
典型喀斯特区不同植被恢复方式苔藓功能性状及其对土壤因子的响应
2022-12-26 13:44肖露梅王彩艳胡培雷陈元凯王克林
生态学报 2022年23期
肖露梅,张 伟,王彩艳,胡培雷,陈元凯,王克林,*
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 中国科学院大学, 北京 100049 4 湖南农业大学, 长沙 410128 5 四川省仁寿县农业农村局, 眉山 620500
植物功能性状是与植物生长、繁殖和生存密切相关的一系列属性,其不仅能响应生存环境的变化,还会影响生态系统功能[1]。因此,研究植物功能性状有利于制定更好的生态系统恢复和管理策略。植物功能性状与环境因子的关系是研究的热点问题,但目前的研究主要针对维管植物[2—3],而缺乏对非维管植物(如苔藓)功能性状与环境因子关系的研究。苔藓作为先锋植物在生态系统中广泛分布,并在脆弱或退化生态系统中占据了重要的生态位[4]。苔藓在水土保持,改善土壤肥力、增加生物多样性和促进生态系统演替等方面具有重要生态作用[5—10]。因此,研究苔藓功能性状及其与环境因子的关系对苔藓生态功能认识和脆弱或退化生态系统恢复具有特殊意义。
研究表明苔藓的分布和功能性状受到气候(如光照、温度和湿度等),地理环境(如海拔、坡度和坡向等),生长基质(如土壤、岩石和树木等)及干扰(如土地利用类型)等的多种环境因子共同影响[11—17]。适宜的气候条件有利于苔藓定殖[11],地理环境和植被类型的差异会引起的生境小气候的差异进而影响苔藓分布及其性状表达[14,16]。土壤是苔藓生长基质之一,其养分、水分和pH也会影响苔藓分布[14,18—20],反过来苔藓又会作用于土壤因子。一方面,苔藓通过光合固碳、生物固氮和截留大气沉降中的养分增加土壤养分的输入[15,21];另一方面,苔藓大量的阳离子交换点和不发达的角质层使其能吸收溶解在土壤水中的矿质元素(如N、P、K等)而使得土壤养分减少[22—23],因此苔藓营养性状与土壤因子的关系存在争议。苔藓冠层性状(如单位面积冠层质量(Canopy mass per area, CMA)、冠层高度(Canopy height, CH)、冠层密度(Canopy density,CD)和苔藓光合作用、养分和水分循环等密切相关[24—28],与苔藓营养性状相互权衡以适应生境[24,29],因此苔藓冠层性状与土壤因子的关系也具有不确定性。苔藓生物量(Bryophyte biomass,BB),苔藓饱和吸水量(Saturated water adsorption content, SWAC)和营养元素含量是苔藓参与土壤养分和水分循环的重要功能性状[10,16,30]。因此,研究苔藓功能性状与土壤因子的关系可以揭示苔藓对土壤的适应策略,从而为表层土壤生态恢复提供依据。
西南喀斯特地区是我国主要生态脆弱区之一,人地矛盾尖锐,部分以石漠化为特征的土地退化严重,生态环境亟需改善[31]。退耕还林还草工程实施后,维管植物迅速恢复[32],苔藓拓殖,形成了维管植物与苔藓共存的生态系统[33]。喀斯特地区部分研究就苔藓生物量、持水性能和营养元素探讨了苔藓对石漠化土地生态恢复的促进作用[16,30,34—35],苔藓在一些不适宜维管植物生长的石漠化地区的土壤和水文过程具有较大的应用潜力[36]。但在与维管植物共存的生态系统中,苔藓在生境的适应能力将会影响其生态功能的发挥。维管植物是喀斯特退化生态系统的恢复主体,可以在数年或十余年内显著改善土壤养分和提升水源涵养,因而不同植被恢复方式间土壤因子存在差异[37—40]。研究不同植被类型下苔藓功能性状及其对土壤因子的适应,有利于探索更优的喀斯特退化生态系统修复方式(如将苔藓和维管植物结合)。因此,本文以喀斯特峰丛洼地退耕后人工林,牧草地和人工林+牧草下苔藓为研究对象,分析了苔藓冠层性状、营养性状(营养元素含量及其化学计量比)、生物量和持水性能及其对土壤因子的响应,以期为苔藓应用于喀斯特退化生态系统恢复提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于广西壮族自治区环江毛南族自治县下南乡古周村典型峰丛洼地小流域(24°54′—24°55′N,107°56′—107°57′E)。该研究区海拔为376—826 m,属亚热带季风气候区,年均气温16.9℃,多年均降雨量1675 mm,雨季集中在4—9月。土壤为碳酸盐岩风化形成的钙质石灰土,洼地平均土壤深度为50—80 cm,坡地平均土壤深度为10—30 cm,基岩广泛裸露。该区域从2002年开始实行“退耕还林还草”生态工程,位于洼地的耕地退耕后采用3种人工恢复方式,分别为单一物种人工林恢复,恢复树种为任豆(Zeniainsignis);人工牧草地恢复,种植桂牧一号牧草 (Pennisetumpurpureumcv,Guimu- 1);以及任豆人工林间作牧草(mix-plantation ofZeniainsignisandPennisetumpurpureumcv,Guimu- 1)。
1.2 样品采集
2020年5月选取该研究区人工林、牧草地和人工林+牧草这三种植被类型,每种植被类型随机设置4个样方,共计12个样方,样方大小均为20 m×20 m。每个样方苔藓盖度(%)用4个1 m×1 m(100个10 cm×10 cm网格)的样方框估算。每个样方随机设置4个1 m×1 m的样方,每个样方再随机设计4个小样方(10 cm×10 cm),即每个样方采集16个小样方用作苔藓物种鉴定。另外,每个样方再随机采集8个10 cm×10 cm的苔藓样品用于功能性状的测定,同时采集苔藓下覆0—2 cm土壤,将土壤混合均匀代表该样地的土壤样品。记录样地裸岩率、植被盖度和苔藓盖度信息,如表1所示。
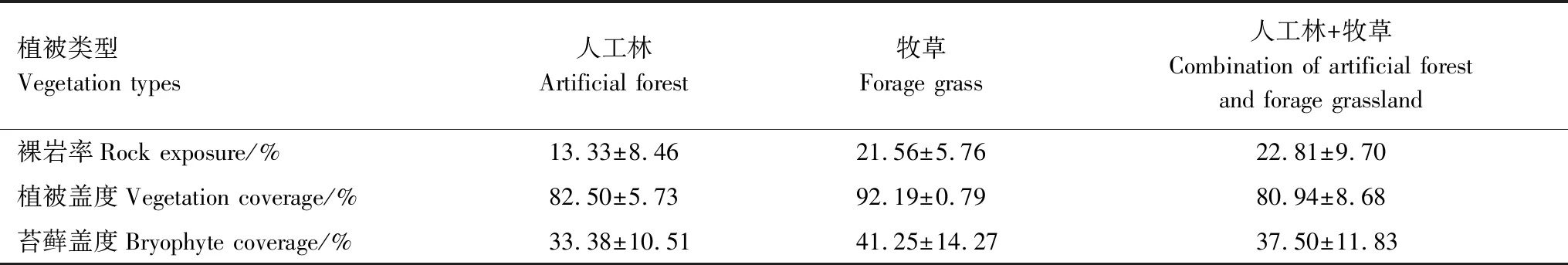
表1 各植被恢复方式下植被和基岩出露特征
1.3 苔藓功能性状和土壤因子测定
采集苔藓样品时测定苔藓CH,每个样方测定8个重复。用于测定功能性状的苔藓样品先用纯水清洗干净,一部分样品在105℃下杀青30 min,再在65℃烘干至恒重,称重并记录冠层重量,然后粉碎过筛;一部分用于SWAC的测定。根据Waite和Sack的计算方法[24],CMA=冠层重量/冠层面积,CD=CMA/CH。BB=CMA×苔藓盖度。苔藓先在65℃烘干至恒重测量干重,饱和吸水后置于过滤网中,待不滴水后测定其饱和吸水重量,苔藓饱和吸水率=(苔藓饱和吸水重量-苔藓烘干重)/苔藓烘干重,SWAC=苔藓饱和吸水率×BB[41]。苔藓C、N采用碳氮元素分析仪(Vario-MAX,德国)测定。苔藓P采用混合催化剂(K2SO4∶CuSO4=10∶1)催化、H2SO4消化后,用流动注射仪测定。苔藓K采用 H2SO4-H2O2消煮、原子吸收法测定。单位面积苔藓C、N、P和K含量(Carea、Narea、Parea和Karea)等于C、N、P和K含量乘以苔藓生物量。
1.4 苔藓鉴定与多样性指数计算
利用Nikon E100型光学显微镜对苔藓样本进行观察,参考《中国苔藓志》第二[42]、四[43]、五[44]、六[45]、七[46]、八卷[47]将苔藓鉴定到种。根据韩淑婷等[17]和申家琛等[48]的计算方法,计算苔藓重要值、物种丰富度、Shannon-Wiener指数、Simpson优势度指数和Pielou均匀度。
1.5 数据处理与分析
采用Excel 2019进行数据整理,使用SPSS 25.0对数据进行正态检验、方差齐性检验和主成分分析。对不同恢复植被方式下的各项指标采用单因素方差分析(One-way ANOVA)和最小显著性差异法(Least significant difference, LSD)进行比较。采用Canoco 5.0对苔藓功能性状和土壤因子进行冗余分析(Redundancy analysis, RDA)。表中数据为平均值±标准误差。
2 结果与分析
2.1 不同植被恢复方式苔藓功能性状
本研究共调查发现苔藓植物9科11属11种,均为藓类(表2)。三种植被恢复方式下苔藓优势种均为匐灯藓(重要值>0.3),此外人工林下苔藓优势种还有薄壁卷柏藓(表2)。苔藓多样性指数在各植被类型间均无显著差异(表3)。
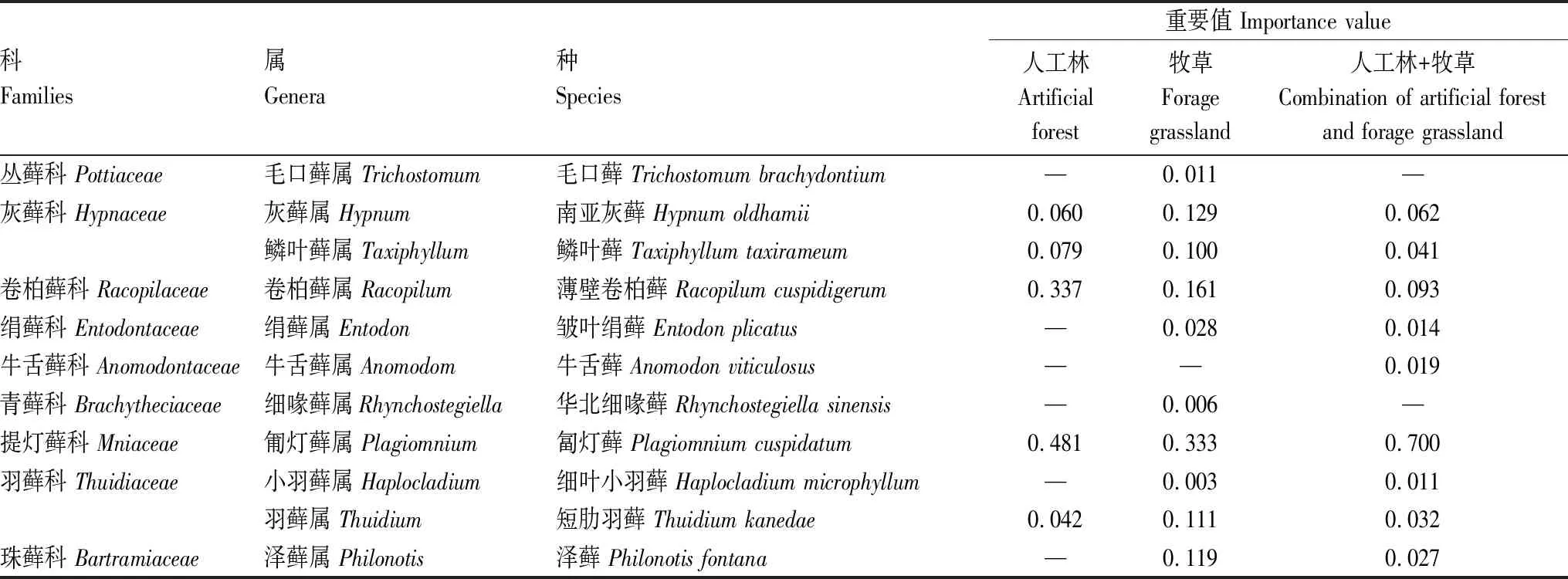
表2 各植被恢复方式下苔藓物种组成及重要值
对苔藓功能性状指标的主成分分析表明,特征根>1的前4个主成分的累积贡献率达90%,能全面反映所有信息(表4)。对苔藓功能性状指标的主成分分析表明,特征根>1的前4个主成分的累积贡献率达90%,能全面反映所有信息(表4)。第一主成分主要反映苔藓营养性状(营养元素含量及其化学计量比),人工林+牧草在第一轴显著区别于人工林(图1),其中苔藓K在人工林+牧草显著低于人工林,苔藓C:K、N:P和N:K在人工林+牧草显著大于人工林(表5)。第二主成分主要反映苔藓冠层性状及生物量特征,牧草地在第二轴显著区别于人工林和人工林+牧草(图1),牧草地苔藓CH、N显著低于人工林和人工林+牧草,但苔藓CMA、CD和BB在牧草地有最大值(表5),第三和第四主成分在各植被类型下无显著差异。
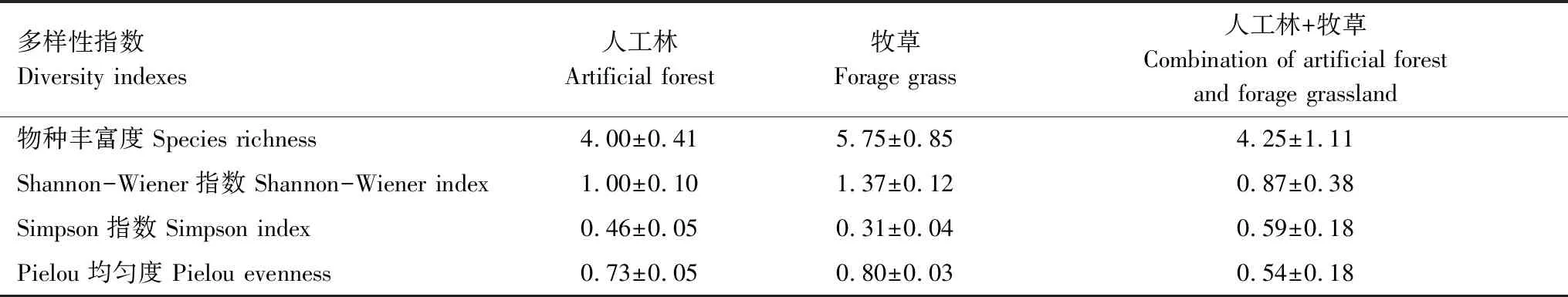
表3 各植被恢复方式苔藓多样性指数
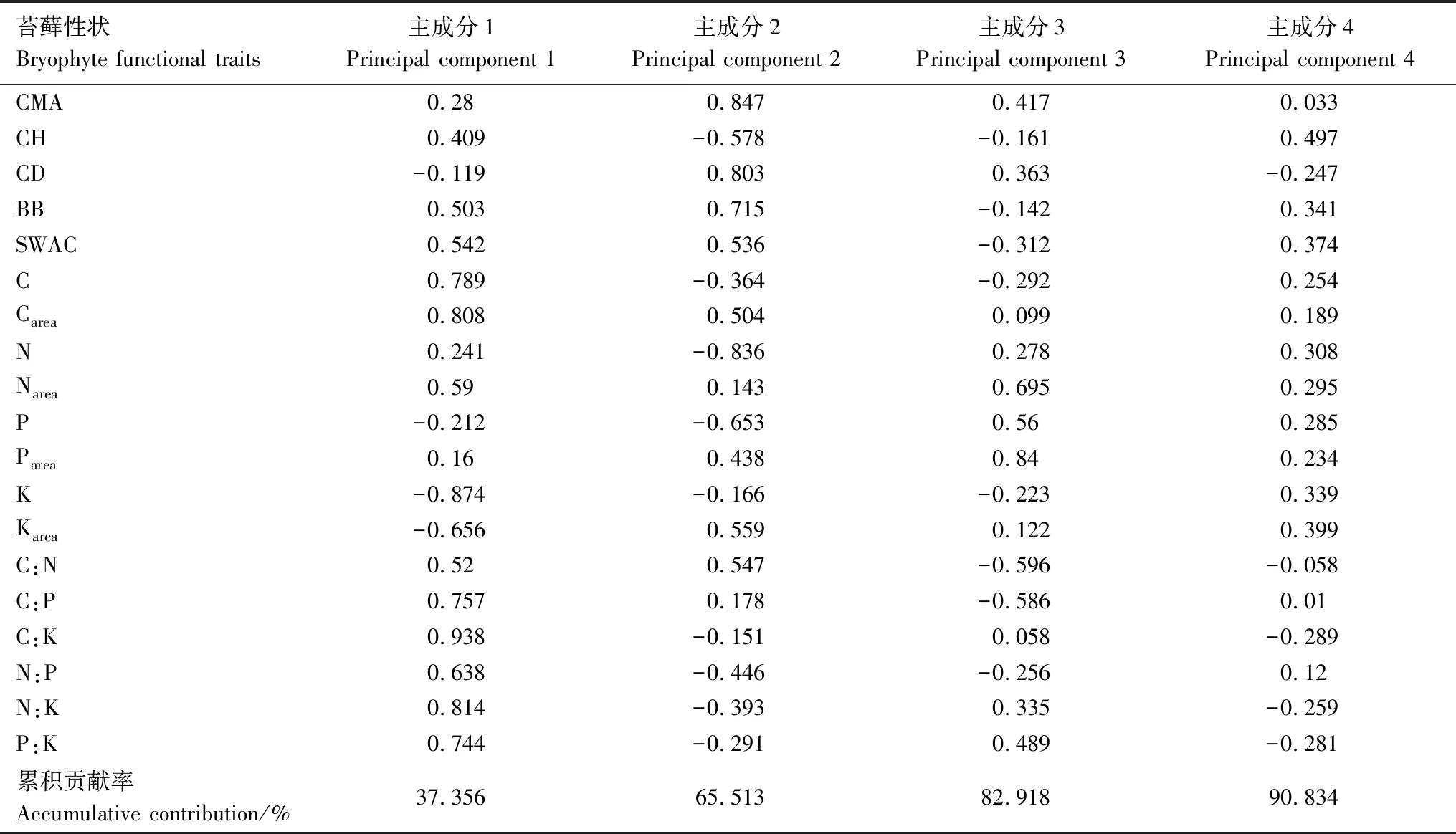
表4 苔藓功能性状主成分分析
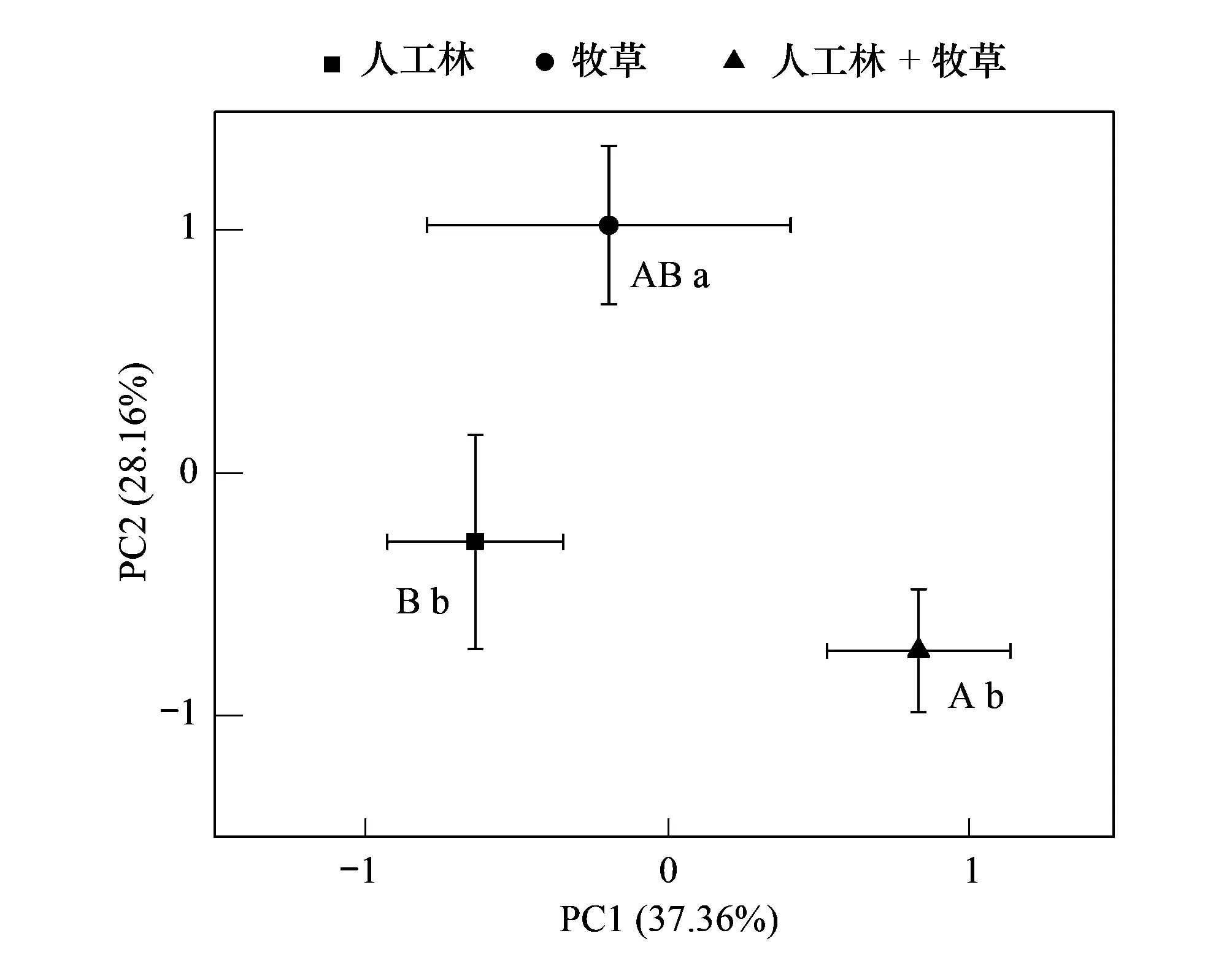
图1 三种植被恢复方式下苔藓功能性状主成分分析PCA Fig.1 Principal component analysis of bryophyte functional traits in three vegetation restoration methods不同大写字母表示三种植被恢复方式在第一主成分的差异显著(P<0.05),不同小写字母表示三种植被恢复方式在第二主成分的差异显著(P<0.05)
2.2 不同植被恢复方式土壤因子
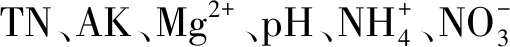
2.3 苔藓功能性状与土壤因子的关系
RDA分析表明,Mg2+和SWC是影响苔藓功能性状的主要土壤因子(图2)。苔藓营养性状(营养元素含量及化学计量比)与土壤因子主要呈正相关关系,苔藓冠层性状(如CMA和CD),生物量及关联指标(BB,SWAC和单位面积营养元素含量)与土壤因子主要呈负相关关系(图2)。
3 讨论
3.1 不同植被恢复方式苔藓功能性状
三种植被类型下苔藓多样性指数无显著差异,但不同植被类型下苔藓功能性状存在差异。苔藓CMA与维管植物单位面积叶片质量具有相似涵义,与苔藓养分获取和水循环有关,CMA越大有利于储存养分和保存水分,反之,有利于增强植物获取资源能力和水分蒸发[24]。苔藓CD与光合作用有关[24],苔藓CH会影响苔藓冠层内部光、温、水的分布,从而影响土壤-大气界面水热交换[25—26]。苔藓CH高、CD小有利于苔藓植株间水汽和热量的对流,蒸发作用强,反之,蒸发作用弱[26]。同时,苔藓CH影响光照的垂直分布,苔藓可利用光随苔藓冠层高度加深而变少[25]。牧草地植被覆盖度高,牧草为苔藓提供遮阴的同时,也会影响苔藓可利用光和水分的有效获取。在阴暗生境,苔藓植株往往高度聚集以获取更多可利用光[49],所以牧草地苔藓CH最低但CD最大,有利于减少水分蒸发和增强光合作用。苔藓SWAC与苔藓蓄水能力有关[30],牧草地苔藓CMA和SWAC较大,说明牧草地苔藓养分储存和蓄水能力较强。同理,人工林和人工林+牧草的植被覆盖度较牧草地低,苔藓能获取的水分和光照更多,因此苔藓CH较高,但CD、CMA和SWAC均较小,苔藓获取资源能力强但蓄水能力较弱。因此,苔藓在牧草地的适应和生存能力更强,且苔藓持水能力也在牧草地更强。
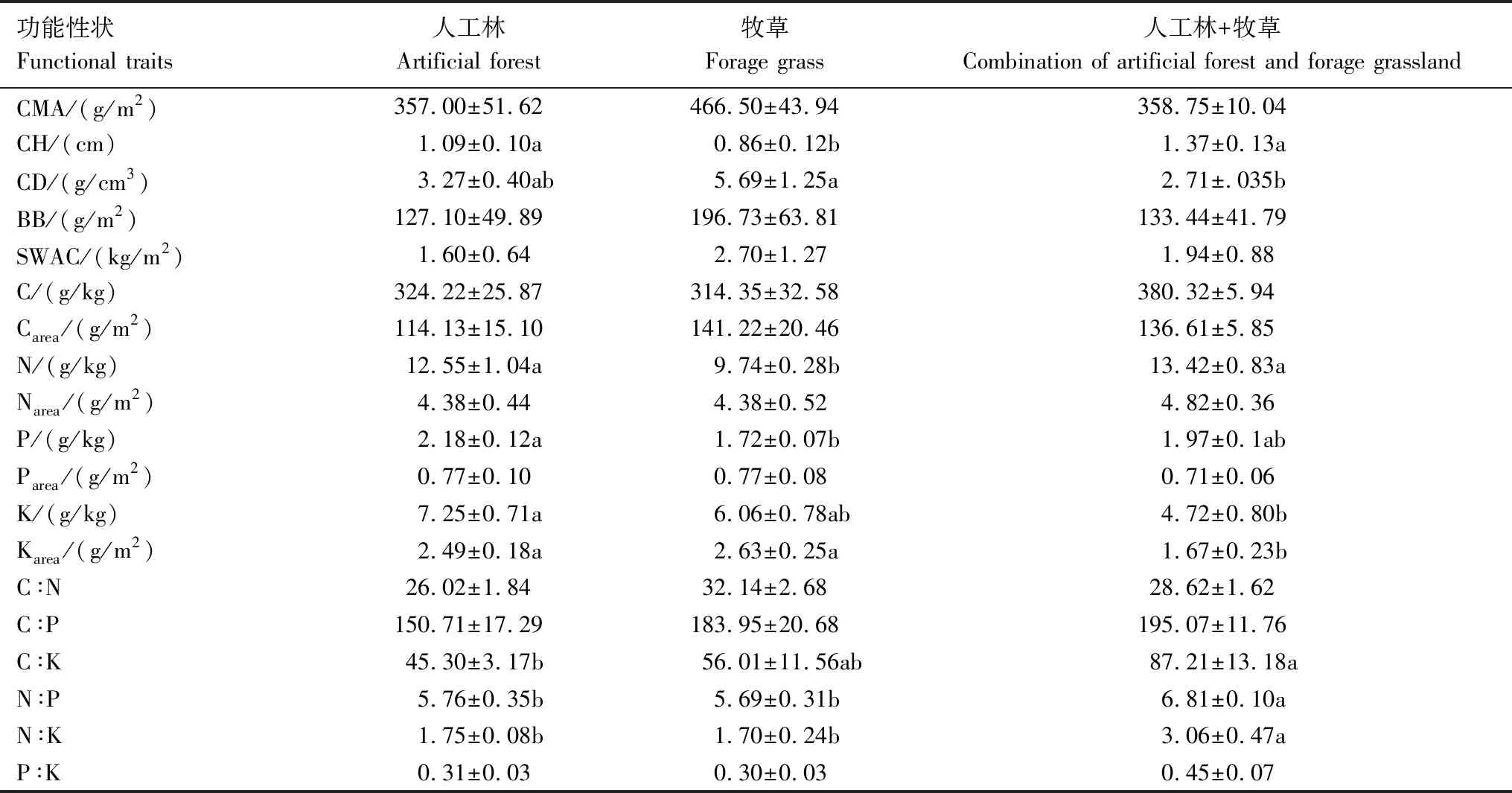
表5 各植被恢复方式下苔藓功能性状
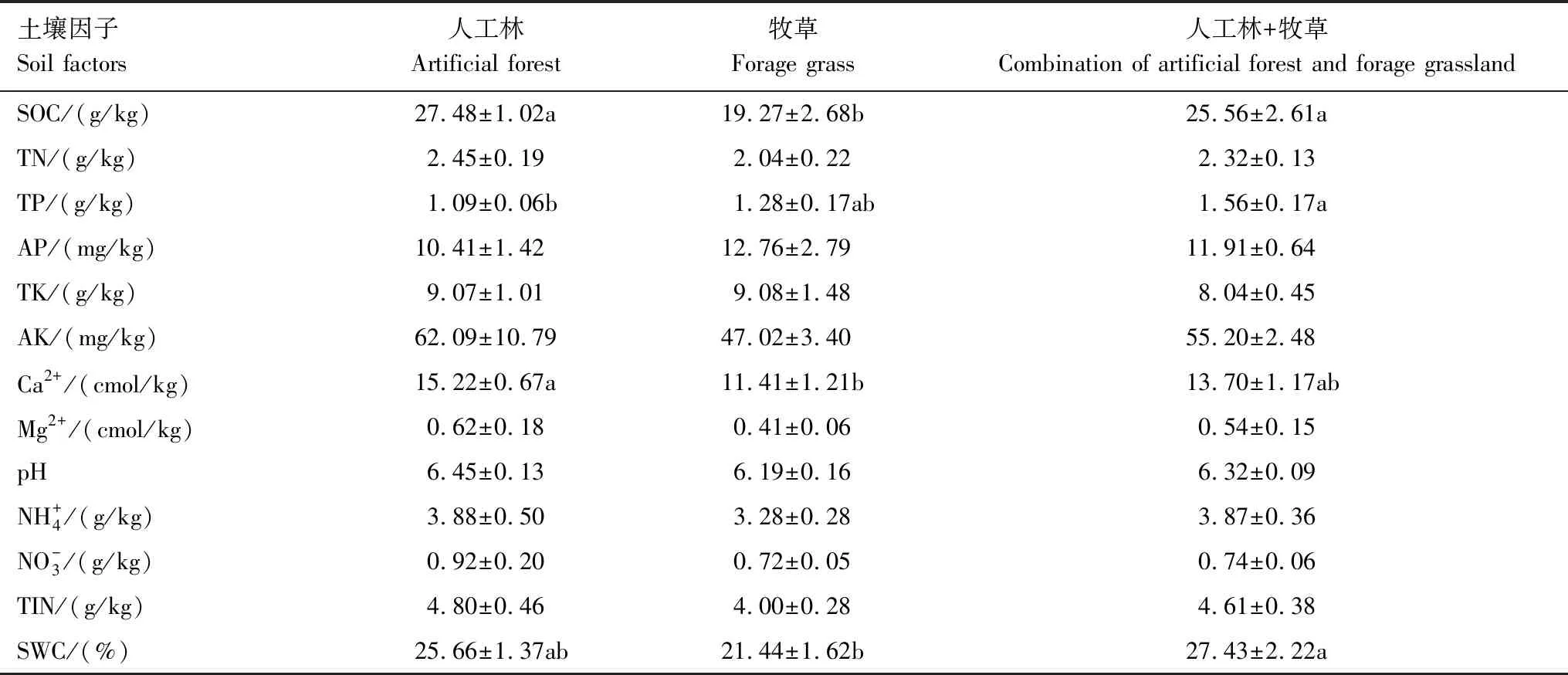
表6 各植被恢复方式下土壤因子
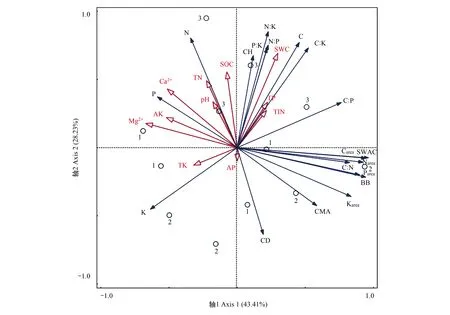
图2 苔藓功能性状与土壤因子的冗余分析Fig.2 Redundancy analysis of bryophyte functional traits and soil factors红色箭头表示土壤因子,蓝色箭头表示苔藓功能性状指标.数字1- 3分别代表不同植被恢复类型,1:人工林,artificial forest;2:牧草,forage grassland;3:人工林+牧草,combination of artificial forest and forage grassland; CMA:单位面积冠层面积 canopy mass per area; CH:冠层高度 canopy height; CD:冠层密度 canopy density; BB:苔藓生物量 bryophyte biomass; SWAC:饱和吸水量 saturated water adsorption content; C:碳含量 carbon concentration; Carea:单位面积C含量 carbon per area; N:氮含量 nitrogen concentration; Narea:单位面积N含量 nitrogen per area; P:磷含量 phosphorus concentration; Parea:单位面积P含量 phosphorus per area; K:钾含量 potassium concentration; Karea:单位面积K含量 potassium per area; SOC: 土壤有机碳 soil organic carbon; TN: 全氮 total nitrogen; TP: 全磷 total phosphorus; AP: 速效磷 available phosphorus; TK:全钾 total potassium; AK: 速效钾 available potassium; Ca2+: 交换性钙 exchangeable calcium; Mg2+: 交换性镁 exchangeable magnesium; 铵态氮 ammonium; 铵态氮 nitrate; TIN:总无机氮 total inorganic nitrogen; SWC: 土壤含水量 soil water content
苔藓营养性状(营养元素含量及化学计量比)能反映苔藓养分利用效率和生存策略[50—51]。N和P主要用于蛋白质和核酸的形成从而影响植物生长发育。牧草地苔藓CMA最大,可能会造成苔藓生长过程中营养元素稀释或牧草地苔藓获取的N和P较少,所以牧草地苔藓N和P含量最低。Waite和Sack的研究结果也表明苔藓CMA越大,N和P越低[24],但也有研究表明随CMA增加,苔藓N和P含量并不减小[27],或CMA和N含量相互独立[52],这种不一致性可能是苔藓生境差异导致的。牧草地苔藓N、P含量显著低于人工林和人工林+牧草,但C:N和C:P较高,表明牧草地苔藓养分利用效率较高。在陆生植物中,常用N:P阈值14—16[53]或10—20[54]表征植物N、P限制。本研究中苔藓N:P的范围为5.69—6.81,远低于阈值14或10,说明该研究区苔藓生长可能受N限制。但苔藓N:P因生态系统不同而存在差异[45—46,55—56],所以不同地区苔藓N、P养分限制可能存在特定的N:P比阈值。因此,喀斯特地区苔藓是否受N限制还需要进一步研究确定。K可以减轻干旱对植物生长的胁迫[57],干旱促使苔藓对K的积累[58]。人工林下苔藓CH高,水分蒸发快,牧草地下CH低,但获取水分少,因此人工林和牧草地苔藓会积累更多K以应对干旱胁迫。所以,苔藓K含量在人工林和牧草地显著高于人工林+牧草,而C:K和N:K在人工林和牧草地下显著低于人工林+牧草。因此,不同植被类型下苔藓的养分利用策略不仅反映了不同植被类型下养分供应情况,也会影响生态系统养分循环。
3.2 苔藓功能性状对土壤因子的响应与适应
苔藓营养元素来源于降水、降尘、林冠淋溶和生长基质等[22]。RDA分析表明,土壤SOC与苔藓C含量正相关,土壤TN和TIN与苔藓N含量正相关,土壤TK和AK与苔藓K含量正相关,表明土壤与苔藓C、N和K的循环和累积紧密联系,二者相互促进。而土壤TP、AP与苔藓P含量相关性较弱,可能是因为苔藓P的积累还来源于降水、降尘或林冠淋溶等[22],土壤P对苔藓P积累影响较小。但四川高山森林的研究表明,土壤P与苔藓P含量显著负相关,因为苔藓P的积累主要来源于雨水[15]。长白山针叶林的研究表明,苔藓对P的吸收造成了土壤P减少,土壤为苔藓提供了主要的P源[23],因此苔藓与土壤P相关性因生态系统的差异而有所不同。苔藓冠层性状及生物量综合反映苔藓对营养元素的吸收和利用情况。苔藓CMA、CD、BB及单位面积营养元素含量与土壤养分主要呈负相关关系,可能有两点原因:一是在维管植物占主导地位的生态系统,表层土壤养分的积累主要来源于因维管植物恢复而增加的凋落物输入,但凋落物覆盖作用会抑制苔藓生长[59];二是苔藓可以有效固持大气氮沉降中的N从而减少土壤N的输入[60],并且苔藓可以吸收溶解在土壤水中的养分用于生物量累积,从而在一定程度上造成苔藓附近土壤养分的减少,因而苔藓生物量及关联指标(BB,SWAC和单位面积营养元素含量)与土壤养分指标负相关。因此,土壤养分会影响苔藓养分获取,冠层性状及生物量的积累,进而影响苔藓生态功能。
此外,喀斯特地区土壤具有高钙镁离子和高pH值特性。研究表明,适宜的Ca2+浓度对孢子萌发和原丝体的发育有积极作用,但Ca2+浓度过高会抑制孢子萌发和原丝体发育[61]。Mg是合成与光合作用相关酶和叶绿素的重要物质,但Mg过量会对植物产生毒害作用[62],而且苔藓更适合在微酸性环境中生长[63—64]。苔藓冠层性状(CMA和CD)、生物量及关联指标(BB、SWAC和单位面积营养元素含量)和养分利用率指标(如C∶N和C∶P)与土壤Ca2+、Mg2+、pH皆呈负相关关系,说明土壤高Ca2+、Mg2+、pH特性在一定程度上会抑制苔藓生长。苔藓是变水植物,只有在湿润的环境中才有生理活性,因此环境水分条件对苔藓光合代谢具有重要作用[65]。苔藓可从雨水、大气及生长基质等获取水源,土壤水分的有效补给有利于苔藓正常光合固碳和生长。土壤SWC越高,苔藓可利用水源多,苔藓既不会因为临时的干旱影响光合作用,也不需要积累大量K应对干旱胁迫,所以苔藓CH、C、C∶K、N∶K、P∶K与土壤SWC呈正相关关系。RDA前两轴的解释率为71.64%,说明土壤因子能很好解释喀斯特峰丛洼地维管植被恢复过程中苔藓功能性状差异,其中Mg2+和SWC是影响苔藓功能性状的主要土壤因子。因为土壤Mg2+和SWC会影响苔藓光合作用受胁迫情况,进而影响苔藓其他生理活动的进行及其功能性状的表达。
人工林和人工林+牧草土壤养分含量和SWC都较高,苔藓具有高CH和低CD、CMA的冠层结构,苔藓BB和SWAC都较低,苔藓营养元素含量较高但养分利用效率较低。牧草地土壤养分含量和SWC较低,苔藓有高CD、CMA和低CH的冠层结构,苔藓BB和SWAC都较高,苔藓营养元素含量较低但养分利用效率较高。而牧草地苔藓较高的生物量和持水能力有利于改善牧草地土壤养分和水分条件。研究表明,苔藓抗逆性强,能通过协调其冠层性状、营养性状、生物量和持水性能适应不同维管植被类型生境,进而参与土壤养分和水分循环。其中,牧草地最适宜苔藓拓殖和生长,其次是人工林+牧草地和人工林。因此,可考虑将苔藓与牧草相结合应用于喀斯特退化生态系统的早期修复中。
4 结论
喀斯特峰丛洼地不同植被恢复方式下维管植物的差异造成了土壤因子的差异,苔藓表征出不同的冠层性状、营养性状、生物量和持水性能组合以适应不同植被类型生境,个别功能性状能在苔藓生理活动受胁迫时发挥其功能。土壤因子与苔藓营养性状主要呈正相关关系,与苔藓冠层性状、生物量及关联性状主要呈负相关关系,交换性镁和含水量是植被恢复过程中影响苔藓功能性状的主要土壤因子。因此在将苔藓运用于喀斯特退化生态系统的修复中时,应考虑土壤因子对苔藓分布和功能性状的影响。牧草地土壤养分和水分含量较低,但苔藓养分利用率高,生物量大,持水能力强,有利于改善牧草地养分和水分条件,因此牧草-苔藓组合可作为喀斯特退化生态系统初期恢复的较好选择。
猜你喜欢
中国农业信息(2022年1期)2022-05-25
农业机械学报(2021年11期)2021-12-07
军事文摘(2021年18期)2021-12-02
润·文摘(2021年11期)2021-11-20
中国水土保持科学(2019年6期)2019-04-26
儿童故事画报·自然探秘(2017年7期)2018-03-14
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
——一种分层模拟的方法
自然资源遥感(2017年2期)2017-04-27
农业环境科学学报(2017年2期)2017-03-20
