
厌氧甲烷氧化及其影响因子的研究进展
2023-02-27 07:14余子洁郭大陆李致同罗红燕
贵州大学学报(自然科学版) 2023年1期
余子洁,郭大陆,李致同,罗红燕
(西南大学 资源环境学院,重庆 400715)
甲烷是一种比二氧化碳温室效应强约28倍的温室气体[1],它在环境系统中的地位对地球气候有着重要的影响。厌氧甲烷氧化(anaerobic oxidation methane,AOM)是一个全球重要的微生物过程,它阻止了自然沉积物中相当大比例的甲烷在大气中释放(在某些环境中85%)[2-4]。联合国政府间气候变化专门委员会报告指出全球稻田土壤排放的甲烷约为35~56 Tg,约占全球甲烷排放总量的25%[2]。过去,研究学者都认为甲烷的主要代谢途径是好氧氧化反应,但自从1976年REEBURGH[3]首次在卡里亚科海沟水域的沉积物中发现AOM反应后,AOM反应开始被广泛研究[4-5]。随着研究逐步深入和拓展,越来越多种类的AOM反应被发现,同时AOM反应也被证实是控制缺氧环境中甲烷通量的关键[6]。基于此,本文对AOM类型、参与过程的微生物种类以及甲烷浓度等影响AOM的因子作一综述,以期为进一步研究AOM过程在碳、氮、硫及金属离子循环以及环境污染中的重要意义提供参考。
1 AOM类型及参与反应的主要微生物分类
1.1 硫酸盐还原型厌氧甲烷氧化反应

(ΔG=-16.6 kJ/mol)
(1)
参与硫酸盐还原型厌氧甲烷氧化反应的微生物主要是一类隶属于广古菌门的厌氧甲烷氧化古菌(Anaerobic methanotrophic archaea,ANMEs),通常分为3类:ANME-1,ANME-2a、ANME-2b、ANME-2c及ANME-3[12]。通过对厌氧甲烷氧化古菌16S rRNA和mcr A基因序列的多样性检测,发现这3类古菌彼此之间进化距离较远,基因序列相似度为75%~92%[13]。ANME-1常发现于深海、火山口等区域,与甲烷八叠球菌(Methanosarcinales)和产甲烷微菌(Methanomicrobiales)相似[12]。ANME-2常被检测于冷泉、垃圾填埋场、湿地、湖泊沉积物及稻田等地方,隶属于甲烷八叠球菌 (Methanosarcinales)[4]。ANME-3在极端环境如冰原等地被检测到,与产甲烷菌( Methanococcoides )相似[4]。
1.2 反硝化型厌氧甲烷氧化反应

(ΔG=-765 kJ/mol)
(2)
(ΔG=-928 kJ/mol)
(3)
参与反硝化型厌氧甲烷氧化反应的微生物主要是CandidatusMethylomirabilis oxyfera(M.oxyfera)[15],隶属NC10门菌,形态呈细杆状、杆状,通常以单细胞或团聚体的形式存在。ETTWING等[16-17]利用宏基因组方法检测湿地沉积物内M.oxyfera细菌后,发现可通过该菌内部NO歧化酶直接将NO氧化成N2和O2。NO歧化酶是一种很古老的酶,可当做检测不同生境M.oxyfera细菌的生物标记物[18]。
1.3 金属离子型厌氧甲烷氧化反应
金属离子型厌氧甲烷氧化反应是以Fe3+、Mn4+和Cr6+等金属离子为电子受体耦合AOM。DICHRISTINA等[19]在1988年发现,自然界中有微生物在厌氧环境中,可以利用铁锰氧化物为最终电子受体获取能量。在2009年,BEAL等[20]在加利福尼亚EelRiver流域的河流底部沉积物中发现,有微生物可以在不存在硫酸盐的情况下,实现厌氧甲烷氧化耦合Fe3+、Mn4+的还原[21-22],反应式如式(4)(5)所示。从热力和动力学角度分析,金属离子型比硫酸盐还原型厌氧甲烷氧化反应更容易发生。
(ΔG=-572.157 65 kJ/mol)
(4)
(ΔG=-789.917 65 kJ/mol)
(5)
这一点的发现成功拓宽了AOM的领域。在湿地厌氧环境中,以金属离子为最终电子受体的AOM反应也大量存在。有学者也分别在印度尼西亚Matano湖[23]和西伯利亚Baikal湖[24]底沉积物中均发现金属离子型厌氧甲烷氧化反应。NORDI等[25]在丹麦淡水湖沉积物中发现以Fe3+为最终电子受体的AOM反应。姜怡如等[26]推测金属离子型厌氧甲烷氧化反应更容易发生在淡水区域或湿地区域。LU等[27]通过16S rRNA基因测序观察发现,在反应器内发现加入Cr5+离子至还原后,ANME-2古菌数量大幅度增加,其中甲烷八叠球菌和红球菌数量增加显著,逐渐成为AOM反应的优势菌群。但目前未有报道表明这两种菌具有还原铬的能力,因此推测ANME-2可还原Cr5+或者可耦合其他还原菌还原Cr5+。从现有的研究结果分析,金属离子型厌氧甲烷氧化反应中所参与的微生物主要是ANME-1、ANME-2、ANME-3及脱硫单胞菌属等[28]。
2 AOM的影响因子
AOM是在无氧或者厌氧条件下,将CH4氧化成CO2的过程。影响AOM的因子有很多,本文结合国内外对AOM的研究进展,主要对甲烷浓度、氧化还原电位、温度、pH值及盐度等几个因子进行了总结。
2.1 甲烷浓度
CH4是发生AOM的基础,是AOM唯一的电子供体,适当提高甲烷浓度对AOM有着促进的作用。PIMENOV等[29]研究结果表明,CH4浓度增加至640 μL/L时,AOM反应速率比未添加样品高出195倍,后有研究表明AOM反应的最适甲烷浓度为1.4 mmol/L[30]。ORCUTT等[31]在墨西哥海湾研究中发现,甲烷浓度最大值区域与厌氧氧化峰值出现的区域相同。SINISCALCHI等[32]也证实提高甲烷浓度可以促进AOM的发生。SHEN等[33]通过对还原性河床进行实验发现,AOM途径的多样性与甲烷浓度有关,甲烷浓度高时,AOM多样性较大。张佩等[34]通过实验证明水合物沉积物中溶解的甲烷浓度高于有机质分解产生的甲烷浓度,因此水合物沉积物中AOM反应速率较快。一般情况,甲烷浓度对AOM反应具有一定的促进作用。
2.2 氧化还原电位
AOM是发生在厌氧的状态下,依靠厌氧微生物进行的反应,因此对氧气浓度有严格的要求,LUESKEN等[35]研究表明氧气的存在会抑制AOM。袁梦冬等[36]对M.oxyfera的cDNA序列分析发现氧气浓度会抑制好氧甲烷氧化过程及反硝化NO还原酶NorZ2基因等。XU等[37]研究发现,不同深度的湖底沉积物检测出功能基因主要分布在深层,而少量分布在浅层。由于浅层区域的湖水更接近于大气,与空气进行频繁的气体交换,湖水中氧容量较高,氧化还原电位较高。HE等[38]研究发现低氧气含量的深层土壤中,M.oxyfera丰度较高于表层土壤。YANG等[39]检测各项多样性指数也存在氧气含量较低的深层样点要高于表层和浅层样点的现象。但LOU等[40]利用厌氧甲烷氧化微生物处理含氧地下水中硝酸盐的研究发现,以亚硝酸盐为电子受体的厌氧甲烷氧化细菌在氧气受限的条件下可以实现相对丰度的增加,而以硝酸盐为电子受体的厌氧甲烷氧化古菌则会被淘汰,推测以亚硝酸盐为电子受体的厌氧甲烷氧化细菌对氧气有耐受性。因此,氧化还原电位的高低对AOM的影响理论尚待完善。
2.3 温度
温度的高低会直接影响微生物体内酶的活性,进而影响微生物的代谢能力。在一定的温度下,升高温度有利于AOM反应的进行。唐荧霜等[41]通过实验证明温度在25~35 ℃时,温度升高会提高反应速率,温度达到35 ℃左右,AOM反应速率最大,而温度到达45 ℃时,AOM反应速率最小。DEUTZMANN等[42]在4 ℃的湖泊沉积物中检测到N-DAMO,表明厌氧甲烷氧化菌对低温有一定的适应性。温度也影响着底物的传输效率。甲烷在水中的溶解度大小受温度的影响,理论上温度越低甲烷在水中的溶解度越高,AOM底物浓度越大。
2.4 pH值
不同pH值条件下的AOM反应速率不同。目前已有研究表明,AOM富集培养最适合的pH值为6.0~8.0,但在pH值<5.0或pH值>9.0的环境中也可以检测出有相关功能基因的微生物[43]。ZHU等[43]和HE等[44]均认为,pH值为7.6左右是AOM最适pH值,此时厌氧甲烷氧化菌的活性最强。赵荣等[45]发现调控pH值至8.5和9.5的碱性环境,此时反硝化厌氧甲烷氧化速率明显降低。基于此,本文推测厌氧甲烷氧化微生物在参与反应过程中,菌体内部酶如甲醇脱氨酶、亚硝酸盐还原酶和NO歧化酶等更适宜中性环境。
2.5 盐度
通过对厌氧甲烷氧化菌富集培养后,HE等[44]发现反硝化型厌氧甲烷氧化反应最佳盐度值为20 g/L,盐度超过20 g/L时,AOM反应活性会明显受到抑制作用。唐良港等[46]的实验研究发现,随着盐度从0 g/L逐渐升至15 g/L时,厌氧甲烷氧化菌对亚硝酸盐的还原速率明显降低。在较高的盐度环境中,甲烷的溶解量会降低。高盐环境也影响参与厌氧甲烷氧化微生物体内多种代谢酶的状态,如甲醇脱氨酶、亚硝酸盐还原酶等[46],从而影响菌群的代谢活性。厌氧甲烷氧化菌对盐度有良好的耐受能力,但浓度过高仍会对菌群产生抑制作用。
3 检测AOM的现代技术方法
传统微生物多样性研究方法是将微生物从环境中分离后再于实验室内培养得到单菌株,对培养环境和仪器设备有较高要求,研究过程困难且获得菌株极少。闵航等[47]首次在浙江象山市郊青紫泥水稻田土壤中分离到1株能独立厌氧氧化甲烷的菌株,但有研究表明富集培养有限生境中甲烷厌氧氧化菌并不能充分描述甲烷厌氧氧化菌的多样性[48]。随着科技的不断创新发展,现代分子生物学技术打破了传统室内培养的局限性,从分子水平客观地揭示不同功能基因的厌氧甲烷氧化菌群的物种多样性,有效地克服传统培养方法的不足,提高了分析检测的速度及结果的准确性和完整性。
现代分子生物学技术包括荧光原位杂交技术、变性/温度梯度凝胶电泳、高通量测序及宏基因组学等[11,49]。荧光原位杂交技术可建立克隆库,在群落水平对环境中厌氧甲烷氧化菌物种进行鉴定、分析种群数量,操作简便,测定灵敏。BOETIUS等[8]通过荧光原位杂交技术首次观察到厌氧甲烷氧化古菌与硫酸盐还原菌的共生体,并证明了其共生关系。RAGHOEBARSING等[14]同样应用荧光原位杂交技术鉴定了厌氧甲烷氧化菌与反硝化菌的共生体。
随着研究深入,许多研究者将16S rRNA基因为标记基因,探究了不同环境中厌氧甲烷氧化古菌。16S rRNA基因测序可从群落水平角度分析和鉴定未知的厌氧甲烷化菌及其系统发育关系,16S rRNA相比于5S rRNA和23S rRNA,核苷酸数量适中、遗传信息较多且应用较广泛[49]。朱群等[48]采用基因克隆文库技术和实时荧光定量聚合酶链反应技术考察了西湖底泥中反硝化厌氧甲烷氧化微生物,16S rRNA基因测序结果表明西湖底泥中有此类微生物。
宏基因组学是对环境中微生物的总基因组DNA进行研究,全面系统分析其组成、分类和发展进化规律,可提高发现新物种的概率。ARSLAN等[50]通过对AOM的富集培养发现一种类似Methanoperedens古菌的基因组和转录组。现代分子生物学检测方法的广泛应用为研究自然生境中厌氧甲烷氧化菌的多样性提供了技术支持,进一步促进了厌氧甲烷氧化菌多样性及生态功能方面的研究[49]。
4 未来展望
AOM是全球气候变化和元素循环的关键环节,充分了解硫酸盐还原型、反硝化型及金属离子型等AOM的反应机理、代谢途径和实际应用,有助于深入认识自然界物质循环过程,对缓解全球气候变暖和加强全球地球化学循环具有一定的积极作用。目前,对于AOM反应研究仍处于初步阶段,后续仍有很多问题有待解决。
1)拓宽研究区域。硫酸盐还原型厌氧甲烷氧化反应过程释放能量较少,且多发现于硫酸盐和甲烷交换地带,是否也广泛分布于淡水区域?近些年来由于过度使用氮肥,部分硝酸盐和亚硝酸盐随着雨水渗透入土壤,使其在水稻田淹水层聚集。由于水稻田是甲烷汇,会释放大量甲烷,因此为反硝化型厌氧甲烷氧化反应提供良好的环境,并且仍有部分氮肥随着地表径流汇入河流,在河流底部沉积物中聚集。由于沉积物中有机质含量较高,因此反硝化型厌氧甲烷氧化反应也很容易发生。流失的氮肥通过河流、地下水等汇入海洋是否对入海口等海洋区域AOM产生影响?金属离子型厌氧甲烷氧化反应作为AOM的新类型,当前研究较少,未来可进一步对金属离子型厌氧甲烷氧化反应体系中,种间电子转移机制和胞内外电子与金属离子之间的转移机制进行研究。
2)寻找更多电子受体。目前,AOM过程可耦合硫酸盐、亚硝酸盐、硝酸盐及铁锰等金属离子,是否还存在其他潜在的电子受体与其耦合?这些潜在受体的发现可拓宽目前对AOM的认识。
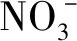
4)获取纯培养物。至今很多学者对厌氧甲烷氧化微生物进行富集和纯培养,但获取物仍较为有限[54]。为获取纯培养物,可进一步采用生物电化学的方法对微生物进行富集,确保得到较高富集度培养物或者纯的培养物。
总之,AOM无论是在自然生境还是在实际应用中,都具有显著的环境生态效益。因此,AOM的相关研究仍是未来碳、氮、硫及金属离子循环的研究热点,可深入探究自然环境中的AOM以及其在实际环境工艺中的应用。
猜你喜欢
土壤学报(2022年3期)2022-08-26
大自然探索(2022年5期)2022-07-11
知识就是力量(2022年6期)2022-06-16
当代医药论丛(2021年2期)2021-02-24
石油沥青(2018年3期)2018-07-14
环境保护与循环经济(2017年7期)2018-01-22
保健文汇(2017年4期)2017-11-03
环境保护与循环经济(2017年10期)2017-03-16
中国环境科学(2016年3期)2016-02-08
应用海洋学学报(2014年2期)2014-11-26
