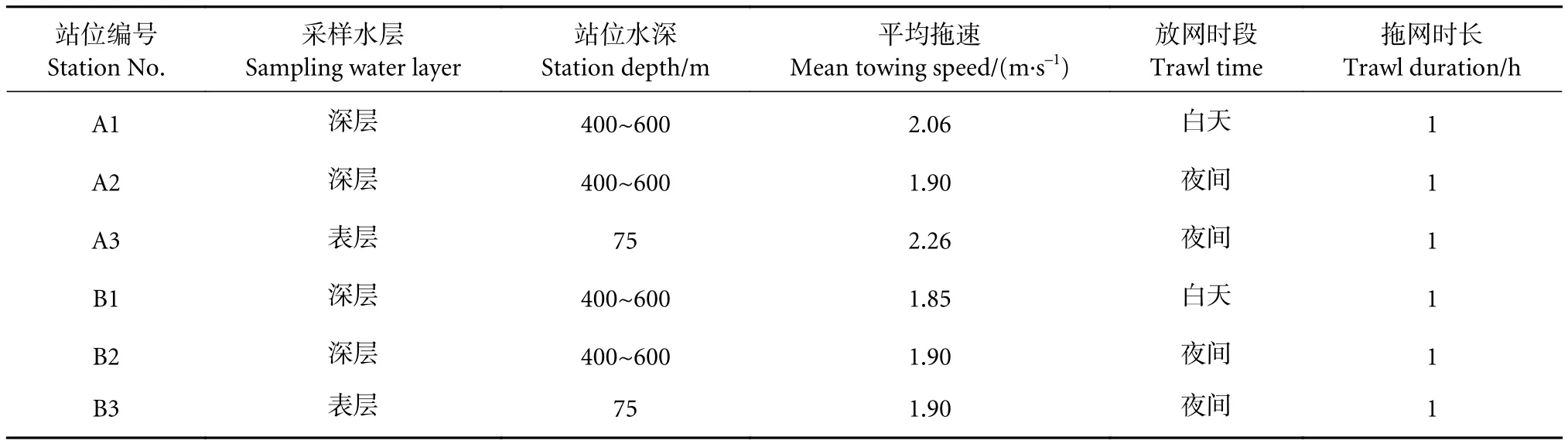
表1 南海西沙冷泉内外区域中层鱼类拖网采样站位信息Table 1 Information of sampling stations for mesopelagic trawl in sea area inside and outside cold seep in Xisha Islands of South China Sea

图1 南海西部西沙海域中层鱼类采样站位图Fig. 1 Sampling station for mesopelagic fish in Xisha Islands of western South China Sea
调查依托中国水产科学研究院南海水产研究所“南锋”号渔业科考船,总吨位1 537,吃水4.8 m。中层鱼类生物样品由专业中层拖网采集,规格为:主尺寸136.80 m×50.85 m (网口周长×网衣全长),上、下纲均为25 m,正方形4片网,网口高度6.20~10.50 m,平均为8.60 m,网囊网目1 cm,拖速 1.9~2.26 m·s—1。起网后,在甲板上将网囊及网衣上的渔获物收集后,在船上生物实验室对样品进行鉴定、计数、测量和记录,包括总质量、种类、尾数、体长 (1 mm)、体质量 (0.1 g)。依据中层鱼类形态特征尽可能精确鉴定到最低分类阶元,部分样本经95%乙醇保存留样。
1.2 研究内容与方法
中层鱼类在海洋中的垂直分布与时间、水深关系密切[14-16],本研究在分析琼东南海盆地冷泉内外区域中层鱼类种类组成和多样性等渔业生物学特征差异时,主要比较冷泉内外区域相同时段和深度站位 (A1对B1,A2对B2,A3对B3)。南海西沙海域冷泉内外区域种类组成差异采用Primer软件对各站位中层鱼类进行等级聚类分析 (Cluster analysis, CLUSTER) 和Jaccard种类相似性指数[19-20]分析;同时采用分类多样性方法 (平均分类差异指数、分类差异变异指数) 分析物种间的差异及其亲缘关系[21]。鱼类群落中的优势种采用相对重要性指数 (Index of relative importance, IRI) 估算[22]。基于传统多样性方法中的Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数计算群落多样性[23-25];并以分类多样性方法为传统多样性方法的补充,在讨论中比较了冷泉内外区域中层鱼类的群落组成差异[26]。采用主坐标分析 (Principal co-ordinates analysis, PCoA) 进行群落排序分析[27];采用Past软件进行百分比相似性分析 (SIMPER) 找出对各分组贡献较大的物种组成,二者共同描述南海西沙冷泉区域鱼类群落组成差异[28]。
Jaccard种类相似性指数(I):
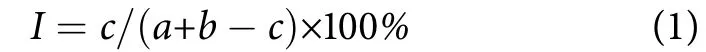
式中:a为冷泉外站点的鱼类种类数;b为冷泉内站点的鱼类种类数;c为冷泉内外共有种数。当0%≤I≤25%时为极不相似,25%≤I≤50%时为中等不相似,50%≤I≤75%时为中等相似,75%≤I≤100%时为极相似[20]。
平均分类差异指数 (Δ+):

分类差异变异指数 (Λ+):
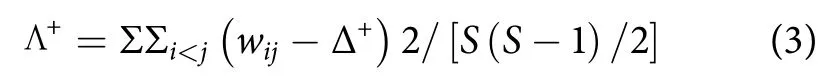
式中:Δ+表示群落全部物种间路径长度的理论平均值;Λ+表示Δ+的偏离程度;wij为第i和第j个种类在分类系统树中的路径长度;S为种类数[21]。
相对重要性指数 (IRI) 和优势度百分比 (IRI%):
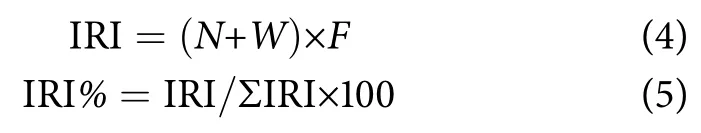
式中:N、W分别表示某一种类的尾数占总尾数和重量占总重量的百分比 (%);F表示某一种类出现的站位数占调查总站位数的百分比 (%)。划分标准为:IRI>1 000,该鱼种为优势种;1 000>IRI>100,该鱼种为重要种;100>IRI>10,该鱼种为一般种;IRI<1,该鱼种为少有种[29]。
Shannon-Wiener多样性指数 (H'):

式中:Pi =ni/Ni,表示第i鱼种所占总个体数的比例。
Pielou均匀度指数 (J'):

式中:H'为Shannon-Wiener多样性指数;S为种类数。
Margalef丰富度指数 (D):
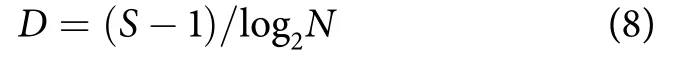
式中:S和N分别为鱼类样本中的总种类数和总个体数。
2 结果
2.1 中层鱼类种类组成
2.1.1 南海西沙冷泉区域中层鱼类种类组成
冷泉区共计采集中层鱼类84种,隶属于11目30科54属,科、种信息见附录A (http://dx.doi.org/10.12131/20210370的资源附件)。其中,冷泉区鱼类种类组成以灯笼鱼目和巨口鱼目鱼类为主(图2),各站位其他目鱼类占比均小于35%。
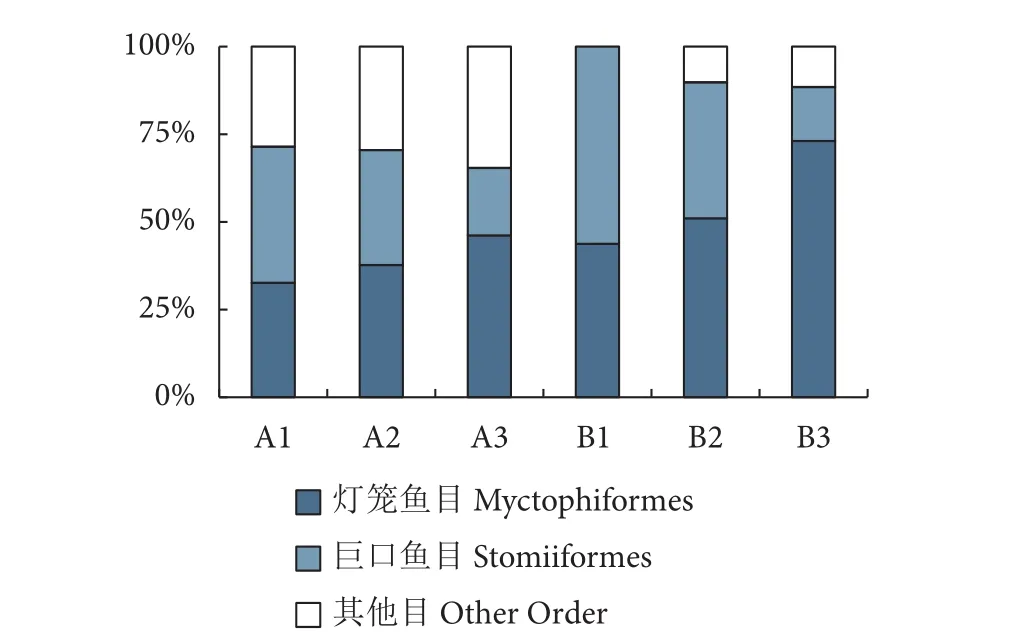
图2 南海西沙冷泉内外区域站位中层鱼类各目种类数占比Fig. 2 Percentage of number of mesopelagic fish species by Order in sea area inside and outside cold seep in Xisha Islands of South China Sea
2.1.2 南海西沙冷泉区域内外种类组成差异
冷泉区采集中层鱼类11目30科54属84种,冷泉外采集鱼类6目13科32属60种 (附录A,详见http://dx.doi.org/10.12131/20210370的资源附件);与冷泉区域对比可知,冷泉外最主要的类群同样是灯笼鱼目和巨口鱼目鱼类,各站位中这两目的鱼类占比介于88%~100% (图2),均高于冷泉内。冷泉区出现的鳗鲡目和鮟鱇目鱼类在冷泉外区域未曾采集到;冷泉内部采集到鲈形目鱼类9种,冷泉外仅采集到该目鱼类1种,即鳞首方头鲳(Cubiceps squamiceps)。在白天的调查中,冷泉内外深层水域 (A1、B1) 分别采集鱼类49和16种。在夜间样本采集中,冷泉内外表层水域 (A3、B3) 所采集的中层鱼类种类均为26种;冷泉内外深层水域 (A2、B2) 采集的中层鱼类种类分别为61和49种。基于种类数和物种丰度的聚类分析共同表明,冷泉内部区域白天深层 (A1) 和夜间深层 (A2)相似度较高;同时,冷泉内外区域之间的夜间表层(A3、B3) 表现出较高的相似度 (图3)。Jaccard相似性系数表明,冷泉内外区域之间在白天深层(A1、B1) 表现为极不相似 (18.18%),在夜间深层(A2、B2) 同样为极不相似 (30.95%),而在夜间表层 (A3、B3) 为中等不相似 (40.54%,表2)。

图3 南海西沙冷泉内外区域中层鱼类站位聚类分析Fig. 3 CLUSTER results of mesopelagic fish species in sea area inside and outside cold seep in Xisha Islands of South China Sea
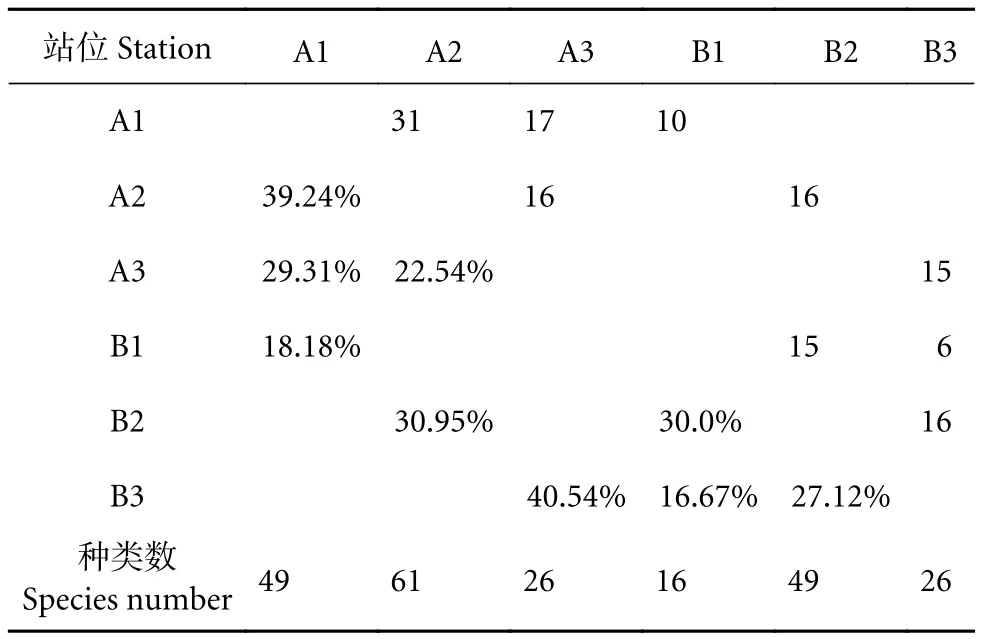
表2 南海西沙冷泉内外区域站位种类组成差异Table 2 Difference of species composition in each station in sea area inside and outside cold seep in Xisha Islands of South China Sea
2.1.3 分类多样性
冷泉内部各站位Δ+范围为70.16~70.74;外部各站位Δ+范围为55.82~64.20;冷泉内部各站位鱼类群落Δ+均显著大于冷泉外部各站位 (t-检验,P<0.01)。冷泉内区域鱼类群落的Λ+范围为275.45~334.20;外部Λ+范围为453.31~531.4;冷泉内区域各站位鱼类群落的Λ+则均显著小于冷泉外区域 (t-检验,P<0.01,表 3)。
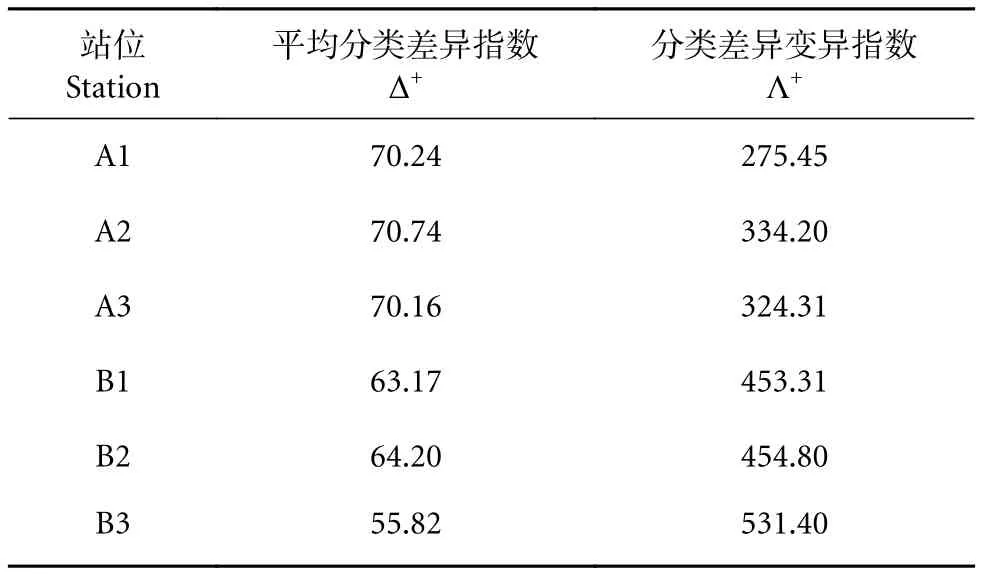
表3 南海西沙冷泉内外区域不同站位鱼类平均分类多样性指数及分类差异变异指数Table 3 Average taxonomic distinctness (Δ+) and variation in taxonomic distinctness (Λ+) in each station in sea area inside and outside cold seep in Xisha Islands of South China Sea
2.2 中层鱼类群落特征
2.2.1 优势种
选取IRI大于1 000的种类作为西沙冷泉内外区域中层鱼类群落的优势种 (表4)。其中,冷泉内部白天深层 (A1) 优势种共计5种,优势度百分比之和为47.93%;夜间深层 (A2) 共计2种 (23.21%),夜间表层 (A3) 共计5种 (75.41%);冷泉外部白天深层 (B1)共计6种 (79.27%),夜间深层 (B2) 共计3种 (29.82%),夜间表层 (B3) 共计5种 (71.72%)。在冷泉内部,白天深层 (A1) 和夜间深层 (A2) 有2种共有优势种;冷泉外部白天深层 (B1) 和夜间深层 (B2) 有1种共有优势种;白天深层 (B1) 和夜间表层 (B3) 有1种共有优势种。冷泉内外区域各相同时间及水层站位之间,白天深层 (A1、B1) 无共有优势种;夜间深层 (A2、B2) 共有优势种1种;夜间表层 (A3、B3) 共有优势种2种。

表4 南海西沙冷泉内外区域各站位中层鱼类优势种Table 4 Dominant species of mesopelagic fish in each station in sea area inside and outside cold seep in Xisha Islands of South China Sea
2.2.2 群落多样性
冷泉内部中层鱼类H' 范围为2.04~3.54,其中夜间表层站位A3的H' 最低 (2.04);冷泉外部的中层鱼类H' 范围为2.23~3.32,其中夜间深层站位B2的H' 最高 (3.32)。冷泉内部J' 范围为0.63~0.89,其中夜间表层站位A3的J' 最低 (0.63);冷泉外部的J' 范围为0.68~0.85,其中夜间表层站位B3的J' 最低 (0.68)。同样,冷泉内部D范围为2.30~7.27,其中夜间表层 (A3)D非常低 (2.30);冷泉外部的D范围为2.58~4.37,其中夜间深层 (B2)D最高 (4.37,表5)。
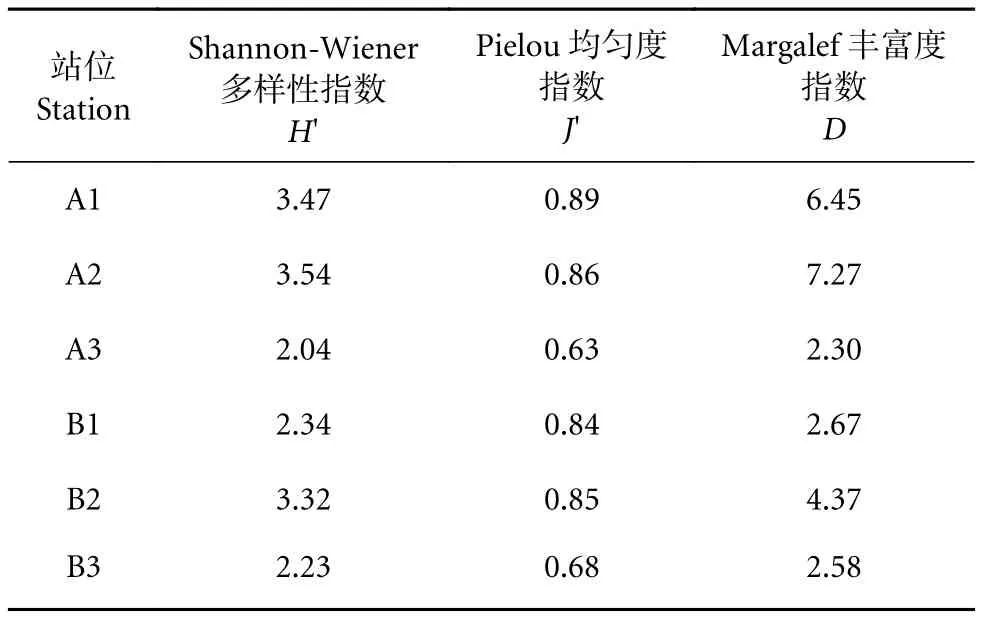
表5 南海西沙冷泉内外区域中层鱼类多样性指数Table 5 Diversity index of mesopelagic fish in sea area inside and outside cold seep in Xisha Islands of South China Sea
2.2.3 南海西沙冷泉内外区域鱼类群落差异
基于种类数的PCoA主坐标轴分析显示,冷泉内部白天深层水域 (A1) 和夜间深层水域(A2) 较为接近,冷泉外部3站位之间距离较远;就冷泉内外区域对比可看出,白天深层 (A1、B1) 和夜间深层(A2、B2) 均差距较大。基于物种丰度的PCoA主坐标轴分析显示,冷泉内部白天深层水域 (A1) 和夜间深层水域 (A2) 较为接近,冷泉外部夜间深层(B2) 和夜间表层 (B3) 之间距离较近;对比冷泉内外区域可知,二者夜间表层站位之间 (A3、B3) 距离非常接近 (图4)。
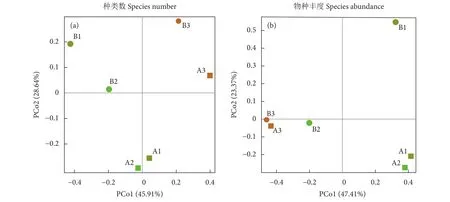
图4 南海西沙冷泉内外区域中层鱼类PCoA主坐标排序分析Fig. 4 PCoA (Principal co-ordinates analysis) ordination of merger of mesopelagic fish in sea area inside and outside cold seep in Xisha Islands of South China Sea
通过SIMPER检验,发现平均不相似度大于2的物种共计8种,累计贡献度达到56.06%,这些物种是造成冷泉内外群落差异的关键种群 (表6)。其中,长鳍虹灯鱼 (Bolinichthys longipes) 和串灯鱼(Vinciguerria nimbara) 的平均不相似度均大于5%,贡献度均大于10%;此外,除喀氏眶灯鱼(Diaphus garmani) 和马来眶灯鱼 (D. malayanus)外,其他种类在冷泉内外的数量百分比均表现为冷泉内部小于冷泉外部。
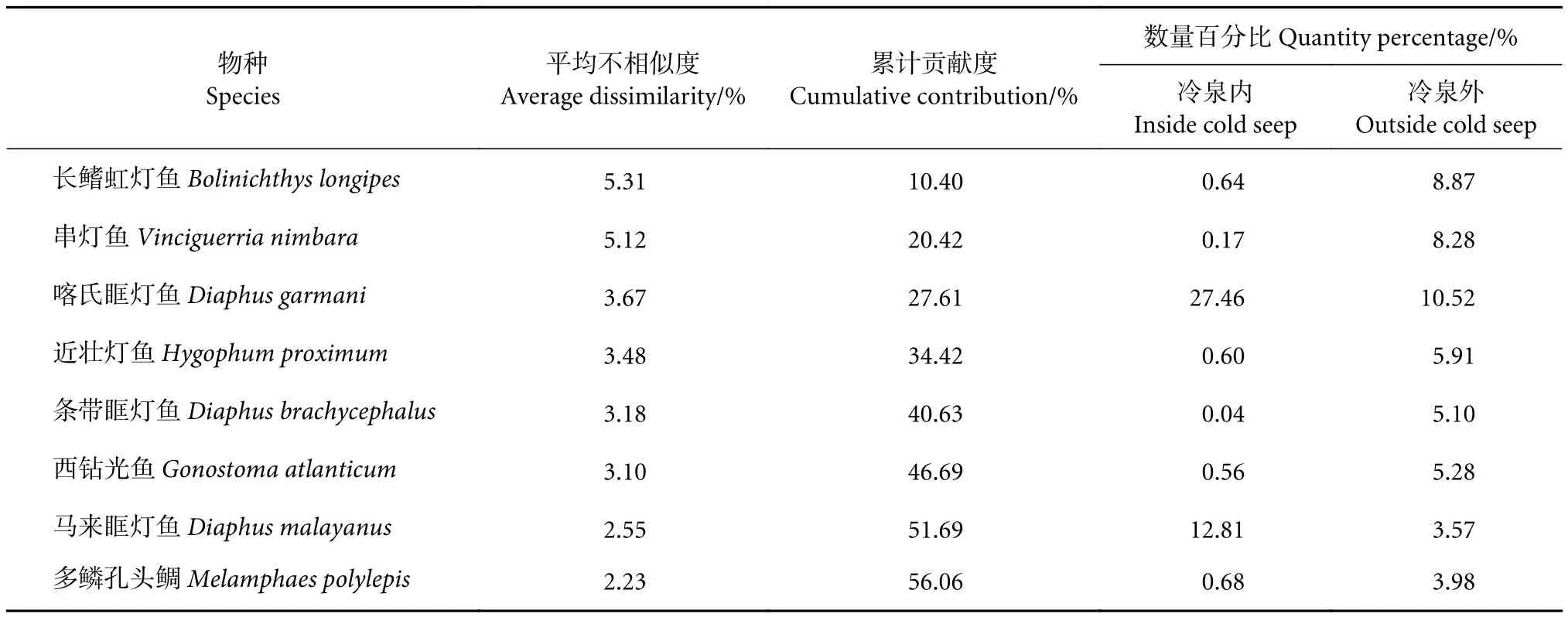
表6 南海西沙冷泉内外区域中层鱼类百分比相似性分析Table 6 Results of similarity percentage analysis (SIMPER) of mesopelagic fish in sea area inside and outside cold seep in Xisha Islands of South China Sea
3 讨论
3.1 南海西沙冷泉内外区域中层鱼类种类组成及差异
调查共采集西沙冷泉区中层鱼类84种,冷泉外部区域60种,冷泉内外区域均以灯笼鱼目和巨口鱼目为主,南海中部海域和南海北部陆坡海域中层鱼类种类组成以灯笼鱼目和巨口鱼目为主,与本研究结论一致[30-32]。然而就冷泉内外区域中层鱼类种类组成对比可见,冷泉内部区域灯笼鱼目和巨口鱼目种类占比相对冷泉外部区域更少,说明冷泉内部鱼类物种丰富度水平相对较高。基于种类数或物种丰度的Cluster聚类分析表明冷泉内外各站位中层鱼类种类的差异显著 (P<0.05),且将冷泉区白天深层和冷泉区夜间深层分为一个群组,同时将冷泉外夜间表层和冷泉内白天表层分为另一个群组,这是由于分组内的种群组成较为相似。Jaccard相似性系数与Cluster聚类分析结果间存在的差异可由计算方法的不同解释,但两者可共同说明,冷泉内外夜间表层的中层鱼类种类组成之间相似性较高,但是冷泉内外白天深层的中层鱼类种类差异较大,导致冷泉内外种类组成差异明显。
此外,冷泉海域内外区域中层鱼类分类多样性存在明显差异,冷泉区域各站位中层鱼类的Δ+显著大于冷泉外区域,冷泉区域各站位的Λ+显著小于冷泉外区域。这表明冷泉内区域鱼类种类间的亲缘关系比冷泉区外更远,但其鱼类群落物种间分类关系的均匀程度比冷泉外区域更均匀。冷泉区域各站位Δ+范围在70.16~70.74,通过与其他海洋生态系统鱼类组成对比可知,西沙群岛冷泉区域站位中层鱼类的Δ+远大于珊瑚岛礁海域 (55.05~58.75)[33-34]以及陆架海域 (65.7~65.8)[35-36],但小于河口区域(72.2~79.9)[37];而西沙群岛冷泉外区域站位中层鱼类Δ+均值 (61.06) 大于珊瑚岛礁海域并小于陆架海域及河口区域,接近近海海湾(62.2~63.5)。冷泉外部Δ+符合史赟荣等[38]认为的河口区域、陆架海域、珊瑚礁海域Δ+依次降低的结论,冷泉内部Δ+则与该结论完全相符,从而进一步反映了冷泉内外种属间存在的差异。
3.2 南海西沙冷泉内外站位中层鱼类群落结构差异
西沙冷泉内外区域中层鱼类群落的优势种组成除前鳍盗目鱼 (Lestidiops jayakari) 外,其余各种均隶属于灯笼鱼目和巨口鱼目。本研究调查发现,西沙群岛冷泉区内部和外部夜间海表灯笼鱼目鱼类的种类及生物量占比均为最高,表明该区域灯笼鱼目鱼类是昼夜垂直迁移的最主要参与者,这一结论与Wang等[39]在南海北部陆坡的调查结果一致。尾明角灯鱼 (Ceratoscopelus warmingii) 同时作为冷泉内外夜间表层的第一优势种,内外优势度百分比分别为26.03%、22.47%,因中层鱼类昼夜垂直迁移的习性,即夜间表层的中层鱼类是进行昼夜垂直迁移的种类[2],笔者推测尾明角灯鱼可能是此次研究海域中层鱼类昼夜垂直迁移研究的模式物种。而斯氏蝰鱼 (Chauliodus sloani) 在冷泉区域的白天深层与夜间深层是优势种,在冷泉外部区域则仅在夜间深层是优势种,3个区域优势种排名及相对重要性百分比依次为第一优势种 (14.79%)、第二优势种(14.64%)、第一优势种 (15.26%);根据研究结果推测产生这一现象的原因是在冷泉外部分布的斯氏蝰鱼在白天下潜至更深的水层。冷泉内外区域各站位的H'、J'、D表现为深层 (A1、A2、B1、B2) 均大于表层 (A3、B3),说明南海西沙海域的中层鱼类在深水层的分布更为丰富。但冷泉内部深层 (A1、A2) 均表现出较高的多样性,冷泉外部仅夜间深层(B2) 表现出较高的多样性,同时,无论白天或夜间,深层区域多样性均为冷泉内部 (A1、A2) 高于冷泉外部 (B1、B2),而表层区域多样性水平则为冷泉外部 (B3) 大于冷泉内部 (A3)。调查发现相较于冷泉外部海域,冷泉内部深层多样性水平高但表层多样性水平低,这说明中层鱼类的水层时间分布受到了冷泉影响,冷泉内部深层适宜更多种类中层鱼类的生存,文章推测冷泉可能影响了中层鱼类的昼夜垂直迁移行为。
基于物种种数和物种丰度的PCoA主坐标轴分析的结果共同反映了冷泉内部白天深层 (A1) 和夜间深层 (A2) 有极大的关联性;而冷泉内外区域的夜间表层 (A3、B3) 虽然在物种丰度上距离较近,但在种类上距离较远,说明冷泉内外中层鱼类种类在夜间迁移至表层摄食的种类差异较大,验证了Δ+在冷泉内部 (70.16) 和外部 (55.82) 夜间表层之间的差异。冷泉区域昼夜深层水域中层鱼类群落组成差异较小但均与表层水域中层鱼类群落组成存在明显差异。若认为冷泉外部海域深层昼夜存在的差异为正常,此差异是由中层鱼类的昼夜垂直迁移引起,则冷泉内部相似性较高的原因可能为冷泉影响了部分种类中层鱼类的昼夜垂直迁移。百分比相似性分析 (SIMPER) 表明,长鳍虹灯鱼、串灯鱼、喀氏眶灯鱼、近壮灯鱼 (Hygophum proximum)、条带眶灯鱼 (D. brachycephalus)、西钻光鱼 (Gonostoma atlanticum)、马来眶灯鱼、多鳞孔头鲷 (Melamphaes polylepis) 均可作为区分冷泉内外区域中层鱼类的关键种群,这些鱼类在冷泉内外区域均有分布,但冷泉内外区域的数目分布差异可以说明冷泉对特定鱼类种群的生存存在影响。本研究结果共同表明,西沙群岛冷泉内外区域中层鱼类的多样性存在一定差异,冷泉影响了中层鱼类群落的种类结构及时空分布;冷泉内深水层更有利于中层鱼类的栖息,初步验证了Macdonald等[6]提出的冷泉对海洋中上层鱼类群落有影响的推测,此外,笔者推测冷泉可能影响了其内中层鱼类的昼夜垂直迁移行为,但需后续研究验证。
迄今西沙群岛冷泉区中层鱼类的群落多样性高的机制尚不清楚,有关昼夜垂直迁移这一特殊行为对群落及结构的影响,以及中层鱼类群落与冷泉生态系统之间的相互影响研究仍在继续。中层鱼类采样区域的选择和站位的设置是首先需要考虑的问题,比如采样季节、站点、时间、水深等。本文仅对西沙群岛冷泉海域内外区域中层鱼类群落组成进行了初步探讨,尚有诸多需要改进之处,尤其需要大幅增加调查站位进行更广泛的采样和比较研究。西沙群岛冷泉海域中层鱼类的群落格局和多样性的维持机制及其如何影响冷泉生态系统的运行仍是非常值得继续研究的科学问题,本研究结果可为冷泉机制研究提供参考资料,同时有助于对西沙群岛冷泉海域海洋渔业资源的开发和利用。
猜你喜欢
传感器世界(2022年4期)2022-11-24
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
建材发展导向(2021年9期)2021-07-16
西夏学(2020年2期)2020-01-24
人大建设(2019年5期)2019-10-08
江苏教育(2019年16期)2019-09-25
山花(2019年8期)2019-08-16
人大建设(2019年3期)2019-07-13
现代企业(2015年6期)2015-02-28
