
白菜TLP基因家族鉴定及表达分析
2023-03-06 07:09王紫雯陈雨吴晓宇赵玉梅任鑫如吕金枝李嘉利段巧红黄家保
山东农业科学 2023年1期
王紫雯陈雨吴晓宇赵玉梅任鑫如吕金枝李嘉利段巧红黄家保
(山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东 泰安 271018)
TLP(Tubby-like protein)最早在肥胖小鼠中发现,并且广泛存在于真核生物中[1]。植物TLP蛋白C末端通常含有一个由270个氨基酸组成的Tubby保守结构域,该结构域由一个包含12条反向平行链和一个中央疏水的α螺旋组成,能与特定的磷脂酰肌醇4,5-二磷酸(PIP2)结合,从而与质膜连接[2,3]。与保守的C末端Tubby结构域相反,TLP的N端在不同物种间存在较大差异,动物TLP蛋白N末端高度分化,大多数植物TLP蛋白N末端含有一个保守的F-box结构域,该结构域同蛋白泛素化密切相关[4,5],且有研究表明Tubby结构域和F-box结构域是协同进化的[6]。
动物中TLP基因的生物学功能已经得到深入研究,例如,在人体中发现TLP基因突变可能会导致肥胖、视力丧失和神经感觉丧失等[7,8]。在植物中发现了更多的TLP基因,但是研究较少,许多功能机制仍不清楚[9]。已有的研究更多集中在植物对非生物胁迫的响应,目前已在拟南芥、水稻、番茄、玉米、杨树中分别鉴定出了11、14、11、15、11个TLP基因家族成员[6,10-12]。通过酵母双杂交试验发现,拟南芥TLP家族成员At-TLP3参与了种子萌发过程中脱落酸(ABA)信号的调节,AtTLP9在盐胁迫与干旱胁迫中发挥作用[10,13]。在鹰嘴豆中,CaTLP1在脱水、高盐、脱落酸(ABA)处理下表达上调,过表达CaTLP1可以提高转基因烟草植株对脱水、盐胁迫和氧化胁迫的耐受性,同时改善植株地上部和根构型,其在细胞核中的优先表达可能与抗逆性增强有关[14]。过表达MdTLP7基因可以显著提高拟南芥幼苗对渗透胁迫、盐胁迫、冷热胁迫的耐受性[15]。这都证明了TLP基因参与了植物的非生物胁迫反应,在植物应对非生物胁迫响应的过程中发挥重要作用。
白菜(Brassica rapa)为十字花科芸薹属蔬菜,是中国、韩国和日本等国的主要蔬菜之一,在世界各地广泛种植。目前,土壤盐碱化成为制约我国农业可持续发展的重要因素之一,我国部分白菜产区土壤盐渍化严重影响白菜的正常生长发育过程,因此,亟需培育耐盐、抗盐的白菜品种。现有研究表明,TLP家族在植物对非生物胁迫的响应中发挥一定作用,所以本研究对白菜TLP家族进行了鉴定,并对其基因结构及系统发育等系列生物信息学进行了分析,同时,对其组织表达模式及盐胁迫响应情况进行了研究,以期为其后续功能研究及种质改良奠定基础。
1 材料与方法
1.1 白菜TLP家族成员鉴定及理化性质分析
根据拟南芥已经鉴定出来的11个TLP成员蛋白序列,通过双向Blast在白菜基因组搜索候选白菜TLP家族成员,再通过保守结构域分析等方法进一步确定候选基因。应用软件TBtools[16]和Expasy(https://www.expasy.org/)[17]分析白菜TLP家族的蛋白质分子量(MW)、等电点(pI)等理化性质。
1.2 白菜TLP基因家族染色体定位、共线性分析及进化树构建
从白菜数据库下载gff3基因组注释数据文件,使用TBtools绘制TLP基因在染色体上的分布图。利用MCScanX[18]绘制白菜与拟南芥TLP基因家族共线性分析图。在Ensembl Plants(http://plants.ensembl.org/index.html)获取番茄及水稻TLP家族氨基酸序列,通过MEGA-X[19]采用最大似然法(Maximum Likelihood Estimate,MLE)构建白菜、拟南芥、番茄及水稻TLP家族系统进化树。
1.3 白菜TLP基因结构和保守结构域分析
使用TBtools绘制白菜TLP基因结构图;通过MEME(http://meme-suite.org/)[20]预测白菜TLP蛋白保守基序;利用软件DNAMAN对白菜TLP序列进行比对,分析其保守氨基酸位点。
1.4 顺式作用元件分析
通过Ensembl Plants[21]查找并下载白菜TLP基因家族的起始密码子上游2 000 bp序列,使用PlantCARE(http:/bioinformatics.psb.ugent.be/webtools/plantcare/htm1/)在线网站[22]对获取的序列进行分析。
1.5 白菜TLP基因表达分析
试验材料为山东农业大学蔬菜生长与生殖研究组植物培养室的‘848蛋黄白’,于2022年2月20日定植,分别取定植70 d白菜幼苗的根、茎、叶和当天开放的花、果荚进行RNA提取,用于组织特异性表达研究;使用150 mmol/L NaCl分别处理0、2、4、6、12 h,各部位及各时间段均取3个生物学重复,进行基因表达分析。
通过qPrimerDB-qPCR Primer Database(https:/biodb.swu.edu.cn/qprimerdb/)[23]查找并设计引物(表1),内参基因为BrActin7,由青岛擎科梓熙生物技术有限公司合成。采用RNA提取试剂盒提取白菜样品的总RNA,参照HiScriptⅡQ RT SuperMix for qPCR(南京诺唯赞生物科技有限公司)试剂盒产品说明书进行反转录得到cDNA,用ChamQ Universal SYBR qPCR Master Mix(南京诺唯赞生物科技有限公司)试剂盒进行定量PCR试验。反应体系(20μL):上、下游引物各0.4 μL,SYBR 10μL,cDNA 2μL,用灭菌ddH2O补至20μL。PCR反应程序:95℃30 s;95℃10 s,60℃22 s,40个循环。数据使用Microsoft Excel进行整理,并使用TBtools进行热图绘制。

表1 白菜TLP基因序列引物
2 结果与分析
2.1 白菜TLP基因家族成员鉴定及理化性质分析
经鉴定得到14个白菜TLP基因家族成员,命名为BrTLP1~BrTLP14(表2),这些基因不均匀地分布在白菜10条染色体中的7条,BrTLP1、BrTLP8、BrTLP13未在染色体中发现,根据亚细胞定位预测其可能分布于叶绿体或线粒体中。

表2 白菜TLP基因家族信息
白菜TLP家族蛋白分子量介于38 735.69~52 747.34 D,等电点介于9.16~9.88。所有TLP家族成员的等电点均大于7.0,说明其均富含碱性氨基酸。
2.2 白菜TLP蛋白保守结构域分析
对白菜的14个TLP蛋白序列进行保守性分析,每个基因均含有Motif 2、Motif 3、Motif 7、Motif 10。除了BrTLP14,其他基因的Motif分布都比较均匀。Motif 9仅在BrTLP3、BrTLP4、BrTLP5上出现,BrTLP14中Motif的数目最少,与其他基因成员差异明显,推测该基因在功能上与其他基因存在较大不同(图1)。

图1 白菜TLP基因家族结构域分析
2.3 白菜TLP基因家族结构分析
在鉴定到的14个BrTLP基因中,BrTLP14含有9个CDS,绝大部分BrTLP基因含有4~5个CDS(图2)。BrTLP14的CDS数目与其他基因存在较大差别,推测其可能在白菜器官中发挥特异性功能。对蛋白的保守功能域进行分析,发现绝大多数白菜TLP家族基因都含有F-box结构域,其中Motif 4序列为F-box结构域序列信息。4个保守功能域分别对应Motif 4、Motif 6、Motif 5、Motif 1保守基序(图3)。

图2 白菜TLP基因家族结构分析

图3 白菜TLP家族氨基酸序列比对
2.4 白菜与拟南芥、番茄、水稻TLP家族系统发育分析
由系统发育树(图4)可知,TLP家族被划分为3个亚族,其中亚族A包括1个BrTLP、1个AtTLP和1个OsTLP,亚族B包括3个BrTLP、2个AtTLP、4个SlTLP和1个OsTLP,亚族C包括10个BrTLP、8个AtTLP、7个SlTLP和12个Os-TLP。白菜和拟南芥、番茄、水稻的TLP家族成员进化关系较近,推测其直系同源基因之间可能存在相似的功能。

图4 白菜、拟南芥、番茄、水稻TLP家族进化树
2.5 白菜TLP基因家族染色体定位及与拟南芥基因的共线性分析
通过TBtools工具对白菜与拟南芥的TLP基因家族成员进行共线性分析(图5),结果显示,有10个白菜TLP家族成员在拟南芥TLP家族中找到了对应的直系同源基因,表明TLP基因家族在白菜与拟南芥间有较高的保守性。
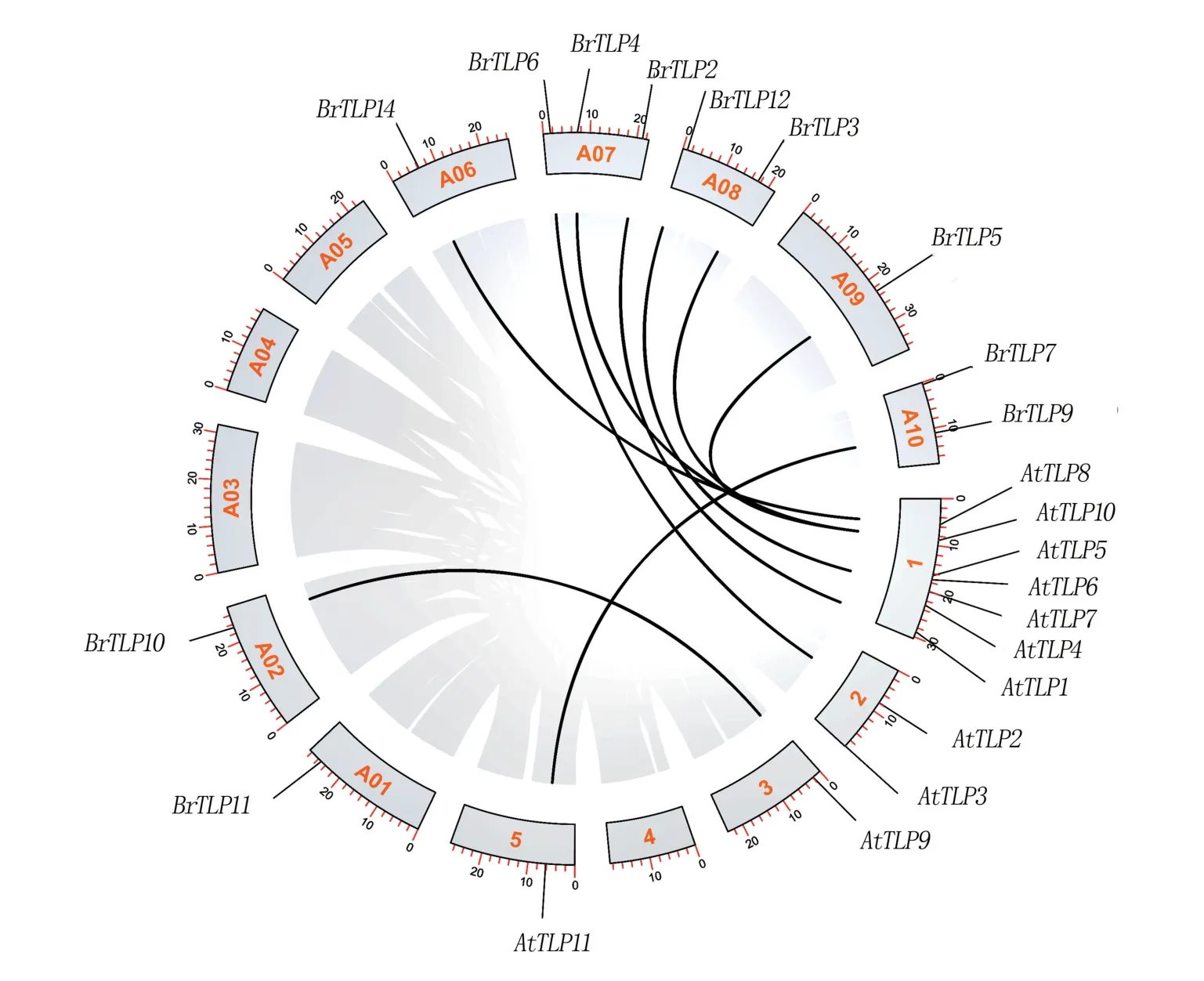
图5 白菜与拟南芥TLP基因家族共线性分析
根据基因位置信息对14个BrTLP基因家族染色体进行定位分析(图6),结果显示其不均匀分布在大白菜7条染色体上,其中7号染色体上的基因最多,有3个成员;其次是8号、10号染色体上有2个基因成员;1号、2号、6号、9号分别各有1个基因成员。

图6 白菜TLP基因在染色体上的分布
2.6 白菜TLP基因家族顺式作用元件分析
通过对白菜TLP基因家族启动子上游2 000 bp的序列进行顺式作用元件分析(图7)发现,14条白菜TLP基因中均含有光响应元件,除了BrTLP9、BrTLP11、BrTLP14,其他11个基因均含有脱落酸响应元件,且BrTLP12最多;除了BrTLP4、BrTLP5、BrTLP10、BrTLP12、BrTLP13,其他基因均含有赤霉素响应元件和生长素响应元件;除了BrTLP7,其他基因均含有厌氧响应位点,且BrTLP14含量最多,推测其可能在厌氧逆境胁迫中起重要作用。除了BrTLP4、BrTLP8、BrTLP11、BrTLP14、BrTLP13,其他9个基因均含有茉莉酸甲酯响应元件,6个白菜TLP基因含有水杨酸响应元件和干旱响应元件。

图7 白菜TLP基因启动子顺式作用元件分析
2.7 白菜TLP基因家族亚细胞定位预测
白菜TLP基因家族主要集中在细胞核、线粒体和细胞质基质(表3),其次为叶绿体、细胞质和细胞骨架。细胞核中所有的基因均有定位,在线粒体中仅有BrTLP2、BrTLP3、BrTLP14没有定位信号,仅有BrTLP6定位在内质网中。每个基因在不同细胞器中的表达位置和表达量各不相同,推测这与基因所执行的功能有关。

表3 白菜TLP基因家族亚细胞定位预测
2.8 白菜TLP基因家族表达分析
2.8.1 组织特异性表达分析 以根的表达量为对照,大多数BrTLPs在白菜花中表达量较高,在茎、叶和果荚中的表达量与根相似。其中BrTLP14在果荚、叶和茎中的表达量最高,BrTLP5在花中的表达量最高,同时在花中表达量较高的基因还有BrTLP8、BrTLP10(图8)。

图8 BrTLPs在大白菜不同器官中的表达
2.8.2 盐胁迫下的基因表达分析 由图9可知,随着盐胁迫时间的延长,BrTLP2、BrTLP6两个基因的相对表达量呈现升高的趋势;BrTLP4、BrTLP7、BrTLP10、BrTLP13的相对表达量均在2 h时最高,包括BrTLP5在内均呈现先升后降的趋势;BrTLP1、BrTLP3、BrTLP8、BrTLP9、BrTLP11、BrTLP12相对表达量较低;BrTLP14在盐胁迫12h时表达量达到最高。

图9 盐胁迫下白菜TLP基因家族表达分析
3 讨论与结论
在动植物生长发育过程中,TLP家族在维持细胞正常生理功能及应对胁迫过程中发挥了重要的调控作用。本研究通过双向Blast在白菜基因组中初步完成了白菜TLP家族成员的鉴定,并对其系统进化关系、保守性及表达特性进行了初步分析,为后续探究白菜中TLP基因功能奠定了基础。
白菜与拟南芥在进化上具有较近的祖先,而白菜经历的全基因组三倍化(WGT)事件导致了其与拟南芥在基因组层面上的一些差异。根据WGT理论,白菜TLP基因数量应为拟南芥的三倍左右[24],而本研究结果表明BrTLP家族数量并未出现显著增加。在其他几种植物中发现的TLP基因家族成员数量均为十几个,表明在不同物种之间具有高度的保守性,白菜与拟南芥TLP基因家族的共线性分析也证明了两个物种间显著的共线性关系。在动物中的研究表明尽管Tubby结构域高度保守,但不同TLP基因成员在功能上却无法相互补偿[25],推测在进化过程中TLP家族可能丢弃了部分冗余基因,仅保留了现有的少数成员高度参与众多真核生物生长发育中的调控过程。
基因结构与蛋白序列的解析为相关基因生物学功能的研究奠定了基础。对白菜TLP基因家族结构及保守基序进行分析表明,白菜TLP家族在进化上距离较近成员之间的基因结构具有一定相似性,而与距离较远的成员基因结构存在一定差异,暗示了家族成员在生物学功能上可能存在不同。另外,保守基序分析表明白菜TLP家族成员虽然在基因结构上存在一定差异,但部分序列仍具有很强的保守性。非保守区段氨基酸序列的变化导致了白菜TLP家族在多物种TLP家族进化树中的不均匀分布,这也进一步暗示了该家族不同成员间的生物学功能可能存在较大差异,而在物种间进化距离较近的TLP成员在功能上可能高度保守。
以往研究证明TLP基因家族在植物应对非生物胁迫过程中发挥了重要调控作用。本研究发现绝大多数白菜TLP成员均对盐胁迫有不同程度的响应,其中BrTLP10在盐胁迫发生后的4~12 h表达明显降低,对盐胁迫的响应十分显著。研究表明,拟南芥中的基因AtTLP9在响应盐胁迫及干旱胁迫过程中具有重要作用[13],多物种TLP进化分析的结果表明BrTLP10与AtTLP9在进化上距离最近,另外,在白菜与拟南芥的共线性分析中也表明这两个成员存在一定共线性关系[10]。根据AtTLP10现有的研究推测BrTLP9不仅参与盐胁迫响应,可能也在白菜应对干旱胁迫过程中具有重要作用,但其在盐胁迫下的具体调控机理以及是否在白菜应对干旱胁迫的过程中发挥作用还有待进一步研究。
猜你喜欢
趣味(作文与阅读)(2022年3期)2022-06-10
小读者(2021年6期)2021-11-23
趣味(作文与阅读)(2021年6期)2021-11-06
科学与财富(2021年3期)2021-03-08
国际放射医学核医学杂志(2020年4期)2020-07-27
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13
温州大学学报(自然科学版)(2019年2期)2019-06-04
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
新闻传播(2015年12期)2015-07-18
