
长白山苔原带凋落物生态化学计量特征及其对模拟氮沉降的响应
2022-11-26 08:18张慧慧白云玉张英洁靳英华许嘉巍水新利刘丽杰
生态学报 2022年21期
张慧慧,白云玉,张英洁,靳英华,*,许嘉巍,陶 岩,水新利,赵 琛,刘丽杰
1 东北师范大学地理科学学院,长白山地理过程与生态安全教育部重点实验室,长春 130024
2 长白山科学研究院,延边朝鲜族自治州 133613
凋落物是联结土壤和植物的纽带,其淋溶和分解是陆地生态系统养分循环的重要载体[1]。凋落物分解速率的快慢和养分释放的多少,决定了陆地生态系统养分过程的特征,也决定了土壤中有效养分的供应状态,进而影响植物的养分吸收[2]。碳(C)、氮(N)、磷(P)元素的养分循环和转化调节驱动着陆地生态系统地上植被生长、群落结构以及地下土壤过程。在凋落物分解过程中化学计量特征直接影响养分归还的质量和速率, 间接影响植物根系的吸收。凋落物分解过程中,凋落物的化学计量比(C∶N∶P)是重要决定因素,其化学计量比对土壤C、N、P循环具有强烈的影响。植物在生长发育的过程中,需要从所处的生存环境中改变自身的养分循环状态,进而植物本体的生态化学计量特征会受到凋落物及土壤的影响[3]。
凋落物在分解过程中,并不总是释放出养分,它与凋落物的类型和分解阶段有关,也与养分本身特性相关。低品质凋落物在分解的初期经常从环境当中固定养分,含养分量高的凋落物则在较短的时间内释放出养分。由于凋落物普遍缺N,所以N成为限制微生物活动的主要因素,外源性N的输入满足了微生物的N需求,加速了凋落物的分解过程[4]。氮沉降的增加将导致生态系统中N富集,对植物凋落物分解过程中养分元素释放和化学计量比造成影响,进而影响土壤养分的转化与固定;氮沉降还会通过增加生态系统的氮输入,影响群落的化学计量特征,进而对陆地生态系统的结构和功能产生影响。国内关于氮沉降对凋落物生态化学计量特征的研究主要集中在亚热带森林生态系统[5]、西北荒漠生态系统[6]和草原生态系统[7],而氮沉降增加引起的苔原生态系统植被变化带来的不同类型凋落物生态化学计量特征变化的研究未见系统报导。
氮素增加越来越呈现出全球化的趋势[8],即使在人类活动很少的高山苔原区氮素也呈升高趋势[9]。苔原是受到较强氮限制的生态系统[10],土壤无机氮含量低,生态系统易受外源氮输入的影响。氮沉降增加已经导致苔原植被组成、结构和功能发生了显著变化[11—12],成为苔原植被变化的主要驱动因素,并带来凋落物质量和数量的变化。在贫氮的苔原生态系统中,不同凋落物类型的分解特征对氮沉降增加响应是否存在差异以及这种差异对苔原土壤养分和植被的可能影响是亟待解决的科学问题。
长白山苔原是我国乃至欧亚大陆东部独有的高山苔原,根据前人调查植被以灌木苔原为主要类型。由于苔原全年气温低、微生物活动微弱,灌木凋落物含较难分解物质多,因此凋落物分解十分缓慢,土壤养分贫乏[13]。在全球变暖背景下,近30年来长白山岳桦林下的草本植物上侵苔原带,原生灌木苔原分化为灌木苔原、灌草苔原和草本苔原,形成了灌木、灌草混合和草本3种不同类型的凋落物,凋落物数量和质量发生显著改变[14];已有的监测和野外施氮控制实验表明外源氮显著增加是长白山苔原植被变化的主要原因之一,加速了植被从灌木到草本的变化过程[15—16]。长白山高山苔原环境特征与极地苔原相似,长白山苔原变化的研究结论对于极地苔原变化具有较高的指示意义。因此,本研究在长白山苔原带氮沉降增加及灌木植被退化、草本植物增加的背景下,对3种类型植物凋落物进行不同梯度的人工施氮,旨在探讨灌木牛皮杜鹃(Rhododendronaureum)、草本小叶章(Deyeuxiaangustifolia)和灌草牛皮杜鹃-小叶章混生群落凋落物分解后的化学组分和生态化学计量特征对氮沉降增加的响应差异,将有助于加强对草本入侵下苔原带养分元素循环过程的理解,同时为深入研究氮沉降增加背景下苔原带生物地球化学循环提供基础数据。
1 材料与方法
1.1 研究区概况
长白山位于吉林省东南部(127°42′55″—128°16′48″E,41°41′49″—42°25′18″N),为中国和朝鲜两国的界山,是中国东北地区的最高山峰(海拔 2691 m)。苔原带位于长白山火山锥体上部,气候特点为冬季漫长严寒、夏季短暂温凉,春季风大干燥,秋季多雾凉爽[17]。苔原带生长季(6—9月份)日平均气温只有5.9℃。降水丰富,年均降水量可达700—1400 mm,6—9月份降水占全年降水量的60%—70%;积雪时间长达6个月(11月至翌年4月);全年多风,湿度大;雾天和风天日数每年都在260 d以上。
苔原带地表多为碱性粗面岩风化物和少量火山灰,地貌为流水改造的火山锥坡面;土壤为高山苔原土,土层很薄,剖面层次不明显。苔原植被以灌木苔原为主要类型,植物种类较少,极地或高山种属约占80%,优势植物以小灌木为主,牛皮杜鹃和笃斯越橘(Vacciniumuliginosum)是常见的建群种。群落只有灌木层和苔藓、地衣两层片。还有少量草木苔原,以大白花地榆(Sanguisorbastipulata)和小白花地榆(Sanguisorbaparviflora)等植物为代表。近年来,长白山西坡苔原出现了草本植物入侵现象,小叶章、大白花地榆等多度增加,形成草本植物层片,目前苔原存在灌木、灌草和草本3种类型的植被斑块。
1.2 实验设计
2018年生长季末在长白山西坡苔原带手捡当年未分解的新鲜牛皮杜鹃和小叶章凋落物带回实验室,本研究中用纯牛皮杜鹃凋落物代表灌木凋落物,牛皮杜鹃和小叶章凋落物1∶1代表灌草凋落物,小叶章凋落物代表草本凋落物。并分别取纯牛皮杜鹃、纯小叶章及牛皮杜鹃-小叶章混生群落中的土壤,带回实验室进行室内模拟凋落物分解时使用。使用一次性餐盒作为分解盒,并进行标记。分解盒下部均匀铺上土壤,土壤体积约0.1 m×0.1 m×0.1 m,上部平放与土壤对应的凋落物。每个分解盒里放置3袋相同凋落物,每袋凋落物6 g,共计放置3种凋落物108袋。2019年3月使用人工气候箱开始进行为期8个月的室内模拟实验。24小时分为4个时间段,光照级数分别为0、1、3、2,以 EM50数据采集器(Meter,America)监测数据计算的长白山苔原带生长季平均温度作为人工气候箱实验参数,模拟生长季凋落物分解。依据长白山苔原带的氮沉降现状以及2030、2060年预计氮沉降量[17],参照Wright等的试验设计[18],本研究设置3个氮沉降水平,分别为对照(CK, 0 g N m-2a-1)、低氮(LN, 10 g N m-2a-1)、高氮(HN, 20 g N m-2a-1)。在每月月初根据不同氮处理水平,将相应质量的 NH4NO3溶于水中,等量喷洒于分解盒中,无施氮组喷洒等量清水。
1.3 样品收集与指标测定
凋落物收集时间为2019年5月、7月、9月和11月,共收集4次,每次收集27袋(3种凋落物,3个氮水平,3次重复)。将烘干并称重过后的样品用球磨仪粉碎,用以测定各项指标。凋落物总碳、总氮使用碳氮分析仪(Vario EL cube, Germany)进行测定;总磷采用 HNO3消解,经前期处理之后使用TAS- 986原子分光光度计测定(普析公司,中国北京);木质素使用酸性洗涤纤维法测定[19]。
1.4 数据处理
凋落物养分元素剩余百分率[20]:
式中,NR为凋落物养分元素剩余百分率,Mt为凋落物t时刻烘干样品重量(g);M0为凋落叶初始风干样品重量(g);Nt为t时刻凋落叶养分浓度(mg/g),N0为初始养分浓度(mg/g)。
凋落物化学计量比采用元素质量比。
用Excel 2013进行数据整理,用SPSS 20.0进行数据统计分析。用单因素方差和显著分析不同施氮处理对凋落物的影响,双因素方差分析对氮添加和植物类型对凋落物化学组分残留率及化学计量特征进行比较。显著性水平设为P=0.05。并用 Origin Pro 9.1绘制示意图。
2 结果与分析
2.1 3种类型凋落物在不同施氮水平下的化学组分动态变化
在未施氮处理中,3种类型凋落物的C元素在分解过程中都呈释放状态,C残留率的下降速率整体表现为:混合凋落物>小叶章凋落物>牛皮杜鹃凋落物(图1)。分解过程中总体上牛皮杜鹃凋落物的N元素呈富集状态,而混合凋落物和小叶章凋落物的N元素呈释放状态,且混合凋落物的N元素释放最快(图2)。3种类型凋落物的P元素在分解末期均呈释放状态,其中,牛皮杜鹃凋落物和混合凋落物主要在分解初两个月大量释放P元素,小叶章凋落物的P残留率在分解的前六个月持续下降;牛皮杜鹃凋落物P元素释放最快,小叶章凋落物P元素释放最慢(图3)。3种类型凋落物的木质素整体上均呈先累积再降解趋势,但在累积过程中不同类型凋落物之间的变化速率有明显差异;牛皮杜鹃凋落物木质素累积最多,混合凋落物木质素累积略大于小叶章凋落物(图4)。
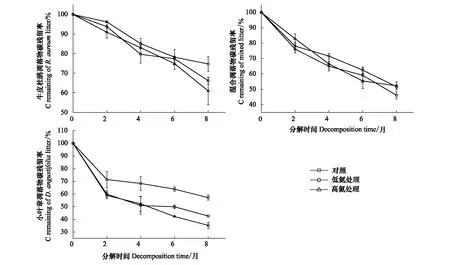
图1 不同施氮处理下3种凋落物分解过程中碳元素残留率的变化
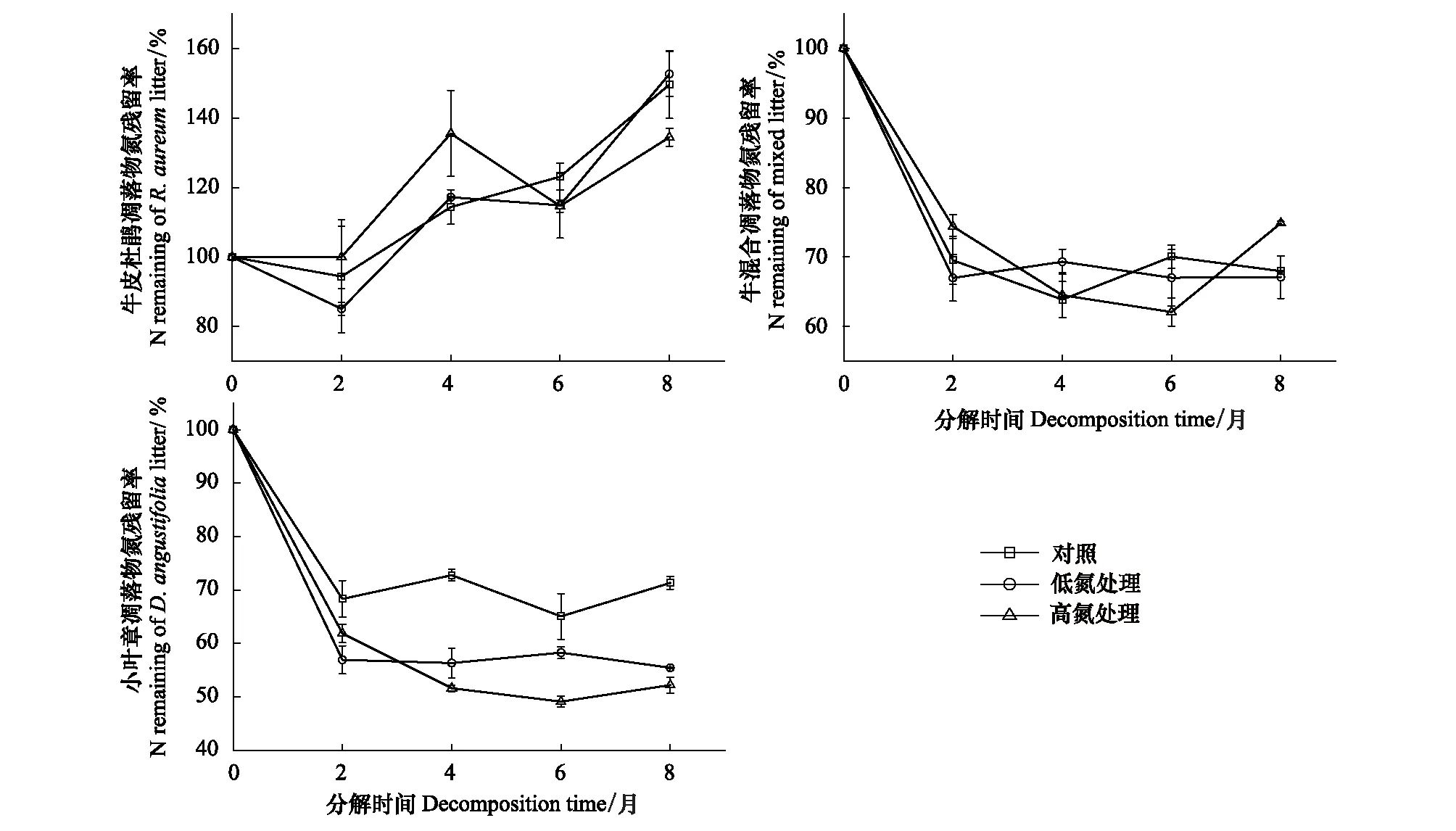
图2 不同施氮处理下3种凋落物分解过程中氮元素残留率的变化
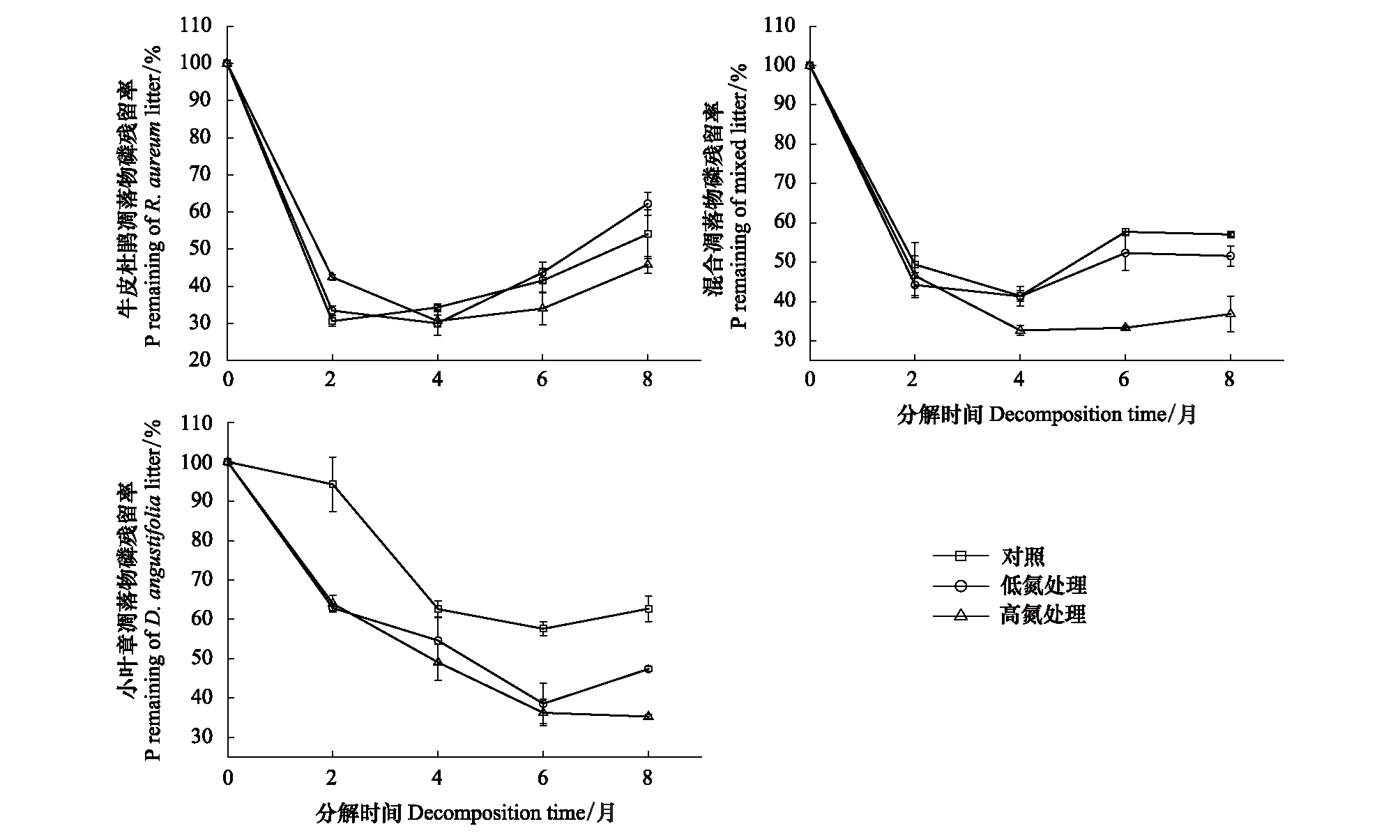
图3 不同施氮处理下3种凋落物分解过程中磷元素残留率的变化
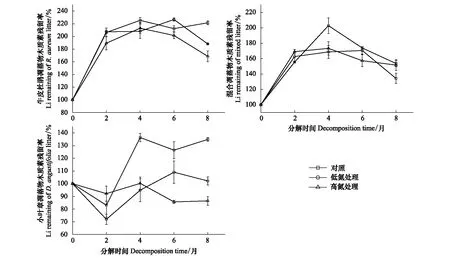
图4 不同施氮处理下3种凋落物分解过程中木质素残留率的变化
如图1所示,在不同施氮梯度处理下,牛皮杜鹃凋落物和小叶章凋落物的C释放均受到促进,且外源氮浓度越高,促进作用越显著,下降速率整体表现为:HN>LN>CK;施氮处理在牛皮杜鹃凋落物分解后期对C释放有着明显促进作用(P<0.05),C残留率在6—8月下降幅度分别是对照组的3.22倍(LN处理)、4.03倍(HN处理)(图1)。混合凋落物在低氮处理下的C释放明显加快(P<0.05),高氮处理对其C元素释放的抑制或促进作用不明显(图1)。施氮处理对小叶章凋落物分解过程中C元素的动态变化及释放程度的影响更为显著(P<0.05)(图1)。
如图2所示,分解结束后,低氮处理对牛皮杜鹃凋落物和混合凋落物的N元素作用效果不明显(P>0.05)。而高氮处理组的牛皮杜鹃凋落物的N元素在分解初始4个月内明显高于对照组(P<0.05),在4—6月快速下降,降幅为20.87%。混合凋落物的N元素在分解前6个月内释放持续受到促进,氮残留率在分解6—8月内有大幅度上升,涨幅约12.79%,故牛皮杜鹃凋落物的N富集及混合凋落物的N释放在分解前期受高浓度氮促进,但在分解后期受到了抑制。施氮处理显著促进了小叶章凋落物N释放(P<0.05),其N元素残留率下降幅度整体表现为:HN>LN>CK。
如图3所示,试验结束时牛皮杜鹃凋落物的P元素均呈释放状态,且在低氮下受到抑制,高氮下受到促进)。施氮处理更有利于混合凋落物和小叶章凋落物的P释放,P元素残留率下降幅度大小比较均为:HN>LN>CK,施氮处理对小叶章凋落物和混合凋落物P元素作用效果主要在表现在分解前期促进其释放。
如图4所示,试验结束时3组牛皮杜鹃凋落物的木质素均发生累积现象,外源氮的输入对木质素的累积起到了抑制作用,大小比较为:CK>LN>HN。低氮处理在凋落物分解前期没有对牛皮杜鹃凋落物的木质素残留率增长造成显著影响,但随着时间的增长其变化速率的差异越来越大。高氮处理在分解0—2月内对木质素的累积起着抑制作用,在6—8月内促进木质素的降解。施氮处理显著抑制了混合凋落物整个分解过程中的木质素累积,在2—4月内对照组的木质素残留率增长幅度达到了47.34%,分别是低氮组(2.98%)和高氮组(4.26%)的15.89倍、11.11倍。施氮处理对小叶章凋落物内木质素变化的作用效果最为显著:施氮量越高,小叶章凋落物内木质素累积越受抑制,3组的木质素残留率均在2—4月内增长最快,对照组的增长幅度(53.37%)分别是低氮组(22.99%)和高氮组(8.10%)的2.32、6.59倍。
2.2 3种类型凋落物在不同施氮水平下的生态化学计量特征
在未施氮处理时,3种凋落物初始分解及分解末期C/N的大小均表现为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物(图5)。3种凋落物初始C/P大小表现为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物;牛皮杜鹃凋落物的C/P在0—2月增长最快,涨幅约为214.52%,在后6个月持续下降,分解末期其C/P显著高于初始值;小叶章凋落物和混合凋落物的C/P在分解末期无明显差异(P>0.05),且低于初始C/P(图6)。3种凋落物初始N/P大小表现为:小叶章凋落物>混合凋落物>牛皮杜鹃凋落物,分解末期N/P均高于初始值且大小表现为:牛皮杜鹃凋落物>小叶章凋落物>混合凋落物(图7)。3种凋落物的Li/N在分解过程中均先升后降,且分解末期的Li/N显著高于初始值(P<0.05),在分解初及分解末的大小表现均为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物。其中牛皮杜鹃凋落物的Li/N在分解初始两个月增长迅速,涨幅为116.36%,在后6个月持续下降,混合凋落物的Li/N在后4个月持续下降,而小叶章凋落物的Li/N则在分解末两个月下降(图8)。
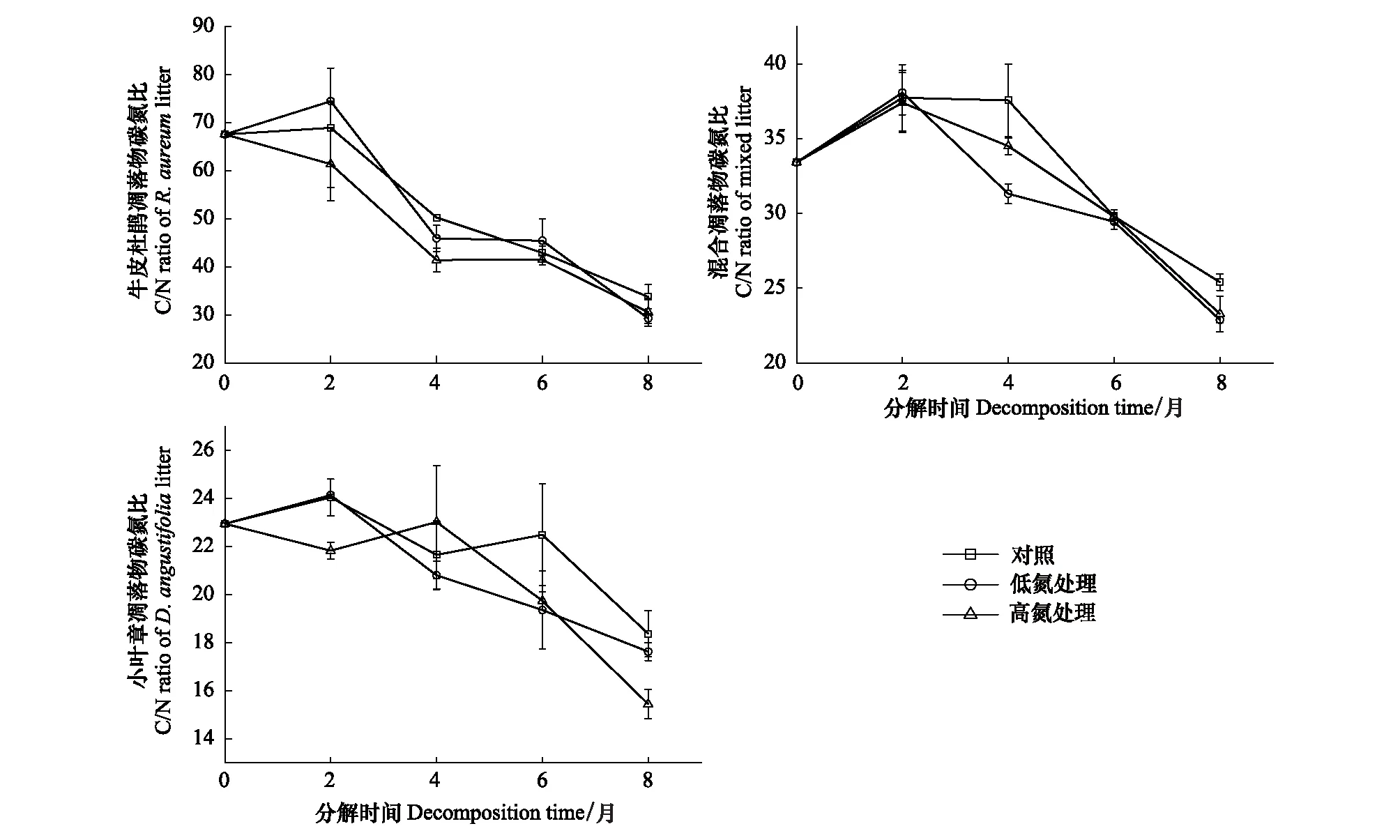
图5 不同施氮处理下3种凋落物分解过程中碳/氮的变化
分解过程中凋落物的生态化学计量特征变化受到氮沉降量及凋落物类型影响,如图5,氮添加明显促进了3种类型凋落物C/N的下降。牛皮杜鹃凋落物的C/N均在分解2—4月内下降最快,最终牛皮杜鹃凋落物C/N表现大小为:CK>HN>LN。混合凋落物的C/N在分解2—6月内各组的下降速率差异显著(P<0.05),大小表现为:LN>HN>CK,最终C/N大小为:CK>HN>LN。小叶章凋落物的C/N受施氮处理影响最大,高氮组的下降速率显著高于低氮组(P<0.05),约为低氮组的2.39倍,最终小叶章凋落物C/N大小表现为:CK>LN>HN。
如图6,在分解初两个月,氮添加显著抑制了牛皮杜鹃凋落物C/P的增长(P<0.05),增长速率表现为:CK>LN>HN;HN处理下C/P的增长速率最慢,但该组增长时间变长,最终C/P大小表现为:CK=HN>LN。施氮处理下混合凋落物的C/P在分解两个月达到峰值后持续下降,高氮组的峰值是低氮组的1.28倍,最终C/P的大小表现为:HN>LN=CK。在分解初两个月,施氮处理抑制了小叶章凋落物C/P的下降,对照组的下降速率分别是低氮组和高氮组的5.04、2.97倍;在增长阶段对照组与低氮组的变化速率显著高于高氮组(P<0.05),最终C/P大小表现为:HN>LN=CK。
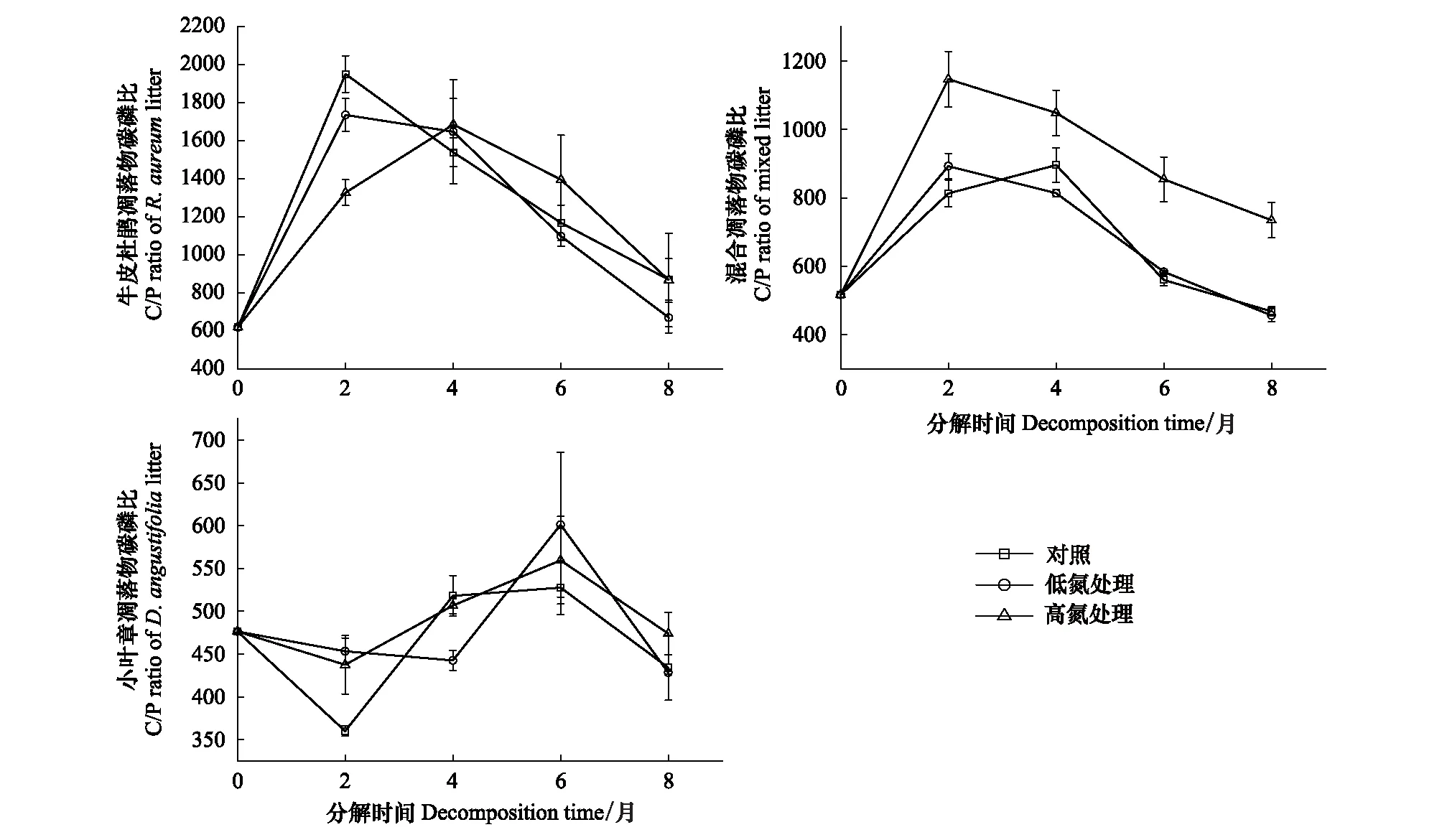
图6 不同施氮处理下3种凋落物分解过程中碳/磷的变化
如图7,牛皮杜鹃凋落物的N/P均在前4个月上升到峰值后持续下降。在分解2—4月内,低氮组和高氮组的N/P增长速率分别是对照组的5.43、8.22倍,在N/P下降阶段,施氮组的下降速率也显著高于对照组(P<0.05),最终N/P大小为:HN>CK>LN。混合凋落物的N/P明显受氮添加的促进,各分解时段的N/P大小比较均表现为HN>LN>CK,试验结束时高氮组N/P分别为对照和低氮组的1.71、1.58倍。施氮处理在分解初始2个月内抑制小叶章凋落物N/P的下降,其中HN处理抑制作用更强,最终小叶章凋落物N/P大小表现为:HN>LN>CK,高氮组的N/P约为对照组的1.30倍。
如图8,对照组牛皮杜鹃凋落物的Li/N在分解过程中呈先升后降的变化趋势,低氮和高氮组呈升-降-升-降的变化趋势,且均在0—2月增长最快,在6—8月下降最快,最终牛皮杜鹃凋落物Li/N的大小表现为:CK>LN=HN。混合凋落物的Li/N在分解过程中均先升后降,在初始两个月内低氮处理下的凋落物Li/N增长最快,在分解末两个月施氮组的下降速率高于对照组,最终Li/N的大小表现为:CK>LN=HN。小叶章凋落物的Li/N均呈先升后降的变化趋势,变化速率整体表现为:HN>CK>LN,最终Li/N大小为:CK>LN>HN。
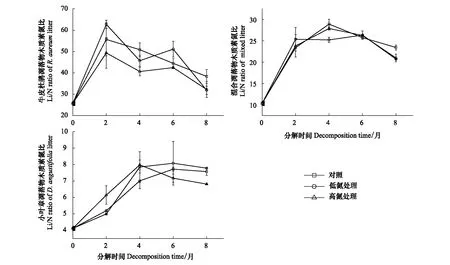
图8 不同施氮处理下3种凋落物分解过程中木质素/氮的变化
如图9,在牛皮杜鹃凋落物分解末期,不同强度的氮添加对于凋落物的C/N变化具有显著影响,牛皮杜鹃凋落物在LN处理(10 g N m-2a-1)下的C/N相较对照组明显下降,降幅约为13.30%;后随氮浓度增加到20 g N m-2a-1,C/N略有上升但仍明显低于对照组(P<0.05)。混合凋落物施氮处理后的C/N均下降,降幅分别为10.55%(LN处理)和8.23%(HN处理)。对于小叶章凋落物,LN处理较CK组无显著影响,HN处理显著降低了其凋落物的C/N(P<0.05)降幅约为15.88%。且由图9可知,同一施氮梯度下的不同类型凋落物之间的碳氮比值具有显著差异(P<0.05)。
如图9,同一施氮水平下,牛皮杜鹃凋落物与混合凋落物、小叶章凋落物之间的C/P具有显著差异(P<0.05),混合凋落物和小叶章凋落物的C/P变化规律大致相同。牛皮杜鹃凋落物在不同施氮梯度处理下的C/P比值具有显著差异(P<0.05),与对照组相比,LN处理下的C/P降幅为22.93%,HN处理未对C/P造成显著影响。LN处理下混合凋落物及小叶章凋落物的C/P与对照无明显差异(P>0.05),HN处理下C/P增幅分别为57.81%、9.01%。
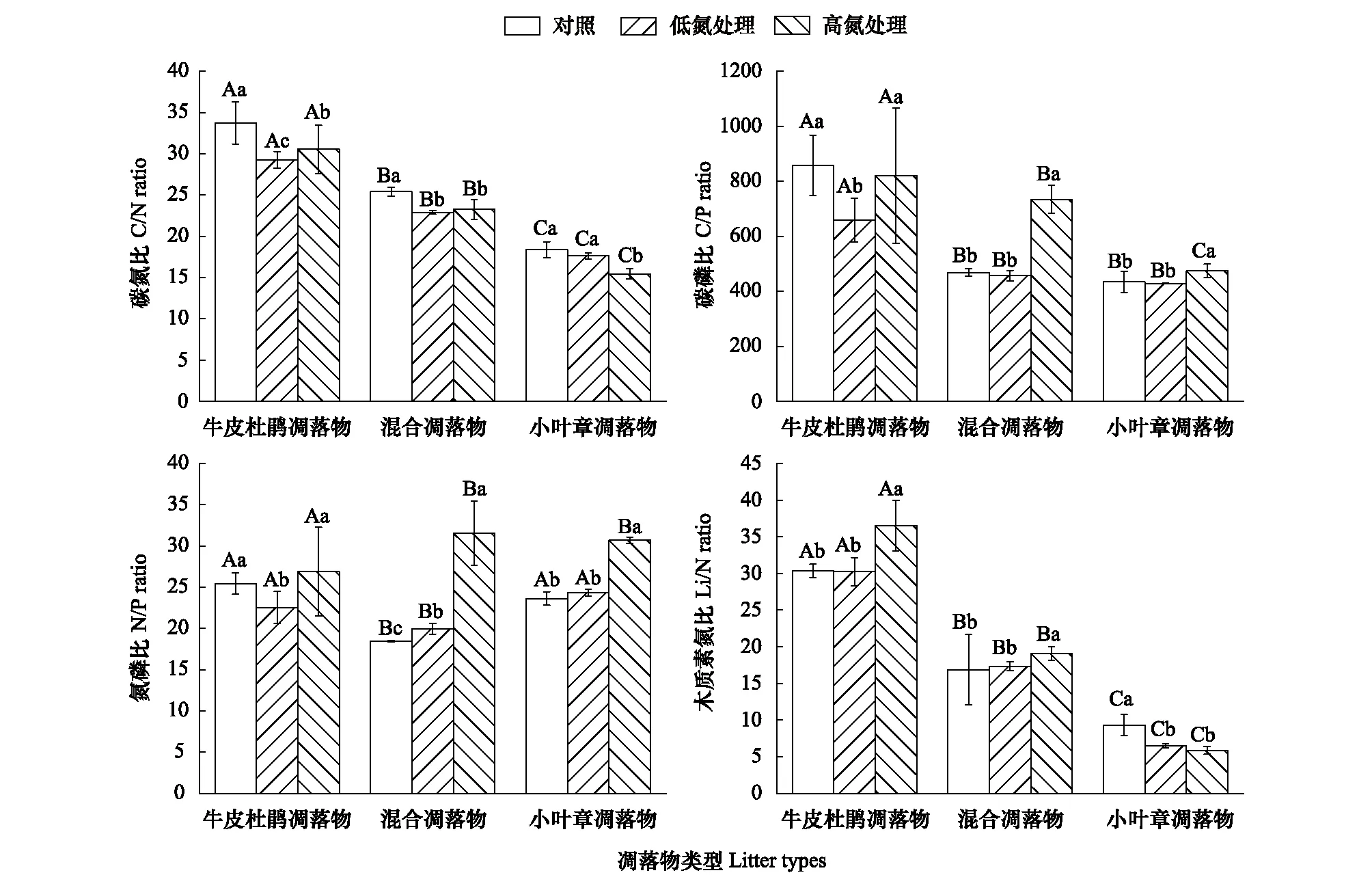
图9 氮添加对牛皮杜鹃凋落物、混合凋落物和小叶章凋落物分解末期生态化学计量特征的影响
如图9,施氮处理下3种类型凋落物的N/P变化具有显著差异。LN处理下牛皮杜鹃凋落物的N/P与对照组相比明显下降(P<0.05),降幅约为10.83%,HN处理后的N/P无明显差异(P>0.05)。混合凋落物在不同施氮梯度处理下的N/P具有明显差异(P<0.05),增长幅度分别为8.13%(LN)、73.06%(HN);LN组小叶章凋落物的N/P与对照组相比无明显差异,HN组增幅约为29.74%。
如图9,分解末期3种凋落物的Li/N大小整体表现为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物。牛皮杜鹃凋落物和混合凋落物的Li/N对于氮沉降的响应一致,LN处理对于两种凋落物的Li/N的影响不明显(P>0.05),HN处理显著促进了Li/N的增长,增长幅度分别为20.28%(牛皮杜鹃凋落物)、13.35%(混合凋落物)。施氮处理对小叶章凋落物的Li/N有显著影响(P<0.05),且随氮浓度的增加其下降幅度变大,分别为30.33%(LN)、37.08%(HN)。
在不同氮添加下,不同类型凋落物分解末期的C/N、C/P、Li/N整体大小表现均为牛皮杜鹃凋落物>混合凋落物>小叶章凋落物,N/P表现为小叶章凋落物>牛皮杜鹃凋落物>混合凋落物。3种类型凋落物的化学计量比值变化趋势上,牛皮杜鹃凋落物的变化规律与其他两种具有明显不同,混合凋落物与小叶章凋落物的变化趋势整体相似。牛皮杜鹃凋落物的化学计量比变化趋势均表现为表现为低氮降低高氮增加。随施氮量的增加混合凋落物的C/N呈现下降趋势,C/P、N/P、Li/N呈现升高趋势;小叶章凋落物的C/N、Li/N整体呈下降趋势,C/P、N/P呈升高趋势。
3 讨论
养分元素残留率指凋落物分解过程中元素在某一时刻占该元素初始质量的百分比,可以直观表现该元素在这一时刻的状况特征。元素残留率受分解环境对元素的补充和再释放的共同影响,可能出现随时间波动变化的现象,这可能与凋落物分解过程中参与分解的土壤微生物群落的活动节律有关[20]。
3.1 植物类型变化带来的苔原带凋落物化学组分和化学计量比的差异
过去的研究表明,低品质凋落物(低氮、高C/N)分解慢,在分解初期经常从环境中固定养分,含养分高的高品质凋落物(高氮、低C/N)分解快,可以在较短时间内释放养分[15],本研究结果与此一致。长白山苔原带草本入侵后凋落物类型发生变化,牛皮杜鹃凋落物质量低于小叶章和混合凋落物[16],灌草和草本凋落物比灌木凋落物分解更快;低质量的牛皮杜鹃凋落物含氮量低,在分解过程中微生物为满足自身养分需求进行了主动运输,进而可能增加了N元素在凋落物中的富集。N含量较高时,超过微生物生长所需的N就会通过分解释放到土壤中,因此小叶章凋落物和混合凋落物的N元素在分解初大量释放,前期分解远远快于牛皮杜鹃凋落物,后期凋落物分解快慢的差异在缩小。
木质素是凋落物中最难分解的复合物,结构复杂、稳定,降解最慢。周世兴等实验发现木质素降解表现为前6个月有一定的波动,随后为释放过程,分解1年后的木质素残留率达到135%[21]。本实验结果也表明3种类型凋落物的木质素残留率在分解4—6个月后开始下降,但分解结束后仍高于初始值,原因是由于本实验时间较短,木质素刚开始进行降解,未来木质素将随分解时间的延长呈稳定的降解趋势。本实验结束时牛皮杜鹃凋落物的木质素残留率最高,小叶章凋落物最低,混合凋落物次之。牛皮杜鹃凋落物中的Li累积量远高于N富集量,小叶章凋落物内Li与N的组分差异最小,混合凋落物中和了两种凋落物内化学组分差异,3种类型凋落物的C/N及Li/N在分解结束后的大小比较均为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物。木质素含量能够影响凋落物分解速率大小,是凋落物分解中极其重要的基质质量指标。C/N越低,凋落物内耐分解的化合物含量越少,凋落物分解越快。从木质素残留率及C/N的大小比较中推测出小叶章凋落物的分解最快,牛皮杜鹃凋落物最慢。
张婷等研究发现不同元素对淋溶的响应特性存在差异,磷比碳、氮更易被溶出[22]。在本试验结束时,3种类型凋落物的P元素均呈现释放状态,牛皮杜鹃凋落物的N/P及C/P均高于初始值,且增幅高于混合凋落物及小叶章凋落物。因此,长白山苔原带草本入侵后可能不利于P的释放。
长白山苔原在草本入侵后,改变了群落的组成,使凋落物质量发生变化。原来牛皮杜鹃灌木群落凋落物质量低分解慢,苔原土壤养分匮乏,当变成牛皮杜鹃-小叶章草灌混生群落和小叶章草本群落后,将加快有机物的分解和养分的释放,而且草灌和草本凋落物的化学计量比C/N、C/P和N/P无论是初始还是分解8个月都低于灌木,这将有助于草灌和草本凋落物的继续分解和养分释放。同时草本凋落物的增加更多的N释放出来,将更有利于喜氮的草本植物生长。
3.2 氮沉降增加带来的凋落物化学组分和化学计量比的差异
多数研究发现,外源 N 添加会影响土壤微生物的生长和活性,增加分解过程中的养分固持或促进了养分元素的释放[23],从而对凋落物的分解产生影响。本研究表明,N 添加对化学组分的影响明显依赖于植物类型和氮沉降水平。外源氮的输入显著促进了3种凋落物的C释放,高氮沉降比低氮沉降更利于牛皮杜鹃凋落物和小叶章凋落物的C释放。LN处理促进了牛皮杜鹃凋落物N富集,HN处理对其有显著的抑制作用,说明氮沉降量达到一定程度时,凋落物N元素的富集会受到抑制,这与樊后保等研究结果相同[24]。宗盛伟等的研究表明伴随小叶章入侵长白山苔原带的强度增加,土壤氮的含量减小[25]。在小叶章群落的养分循环中,小叶章在生长过程中从土壤中吸收了更多N元素,由于小叶章凋落物富含N元素,外源氮的添加使凋落物的N释放有显著的促进作用。高氮处理下抑制了混合凋落物N元素的释放。可能是由于牛皮杜鹃和小叶章两种不同质量的凋落物混合,发生了元素转移。3种凋落物施氮分解8个月后C/N都显著下降,原因可能是凋落物与土壤内N含量充足,外源氮输入造成N在凋落物组分内积累以及组分内的C元素释放,降低了C/N,这与Yue等的研究结论大致相同[26]。低氮处理下牛皮杜鹃凋落物和混合凋落物的C/N低于高氮组,小叶章凋落物的C/N均表现为CK>LN>HN。在外源氮添加在到达一定量(阈值)时,凋落物的C/N将到达最低值,超出阈值后,氮沉降对凋落物C/N下降的促进作用将减弱。本研究中发现氮素添加为20 g N m-2a-1时,牛皮杜鹃凋落物的C/N下降幅度减小,说明外源氮添加量已超出阈值;而小叶章凋落物C/N的下降幅度仍随氮素添加量升高而增大,可知其对应的氮添加阈值必高于牛皮杜鹃凋落物所对应的阈值,但具体的添加量还需设计更高梯度的模拟氮沉降实验来进行探究。
模拟氮沉降对混合凋落物、小叶章凋落物P元素的释放有促进作用,且氮浓度越高,促进作用越显著。低氮沉降显著抑制牛皮杜鹃凋落物的P释放,高氮沉降显著促进。这与宋学贵等对川西南常绿阔叶林凋落物的外加N试验研究表明的只要施加氮肥就会抑制凋落物中的P元素释放结果不一致[27]。其原因与参与凋落物分解的微生物群落有关,N添加影响了微生物分解者的群落结构,抑制了参与凋落物分解的微生物活性[28],从而抑制牛皮杜鹃凋落物的 N、P 释放,而促进小叶章凋落物的养分释放。但考虑到本实验中并未测定微生物酶活性和群落结构,各凋落物分解过程中的养分释放对 N 添加的不同响应是否与微生物分解者有关,仍需进一步研究。
N/P是植物生长养分限制的敏感性指数[26]。凋落物的分解往往受土壤N或P有效性的限制,因此养分限制类型也能通过凋落物的N/P来预测[29]。通常认为,N/P较低反映植物受到N限制,而较高的 N/P 则反映植物受到P限制[30]。高氮处理下牛皮杜鹃凋落物的N/P与对照组差异不大,但均高于低氮组;小叶章凋落物和混合凋落物的N/P均表现为HN>LN>CK。说明在氮素输入超出阈值后,氮浓度对牛皮杜鹃凋落物N/P下降的促进作用将减弱,而小叶章凋落物N/P随氮浓度增加而持续上升,在未来小叶章凋落物的分解很有可能由受N限制转为受P限制。
Vitousek等将凋落物分为代谢物质和结构物质,木质素、纤维素等难分解的物质是结构物质,结构物质中木质素与纤维素的含量越高,凋落物分解速率越低[31]。凋落物分解后期,木质素含量和分解过程中产生的抑制分解的化合物成为分解速率的主要限制因子。在本研究中,施氮处理显著抑制了凋落物木质素的累积,且在不同氮浓度处理下木质素残留率均呈现先升高后下降趋势,这与宋学贵等研究结果一致[27]。小叶章凋落物中的木质素在增长阶段受施氮处理的抑制最为明显,牛皮杜鹃凋落物的木质素在增长阶段虽也受氮素抑制,但被抑制程度远不如小叶章凋落物;在分解后期,凋落物内木质素的降解受到外源氮的促进,且氮素添加量越高,各群落凋落物的木质素残留率越低。3种凋落物的Li/N在施氮处理后都有显著下降,LN及HN处理对于牛皮杜鹃凋落物及混合凋落物的Li/N没有显著差异,但高浓度的氮沉降在分解后期明显促进了小叶章凋落物的木质素降解,使得Li/N在分解后期显著下降。通过实验结果有理由相信在未来高浓度的氮沉降下,长白山苔原带凋落物的分解将受到促进,其中小叶章草本群落凋落物所受影响最大。
3.3 氮沉降对土壤肥力及土壤微生物的影响
生态系统中凋落物将养分归还土壤,对维持土壤碳库和养分稳定有重要意义[32]。从凋落物8个月内的分解过程看出,施氮处理虽对化学组分的影响较为复杂,但从总体来看施氮促进了凋落物C、N、P的释放及木质素的降解,且氮素添加量越多,促进作用愈发明显。因此,可以推断施氮特别是高氮水平下有利于苔原3种凋落物C、N和P养分释放到土壤中,提升土壤肥力,有利于植物的吸收。同时模拟氮沉降显著降低了凋落物的C/N和Li/N,这两者的降低将有利于凋落物的分解,使植物可从土壤中获取更多的养分来维持体内的元素平衡,从而加快植被和土壤碳库的养分循环速率,因此氮沉降量增加将对牛皮杜鹃与小叶章的生长有促进作用。从化学组分及生态化学计量特征的变化来看,小叶章凋落物对于氮沉降的响应最为显著;从影响分解速率的基质指标来看,氮沉降下小叶章凋落物的低C/N及低Li/N都将显著促进其分解,因此小叶章凋落物的分解速率最快,牛皮杜鹃凋落物最慢,混合凋落物由于中和了两种凋落物,化学组分及化学计量比的变幅也常处于牛皮杜鹃凋落物及小叶章凋落物的中间水平,其分解速率也次于小叶章凋落物。因此推测在未来氮沉降下,小叶章草本植物、小叶章凋落物与土壤之间的养分循环加速,这一变化也将更有利于小叶章草本植物入侵长白山苔原带,并在与牛皮杜鹃灌木植物的竞争中获胜。
由于大气氮沉降的持续性和不可控性,且有研究发现凋落物的分解过程中化学组分及生态化学计量特征的变化还受微生物、土壤、水分等其他外界因素影响,因此,在全球气候变化的背景下,氮沉降持续增加对凋落物养分元素释放和生态化学计量特征的影响趋势和作用机理还应结合土壤、微生物、处理时间等进行研究。
4 结论
(1)灌草混合和草本凋落物比原生的灌木凋落物C和N元素释放快且Li累积更少;而原生的灌木凋落物的P释放略快于灌草和草本凋落物。3种植被类型凋落物的C/N、C/P、Li、Li/N大小表现均为:牛皮杜鹃凋落物>混合凋落物>小叶章凋落物,N/P表现为:小叶章凋落物>混合凋落物>牛皮杜鹃凋落物。因此,长白山苔原带草本植物入侵引起凋落物类型的变化将带来凋落物分解加快导致养分循环的变化。
(2)N 添加对化学组分及生态化学计量特征的影响明显依赖于植物类型和氮沉降水平。模拟氮沉降促进3种类型凋落物分解过程中化学组分的释放,且氮浓度越高促进作用越显著。在牛皮杜鹃凋落物分解过程中,氮素添加到达某一阈值后,其C/N、C/P、N/P、Li/N的降幅最大,后续若再增加氮素,其对化学计量比的影响均会减弱。但本实验中氮素添加量的增加促进了小叶章凋落物的C/N、Li/N下降,说明分解过程中小叶章凋落物所对应的氮素添加阈值必高于牛皮杜鹃凋落物,更高浓度的氮沉降对小叶章凋落物的影响将更为显著。
(3)长白山苔原带自然环境恶劣,影响凋落物分解的因素众多,因此,本实验将3种凋落物转移到室内进行模拟氮沉降,进而排除台风、火灾干扰、气候变化、植被演替等影响群落变化的外界因素。在单纯的外源氮添加下,小叶章凋落物的C、Li残留率及C/N、Li/N都远小于其余两种类型凋落物,说明小叶章凋落物内难分解化合物减少,其分解速率加快,土壤肥力变高,进而促进了小叶章的生长。该结果一方面佐证了近年来小叶章草本植物入侵长白山苔原带与大气氮沉降增加有紧密联系,一方面说明在未来随着氮沉降量的增加将加快凋落物与土壤、草本植物的养分循环,更有利于小叶章在与牛皮杜鹃的竞争中获胜。另外,本实验主要研究凋落物在八个月内的分解中,其化学组分及生态化学计量特征对增加的氮沉降量如何响应。而大气氮沉降对凋落物分解及生态的影响是一个长期、复杂的过程,因此,在未来仍需结合原位实验对其进行长期研究。
猜你喜欢
造纸信息(2022年8期)2022-11-10
猪业科学(2022年8期)2022-09-09
大连工业大学学报(2022年1期)2022-03-14
中国土壤与肥料(2021年5期)2021-12-02
新能源进展(2020年1期)2020-03-09
中国造纸(2019年6期)2019-09-10
山东农业科学(2017年3期)2017-03-29
读书文摘·经典(2017年3期)2017-03-21
中国汽车界(2016年1期)2016-07-18
江苏农业科学(2016年4期)2016-06-14
