
内蒙古大兴安岭南段山杨径向变化特征1)
2022-12-28 05:37陈佳佳周梅赵鹏武
东北林业大学学报 2022年12期
陈佳佳 周梅 赵鹏武
(内蒙古赛罕乌拉森林生态系统国家定位观测研究站(内蒙古农业大学),呼和浩特,010019)
石亮 许重阳
( 地表过程分析与模拟教育部重点实验室( 北京大学城市与环境学院) )
王满才
( 巴林右旗森林草原保护发展中心)
中国北方半干旱区森林生长受气候干旱化制约明显,并且这种趋势还在增加[1],天然林内已经出现了不同程度的生长下降现象[2-3]。气候变化对半干旱区树木径向生长以及生态系统结构、功能的影响巨大[4],因此,掌握树木生长对气候的响应规律和应对干旱胁迫的调节机制,探讨气候变化对森林生态系统的影响,对半干旱区森林的管理及可持续发展具有重要意义。
大兴安岭南段是我国北方半干旱区的典型区域,以往对这一区域内山杨径向生长响应气候变化的研究是基于年际尺度上进行的[5-6],对于天或季节尺度上的径向生长动态及其与气候变化关系的研究相对较少。以往年际尺度上的树木径向生长研究主要通过相关分析获得径向生长与环境因子之间的关系,缺乏明确的生理基础[7-8]。而运用树木径向生长测量仪则可以开展高分辨率的连续监测,明确树木的年内径向生长动态,并从机理上解释树木生长对气候的响应。树木径向生长测量仪是一种常用的连续监测方法,其原理是通过安装感应探头监测树木的径向变化,具有分辨率高、稳定性和可操作性强、对树木伤害小等优点[9],因此我们可以通过高分辨率的数据获得天或季节尺度上树干径向生长对气候的响应规律。
本研究以我国北方半干旱区内蒙古赛罕乌拉国家级自然保护区为研究区域,针对研究区内广泛分布的山杨次生林开展树木径向生长的日尺度和季节尺度的动态监测,并分析树木径向生长对气候因素的响应。本研究旨在从树木个体生长角度揭示北方半干旱区山杨次生林的日尺度和季节尺度上的动态变化规律,明确山杨的生长过程及其影响因子,进一步在气候变化背景下为半干旱区树木生长机制提供新的认识,为区域森林的可持续发展提供科学依据。
1 研究地概况
研究地位于内蒙古赛罕乌拉森林生态系统国家定位观测研究站山杨次生林内,地处内蒙古赛罕乌拉国家级自然保护区(118°18′~118°55′E,43°59′~44°27′N)。研究区属温带半湿润温寒气候区,年降水量约为400 mm,降水集中在6—8月。2021年最高气温达34 ℃,最低至-32 ℃;大兴安岭南段是草原向森林、东亚阔叶林向大兴安岭寒温带针叶林双重交汇的过渡地带,是典型的半干旱区域。本研究区域在山杨次生林内,伴生白桦(Betulaplatyphylla)、蒙古栎(Quercusmongolica)、黑桦(Betuladahurica)等树种;林下灌木有土庄绣线菊(Spiraeapubes-censTurcz.)、虎榛子(OstryopsisdavidianaDeca-isne)等[5];土壤类型以山地黑土、灰色森林土、棕壤土和暗栗钙土为主[10]。

表1 试验地山杨样木的基本情况
2 材料与方法
2020年7月在内蒙古赛罕乌拉森林生态系统国家定位观测研究站山杨次生林内布设30 m×30 m的样地,在样地内按平均木法选择4棵样树(表1),在其胸径位置(1.3 m高)安装点触式树木径向生长仪(德国Ecomatik公司的dendrometer),为了避免树皮膨胀收缩过程对数据准确性的影响,在安装前需要除去胸径处的部分树皮;在4棵样树周围选择1棵树在1.3m高位置安装大气温湿度记录仪(美国Onset公司的HOBO pro v2 U23),在附近选择地势平缓处挖一个深度为90 cm的垂直剖面埋放土壤温湿度记录仪(美国Decagon公司的Em50/G土壤水分数据采集器),观测深度为10、20、40、60、90 cm,每隔30 min记录1次数据,记录时间为2020年8月22日—2021年9月3日。
气象数据中的空气温度(Ta)、相对湿度(HR)来自山杨次生林样地布设的大气温湿度记录仪;土壤温度和土壤含水量来自山杨次生林样地布设的土壤温湿度记录仪,本研究只取用深度为40 cm的土壤温度和土壤含水量用作分析;降雨来自赛罕乌拉生态站综合气象站观测场;饱和水汽压差[11](DVP)根据空气温度(T)和相对湿度(HR)计算:
DVP=(1-HR)×0.610 8×exp[17.27/(T+273.3)][11]
本研究所用的是2020年8月—2021年9月观测的树干径向生长数据和气候因子数据。径向生长数据采用最大值法[12]获得径向生长的信息,即从每日所获得的48个原始树木茎干数据中提取每日径向数据中的最大值,计算连续两日最大值的差值,得到日变化的数值;将每日记录的起始值记为0,每日记录的48个值与每日的起始值相减,分析树木径向生长的日循环模式。采用日平均值法[12],提取树干的径向累积变化量。因采集到的数据较为离散,无法排除水分对山杨在某一时间点生长速率的干扰,所以利用Gompertz生长模型对径向累积变化量进行函数拟合,模拟树木的生长过程[13]。对Gompertz函数进行一阶求导得到树木的生长速率曲线,生长速率大于(4 μm·d-1)的时期确定为树木的主要生长时期[8,14],即树木持续生长的时间,Gompertz函数式中(Y为树干径向累积变化量;Y0为下渐近线,代表树木生长季开始时的初始状态;A为上渐近线;β为与Y初始值有关的参数;k为生长率;t为年积日。)
Y=Y0+A×exp[-exp(β-kt)]。
数据采集器内收集到的数据在Excel内进行整理,利用Origin、SPSS进行数据分析、绘图制作。
3 结果与分析
3.1 山杨茎干的日变化特征
山杨茎干日动态变化见图1,山杨2020年10月—2021年9月的茎干日动态变化在生长季节与非生长季节的振幅具有反相位的变化特征,有明显的季节性特点。山杨茎干日动态在生长季(5—9月)表现为在白天达到谷值、在夜间达到峰值,其中6月份的振幅最大;在非生长季(2020年10月—2021年4月份)则表现为在白天达到峰值,在夜晚达到谷值,2、3月份的振幅最大,在4月振幅显著减小。在生长季山杨茎干日动态最大值出现在上午06:00—09:00,日最小值出现在下午14:00—17:00;在非生长季,山杨茎干日动态最大值出现在下午15:00—18:00,日最小值出现在上午06:00—09:00。由于2020年12月—2021年1月中旬日最高温与日最低温均在0 ℃以下,昼夜温差小,振幅较小。因此,2020年12月—2021年1月的茎干日动态变化幅度与其他非生长季月份相比较小。

(a)生长季山杨茎干日变化特征 (b)非生长季山杨茎干日变化特征
3.2 山杨茎干的季节变化特征
从山杨茎干变化的累积曲线来看(如图2),4棵样树的变化趋势一致,茎干变化在春季(4月上旬—5月中旬)迅速增加,山杨启动生长;在夏季在(5月中旬—7月下旬)树木进入快速生长期,在秋季(8月上旬—11月中旬)径向生长曲线平稳波动;在冬季(11月中旬—4月上旬)生长曲线有明显波动且缓慢降低的特点。
利用Gompertz函数对树木径向生长数据进行拟合(图2a),生长速率超过4 μm·d-1的时间认为是持续生长的时间。拟合结果显示(图2b),4棵样树开始生长的时间为第(132.25±2.26)d(5月12日左右);最大生长速率出现在第(156.75±3.10)d(6月5日左右),生长速率为(25.79±6.18)μm·d-1;生长结束时间为202.25±10.81 d(7月21日左右),平均生长时长为70 d。
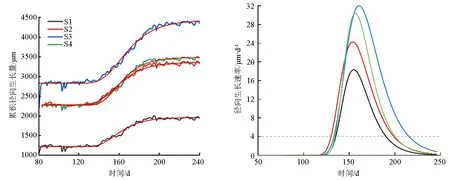
(a)山杨累积径向生长曲线拟合 (b)山杨径向生长速率
3.3 山杨茎干径向变化与环境因子的关系
通过分析山杨茎干径向变化与空气温度、土壤温度和土壤水分等气候因子的关系发现,不同时期影响山杨茎干径向变化的气候因子不同。在春季(4月上旬—5月中旬)山杨进入生长季之前,山杨茎干径向变化主要受土壤温度和土壤含水量的影响(R=0.526,P<0.001)(表2);进入生长季后(5月中旬—7月下旬),茎干径向变化主要受到温度(空气温度、土壤温度)、降水和饱和水汽压差的显著影响(图3)。

表2 不同生长阶段山杨树木径向变化与环境因子的Pearson相关系数

图3 山杨树木茎干变化与气候因子的季节动态
4 讨论
4.1 山杨茎干径向日变化模式
本研究结果发现我国北方半干旱区山杨的茎干日动态变化在生长季和非生长季出现反相位模式,在生长季表现为在白天达到谷值、在夜晚达到峰值,在非生长季表现为在白天达到峰值,在夜晚达到谷值。在生长季,空气温度逐渐升高,树木的蒸腾作用引起树木失水,树木通过根系吸水来维持细胞间的水分平衡,使茎干日动态变化出现了白天收缩和夜间膨胀的现象[15-17]。在非生长季环境温度逐渐降低,树干的冻融作用导致树木茎干日动态发生了变化,出现白天膨胀、夜间收缩的现象。夜间由于温度持续降低,细胞外的水开始冻结,此时细胞内的水分由于渗透作用转移到木质部细胞,从而使细胞体积减小并最终导致茎干收缩,白天温度上升,这个过程被逆转,水分流回细胞,导致树干扩张达到每天的最大值。所以导致山杨茎干日动态在生长季与非生长季出现反相位模式[12]。
前人的研究表明,树木在夏季的日动态变化振幅要高于冬季[18],但本研究区域的山杨日动态变化在冬季的变化振幅要高于夏季,这是由于该研究区季节温差大,导致山杨在秋冬季节的日动态变化振幅要高于夏季[19]。孟盛旺等[20]人对杉木的研究结果表明树木茎干日动态变化也可以反映出树木的水分利用状态与光合能力。还有研究表明,在生长季茎干日动态变化模式受水分(降水、土壤水分)的控制,这也反映出水分在“根系-土壤”运输中的平衡状态[21,35],而在非生长季茎干日动态变化模式与空气温度密切相关,综上可知水分和温度是影响本研究区山杨茎干日动态变化的主要因素。
4.2 山杨茎干径向季节变化模式
在季节尺度下山杨茎干径向变化具有明显的季节特征。在冬季(2020年10月上旬—2020年4月上旬),山杨茎干径向变化在2020年10月上旬—2021年2月上旬呈持续下降的趋势,在2—3月上旬出现强烈的径向波动现象,随后生长曲线趋于平稳。这是由于在生长季末期,温度保持在0 ℃以下,树木中储存的水分冻结,造成细胞内外形成水势差,细胞体积减小,导致树木茎干收缩[17,22,36]。在2—3月出现强烈的径向波动现象主要是由于该阶段内气温逐渐上升,白天气温会高于0 ℃,树干冻结的液体会融化,使树干径向迅速增加,当夜晚气温降至0 ℃以下,解冻的水分会重新冻结,造成树干径向迅速收缩,频繁的冻融造成了这段时期内剧烈的茎干径向收缩现象。Zweifel et al.[23]在研究中指出在冬季,由于温度的变化,霜冻收缩和解冻膨胀交替进行,导致茎干变化非常剧烈。
在春季(4月上旬—5月中旬),随着早春期间的气温上升,土壤温度也逐渐升高,土壤含水量回升,并在5月8日达到最大值,此时树木茎干组织的水分得到补充,这一过程为形成层细胞分裂和伸长提供动力[24,37]。但此时的土壤温度仍处在0 ℃左右,并未达到树木生长所需的环境条件,树木未开始生长。董满宇等[25]对芦牙山林线华北落叶松的研究结果表明,土壤低温会抑制树木的光合作用,影响树木径向生长。土壤低温也会影响树木根系对水分的吸收,抑制形成层细胞的活力,无法保证树木的正常生长[26]。
在夏季(5月中旬—7月下旬),山杨在5月中旬进入生长期,在5月10日之后,空气温度上升,春季积雪融化为树木早期生长提供了充足的水分,土壤温度保持在0 ℃以上,形成层活动开始,山杨开始生长。孟盛旺等[20]对杉木径向生长动态的研究表明,温度是影响生长季初期木质部活动的主要因素,春季升温也会导致树木形成层活动时间提前。山杨的径向生长主要集中在6—8月,生长速率在6月上旬达到了最大值,径向生长量也在8月上旬有明显的增加。该研究区在6月中旬出现了连续降水,此时的外界环境条件对早材的形成起主导作用。高佳妮等[27]在对贺兰山油松的研究中发现,生长季早期的降水为树木生长提供丰富的水源,保证树木的正常生长,生长季中期的降水也对树木生长具有同样的意义。
在秋季,8月上旬开始,径向生长曲线趋于平缓,此时形成层细胞生长缓慢[28,38]。在生长季后期,细胞壁木质化过程完成[29,39]。在9月份之后,空气温度、土壤温度逐渐降低,降水量减少,树木生长趋于停止。
4.3 山杨茎干径向变化与环境因子的响应关系
山杨茎干径向变化在生长季早期主要受到土壤温度和土壤水分的显著影响,生长季前期土壤水分是山杨径向变化的主要影响因素。山杨径向变化在进入生长季之前,树木茎干组织水分通过土壤水分得到补充,使茎干组织中的水分含量满足了生长开始的需要[30]。山杨在进入生长季后,温度(土壤温度、空气温度)和降水对山杨的径向生长呈显著正相关,与饱和水汽压差呈显著负相关。大量的研究表明,温度和降水是形成层活动开始的主要气候因素[31],升温有利于植物体内的非结构性糖转化为糖类为细胞生长提供能量基础[32,40]。该研究区降水集中在6—8月,降水是补充生长在干燥的有机层上的树木内部水储备的一个重要因素[19]。对大兴安岭南段山杨径向生长与气候变化的研究结论也表明生长季降水对山杨径向生长有积极作用,而春季降水会促进树木形成宽年轮[5]。前人对北方森林的树木气候学研究表明,初夏的降水对树木径向生长有积极的影响[33-34]。
5 结论
我国半干旱地区的山杨茎干日动态变化,在生长季和非生长季出现反相位模式,在生长季,山杨茎干日动态变化是由空气温度上升,树木蒸腾作用增强而引起的;在非生长季,山杨茎干是由冻融作用引起的日动态变化;水分和温度是影响本研究区域内山杨树干径向生长日动态变化的主要因素。山杨茎干径向变化在生长季前期受到土壤水分的显著影响,在进入生长季之后,山杨径向生长在降水和温度(土壤温度、空气温度)的共同作用下生长,而饱和水汽压差则在生长季显著限制了山杨的径向生长。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
西北林学院学报(2022年5期)2022-10-04
土壤(2022年1期)2022-03-16
林业科学(2021年5期)2021-07-13
中学生数理化·高一版(2021年3期)2021-06-09
航空发动机(2021年1期)2021-05-22
林业科学(2021年1期)2021-03-13
棉花学报(2020年3期)2020-08-08
农业机械学报(2020年3期)2020-04-01
江苏农业科学(2017年21期)2017-12-13
