
病原菌可可毛色二孢Lasiodiplodia theobromae对油梨枝条活性氧代谢的影响探究
2022-12-22 11:51刘馨怡刘远征李艳霞马蔚红王甲水李树和
中国南方果树 2022年6期
刘馨怡,徐 丹,刘远征,李艳霞,马蔚红,王甲水,李树和,张 贺
(1 天津农学院,天津,300384;2 海南大学,海口,570208;3 中国热带农业科学院海口实验站,海口,571101)
油梨PerseaamericanaMill.,又称鳄梨、牛油果等,原产于墨西哥、中美洲等地区[1]。油梨富含多种维生素以及不饱和脂肪酸,其营养价值高,深受消费者喜爱。随着我国消费水平的提高,油梨栽培面积日益扩大,当前我国油梨主要种植在海南、云南、广西、广东、福建、四川及台湾等热带亚热带地区[2-3]。近年来,油梨枝干溃疡病发生普遍,造成树势衰弱,引起早衰和枯死,严重影响油梨品质和产量。可可毛色二孢Lasiodiplodiatheobromae是油梨枝干溃疡病的主要病原菌之一[4-6],其引起的油梨枝干溃疡病主要为害枝干,枝条表皮呈黑褐色坏死,病斑位置稍凹陷,发展迅速,后期会导致整株死亡,给油梨产业造成较大的经济损失[7-8]。
目前L.theobromae引起油梨枝干溃疡病的致病机理尚不清楚。活性氧在植物和病原菌互作中扮演关键角色。病原菌和寄主植物互作过程中,寄主植物通常在侵染部位产生大量的活性氧,主要包括超氧阴离子和过氧化氢;活性氧作为毒性分子,在较高浓度下,可能引起一系列的分子伤害,如膜脂、蛋白过氧化以及DNA突变等,甚至引发植物细胞程序化死亡等[9]。鉴于当前尚缺少寄主油梨和病原真菌L.theobromae互作过程中活性氧代谢变化相关的系统研究,以及活性氧代谢变化和病情发展的密切关系研究,本研究探讨了油梨枝干溃疡病发生期间活性氧的产生、抗氧化酶以及抗氧化剂的变化特点,希望找到活性氧产生—消除的系统变化规律及其与病情发展的关系,为研究该病害发生机制以及开发高效防治技术提供参考。
1 材料与方法
1.1 材料
供试菌株DZN-29,分离自油梨枝干溃疡病典型症状枝条,经致病力测定具有强致病性,经形态鉴定及分子鉴定为可可毛色二孢Lasiodiplodiatheobromae。供试油梨品种为中国热带农业科学院海口实验站油梨种质资源圃内种植的4年生“Hass”油梨。
1.2 试验设计
试验在中国热带农业科学院海口实验站实验室进行。菌株DZN-29在PDA平板上活化培养3 d,待菌丝体长满全皿后,使用直径5 mm打孔器在菌落边缘取菌饼备用。选取健康生长、粗细一致、半木质化的当年生“Hass”油梨枝条,参考Li等[10]方法采用刺伤法接种菌饼,以接种无菌PDA作为对照。接种后0~5 d分别测量病斑大小,并取接种点周围0.5~1 cm宽皮部组织,立即放入液氮速冻,保存在-80 ℃超低温冰箱中备用,重复3次,每次每个时间点15条枝条取样。
1.3 测定方法
1.3.1 总蛋白含量测定 使用南京建成生物工程公司的总蛋白试剂盒A045-3测定。取枝条皮部样品0.1 g,加入预冷的100 mM,pH值7.4磷酸盐缓冲液1 mL研磨混匀,混合液转移至2 mL离心管中,4 °C 3 000 r/min离心10 min,吸取上清液0.1 mL稀释10倍后使用MULTISKAN GO全波长酶标仪(Thermo scientific,美国产)在562 nm处测定吸光值,按照试剂盒说明书测定,总蛋白含量单位为μg/mL。
1.3.2 超氧阴离子含量测定 样品组织中的超氧阴离子(O2-)含量采用北京索莱宝科技有限公司超氧阴离子试剂盒BC1295测定。取样品0.1 g,加入预冷的65 mM,pH值7.8磷酸钾缓冲液1 mL研磨混匀,混合液转移至2 mL离心管中,3 000 r/min离心10 min,吸取上清液按照试剂盒说明书测定,O2-含量单位为μmol/g。
1.3.3 过氧化氢含量测定 使用南京建成生物工程公司的过氧化氢试剂盒A064-1测定。取样品0.1 g,加入预冷的100 mM,pH值7.4磷酸盐缓冲液1 mL研磨混匀,混合液转移至2 mL离心管中,3 000 r/min离心10 min,吸取上清液在415 nm处测定吸光值,按照试剂盒说明书测定,过氧化氢含量单位为mmol/g。
1.3.4 丙二醛含量测定 使用南京建成生物工程公司的丙二醛试剂盒A003-1测定,方法参考试剂盒说明书,丙二醛含量单位为μmol/g。
1.3.5 抗氧化酶活性测定 枝条样品中的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)分别采用南京建成生物工程公司试剂盒SOD(A001-3),CAT(A007-1)测定,方法参考试剂盒说明书,酶活性单位为U/mg。
1.3.6 抗氧化物质含量测定 枝条样品中的抗氧化物质——抗坏血酸(AsA)、谷胱甘肽(GSH)分别采用南京建成生物工程公司试剂盒AsA(A009-1),GSH(A006-2)测定,方法参考试剂盒说明书,AsA含量单位为mmol/g,GSH含量单位为μmol/g。
1.3.7 总抗氧化能力测定 枝条样品的总抗氧化能力(T-AOC)采用南京建成生物工程公司试剂盒A015-2测定,方法参考试剂盒说明书,T-AOC单位为mmol/g。
1.4 数据分析
数据采用Excel 2010软件及SAS(version 8.1,SAS Institute,美国产)进行统计分析。t测验分析两个样品间的显著性差异。
2 结果与分析
2.1 接菌枝条发病情况
油梨当年生枝条在接种L.theobromae菌株DZN-29后出现褐色坏死性病斑,随着接种时间延长,病斑快速扩展,接种后5d,病斑长度达到60.97 mm;对照接种点没有变化(见图1)。

图1 “Hass”油梨当年生枝条接种可可毛色二孢Lasiodiplodia theobromae后发病情况
2.2 超氧阴离子(O2-)和过氧化氢(H2O2)含量变化
试验结果看出,接菌枝条皮部中O2-含量在接种后1~5 d均显著高于对照。接菌后,O2-含量在第1天至第4天持续增加,随后在第5天略有下降;而对照枝条皮部中O2-含量变化较小,在调查时间点均保持较低水平。接菌枝条皮部中H2O2含量快速增加,在第1天达到峰值,随后呈现下降趋势;接菌处理的H2O2含量在接种后0~3 d极显著高于对照,第4天和第5天极显著低于对照;对照的H2O2含量在观察时间点均保持较低水平(见图2)。

注:*和**分别表示差异显著(p<0.05)和极显著(p<0.01)。图3至图5同。
2.3 超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性变化
试验结果看出,接菌枝条皮部中SOD活性在接种后1~3 d快速上升,第3天达到峰值,随后两天活性下降;接种后1~5 d接菌处理的SOD活性均极显著高于对照,对照SOD活性在观察时间点均保持较低水平。接菌后,枝条皮部CAT活性快速升高,第1天达到峰值,第2天活性缓慢下降,第3天上升到较高水平,随后迅速下降;接种后1~3 d,接菌处理的CAT活性显著高于对照,而第4天和第5天接菌处理的CAT活性显著低于对照(见图3)。
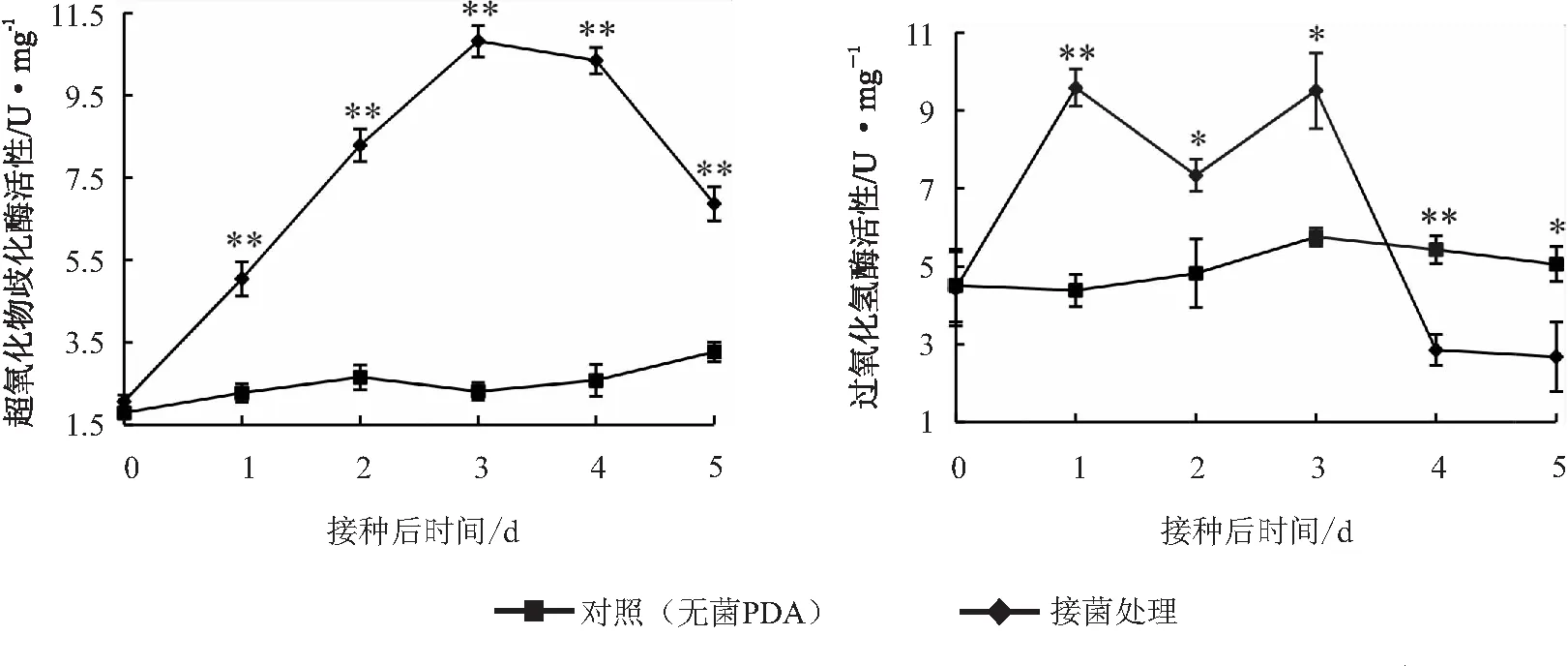
图3 “Hass”油梨当年生枝条接种可可毛色二孢Lasiodiplodia theobromae后枝条皮部超氧化物歧化酶和过氧化氢酶活性变化
2.4 抗坏血酸(AsA)和谷胱甘肽(GSH)含量变化
接菌枝条皮部AsA含量迅速上升,接种后1 d达到峰值,随后迅速下降,2~5 d一直保持较低水平;接种后1~4 d,接菌处理的AsA含量均极显著高于对照。接种后1 d,接菌处理的GSH含量快速升高至峰值,随后2~3 d呈现下降趋势,4~5 d又稳步上升;接种后1~5 d,接菌处理的GSH含量均极显著高于对照(见图4)。

图4 “Hass”油梨当年生枝条接种可可毛色二孢Lasiodiplodia theobromae后枝条皮部抗坏血酸和谷胱甘肽含量变化
2.5 丙二醛(MDA)含量和总抗氧化能力(T-AOC) 变化
接种后1~2 d,接菌处理的枝条皮部丙二醛含量快速增加,第3天稳定在较高的水平,随后4~5 d略有下降;接菌处理的丙二醛含量在测定时间均显著高于对照。接菌处理的T-AOC迅速上升,第2 天达到峰值,在剩余的测定时间呈下降趋势;接种后1~5 d,接种处理的T-AOC在测定时间均显著高于对照(见图5)。

图5 “Hass”油梨当年生枝条接种可可毛色二孢Lasiodiplodia theobromae后枝条皮部丙二醛含量和总氧化能力变化
3 结论与讨论
病原菌侵染植物过程中,寄主一般迅速地产生大量的活性氧,如O2-和H2O2,作为防御反应的信号物质调控一系列的抗性反应;但是大量活性氧的积累会形成氧化胁迫,造成植物细胞中DNA、蛋白质和脂质等大分子物质氧化损伤等,严重时导致细胞死亡,组织褐化、坏死等症状[11]。本研究发现,L.theobromae侵染油梨枝条前期,接种后1~3 d,接种点周围组织中O2-、H2O2和膜脂过氧化产物MDA的含量均大量增加,且显著高于对照。说明在油梨枝干溃疡病发病过程中,活性氧大量产生且可能引起枝条组织中细胞发生氧化胁迫。活性氧作为毒性分子,可能导致寄主植物组织细胞的过氧化,引起细胞坏死。类似的结果在其他病害发生时也观察到,如小麦壳针孢叶枯病菌Septoriatritici在侵染小麦过程中伴随着大量过氧化氢的产生和积累[12]。在L.theobromae侵染龙眼果实的过程中,O2-和MDA含量大量增加,促进了龙眼果皮褐化和细胞坏死,加速了病害的发展[13]。桃流胶病发展过程中也发现,L.theobromae侵染枝条导致寄主组织中O2-、H2O2和MDA的含量显著提高,引发寄主植物细胞膜脂过氧化[14]。由此可知,病原菌侵染过程中,寄主组织活性氧和MDA含量变化和病情发展具有重要关系。
活性氧产生和消除的动态平衡对于保持细胞稳态具有重要意义。当活性氧大量产生时,植物通过激活自身的抗氧化系统,如抗氧化酶和抗氧化剂进行活性氧的消除,从而维持动态平衡。如果抗氧化系统无法及时消除活性氧,导致活性氧在细胞中积累,细胞稳态被打破,造成伤害。本研究发现,接种L.theobromae的油梨枝条中SOD活性在接种后1~3 d快速提高,第3天达到峰值,随后SOD活性有所下降;在整个侵染期间SOD活性均极显著高于对照。而CAT活性在病原菌侵染期间存在较大程度的波动,接种后1 d快速增加达到峰值,第2 天活性缓慢下降,第3天上升到较高水平,随后迅速下降至较低水平。说明病原菌L.theobromae在侵染前期明显诱导SOD、CAT活性增强,提高了寄主植物的抗氧化防御能力,促使其消除活性氧,从而避免细胞遭遇氧化胁迫,细胞损伤和死亡。而SOD、CAT活性在接种后3~5 d逐渐下降可能与寄主细胞已遭受氧化胁迫,寄主细胞膜过氧化,细胞结构发生氧化损伤有关。有研究表明,植物抗氧化酶的活性和寄主的抗病能力密切相关。例如,灰霉菌Botrytiscinerea侵染番茄早期,寄主中的抗氧化酶如SOD、CAT活性显著提高,增强了寄主的抗氧化能力。随后伴随着病情发展,寄主SOD、CAT活性快速下降,表现出明显的相关性[15]。龙眼感染病原菌L.theobromae后,早期SOD、CAT活性迅速上升,后期有所下降,活性氧清除能力下降导致活性氧在寄主组织中大量积累,从而导致龙眼果实抗病性丧失[13]。而真菌Pyriculariaoryzae侵染小麦期间,小麦抗病品种中SOD、CAT活性快速升高,发病症状较弱[16]。甘蔗过氧化氢酶编码基因ScCAT2在烟草中超表达,显著提高了烟草对病原菌Ralstoniasolanacearum和Fusariumsolanivar.coeruleum的抗性,表明过氧化氢酶参与寄主植物的抗性反应[17]。
本试验中,抗氧化剂AsA和GSH在接种L.theobromae后快速增加,第1 天达到峰值,随后下降;而接菌后1~4 d枝条皮部的AsA和GSH含量均显著高于对照。接种L.theobromae后,油梨枝条的总抗氧化能力也快速增加,在第2 天达到峰值,随后下降。表明抗氧化剂和植物抗氧化能力在病菌侵染早期可能发挥重要作用,参与活性氧清除。同时,接菌枝条组织中O2-和MDA含量仍维持在较高的水平。可以推断病原菌L.theobromae侵染枝条导致寄主组织快速产生了大量的活性氧,激活了寄主的抗氧化系统。但是,随着侵染不断发展,病原菌可能抑制寄主细胞中抗氧化剂的合成,导致寄主细胞的抗氧化能力下降,从而累积活性氧,引发寄主细胞膜脂过氧化,促进了病害发生。这与接菌枝条病斑在侵染后期快速扩展是一致的。病原真菌樟疫霉Phytophthoracinnamomi侵染桉树的研究表明,对樟疫霉抗性较高的桉树中,AsA和GSH含量在病原菌侵染期间是不断提高的,表明寄主植物桉树对樟疫霉的抗性和AsA和GSH的含量密切相关[18]。此外,病原菌L.theobromae侵染龙眼的研究表明,寄主组织中AsA和GSH的含量的下降导致了龙眼果实表皮褐化以及果肉腐烂[13]。综上所述,推测油梨对枝干溃疡病的抗病能力与寄主的抗氧化能力密切相关,寄主抗氧化能力变化影响油梨枝干溃疡病病情发展。这一推论仍需要进一步验证。
本研究以“Hass”油梨当年生枝条为材料,研究了接种病原菌L.theobromae后枝条组织中活性氧产生—消除系统相关物质及酶活性的变化趋势。结果表明,接种病原菌早期,油梨枝条的O2-、H2O2和MDA含量快速增加,显著高于对照;而抗氧化酶SOD和CAT活性和抗氧化剂AsA和GSH在侵染早期含量较高,后期呈下降趋势。本研究表明,病原菌L.theobromae侵染引起了寄主活性氧的暴发并激活植物抗氧化系统,随着病情的发展,寄主植物的抗氧化能力不断削弱,导致植物细胞中活性氧不断积累以及产生MDA,加速了病害发展。病原菌L.theobromae侵染油梨枝条过程中,寄主植物活性氧代谢变化显著,可能和病害发生密切相关,其具体机制有待进一步研究。
猜你喜欢
现代农业科技(2022年5期)2022-12-14
当代水产(2022年1期)2022-04-26
科学导报(2022年11期)2022-03-03
中国农业科学(2021年23期)2022-01-14
昆虫学报(2021年3期)2021-05-11
智慧健康(2020年9期)2020-12-03
中国瓜菜(2020年8期)2020-09-26
世界热带农业信息(2016年6期)2016-07-02
世界热带农业信息(2016年3期)2016-04-05
世界热带农业信息(2016年3期)2016-04-05
