
不同波长光照对草地贪夜蛾成虫趋光行为及视蛋白表达量的影响
2023-05-30 11:17刘思敏汪永乾汤金荣张永军付晓伟梁革梅
植物保护 2023年2期
刘思敏 汪永乾 汤金荣 张永军 付晓伟 梁革梅


摘要 為提高诱虫灯对草地贪夜蛾Spodoptera frugiperda的诱集效果,在室内测试了草地贪夜蛾雌雄成虫对390~550 nm 10种不同波长光源的趋光行为反应,以及光刺激对其4种视蛋白基因表达量的影响。结果表明:草地贪夜蛾雌蛾和雄蛾对10种波长光源的趋光率均存在显著差异,其中雌蛾对绿光区510、520 nm和550 nm的趋性较高,趋光率分别为68.5%、65.0%、63.5%;雄蛾对绿光区520 nm及紫光区420 nm的趋性较高,趋光率分别为69.0%和60.5%。雌雄成虫对510 nm 和550 nm的趋光率存在显著的性别间差异,但其他供试波长处理下,雌雄成虫的趋光率均无显著的性别间差异。520 nm光源刺激下,雌蛾和雄蛾均表现为3日龄(雌:65.0%;雄:69.0%)和5日龄(雌:59.1%;雄:61.4%)的趋光率最高,而1日龄(雌:41.4%;雄:24.1%)的趋光率最低。雄蛾受520 nm光刺激20 min后,其紫外光敏感蛋白(UV-opsin)基因的相对表达量显著升高,是对照组的3.61倍;但长波敏感蛋白1(LW-opsin1)、长波敏感蛋白2(LW-opsin2)和蓝光敏感蛋白(B-opsin)基因的相对表达量与对照组相比无显著差异。雌蛾受520 nm光刺激20 min后,其4种视蛋白基因的相对表达量与对照组相比均无显著差异。综上所述,草地贪夜蛾成虫对绿光和紫光较为敏感,羽化3~5 d的成虫趋光性最强,视蛋白基因表达量的改变可能是草地贪夜蛾成虫对光刺激产生行为反应的关键因素。
关键词 草地贪夜蛾; 成虫; 波长; 趋光率; 光刺激; 视蛋白
中图分类号: S 433.4
文献标识码: A
DOI: 10.16688/j.zwbh.2021678
Abstract In order to improve the light-trapping efficacy on Spodoptera frugiperda in the field, the phototactic behavior of male and female moths to 10 light sources with different wavelengths (390-550 nm) and the effect of light stimulation on the expression levels of four opsin genes of S.frugiperda were tested in the laboratory. The results showed that there were significant differences in the phototactic rate of both male and female moths to 10 light sources. The female moths showed relatively higher phototaxis to 510 nm, 520 nm and 550 nm (green light), with a phototactic rate of 68.5%, 65.0% and 63.5%, respectively. The male moths showed relatively higher phototaxis to 520 nm (green light) and 420 nm (purple light), with a phototactic rate of 69. 0% and 60.5%, respectively. For the 10 different light wavelengths tested, there were significant inter-sexual differences in the phototaxis of S.frugiperda moths to 510 nm and 550 nm, but no significant differences to the other wavelengths. The phototaxis of moths at different day-ages to the stimulation of 520 nm light was tested, and the results showed that both female and male moths had the highest phototactic rate at the age of 3 day-old (female: 65.0%; male: 69.0%) and 5-day old (female: 59.1%; male: 61.4%), while the lowest at the age of 1-day old (female: 41.4%; male: 24.1%). When male moths were stimulated by 520 nm light for 20 min, the relative expression level of ultraviolet-sensitive opsin (UV-opsin) gene increased significantly, which was 3.61 times of the control. However, the relative expression level of the other tested opsin genes had no significant difference from the control group, including the long-wavelength-sensitive opsin 1 (LW-opsin1), the long-wavelength-sensitive opsin 2 (LW-opsin2) and the blue-sensitive opsin (B-opsin) genes. When female moths were stimulated by 520 nm light for 20 min, the relative expression level of the four tested opsin genes had no significant differences from the control. In conclusion, S.frugiperda moths had higher sensitivity to green and purple lights, and the highest phototaxis occurred at the age of 3-day and 5-day old after emergence; meanwhile, the changes of expression level of opsin genes might be the key factor affecting the response of S.frugiperda to different light stimulations.
Key words Spodoptera frugiperda; adult; wavelength; phototactic rate; light stimulation; opsin
草地贪夜蛾Spodoptera frugiperda(J. E. Smith),属鳞翅目Lepidoptera夜蛾科Noctuidae,原产于美洲热带、亚热带地区[1],在美洲分布十分广泛,其种内分化为玉米型和水稻型[2]。幼虫可为害76科350多种植物[3]。该虫具有较强的迁飞能力,已在世界范围内迅速扩散[4]。草地贪夜蛾于2018年12月侵入我国,到2019年10月已扩散至26个省份[5],对我国玉米、水稻、小麦、大豆等生产造成了极大威胁[6-8]。
趋光性是指昆虫视觉器官对光线刺激产生的趋向反应,是昆虫长期适应环境的本能反应。昆虫的敏感光谱多集中于253~700 nm[9-10]。基于昆虫趋光性原理开发的诱虫灯对多数昆虫具有较好的诱杀效果;但传统诱虫灯的光谱范围宽、选择性差,在誘杀害虫的同时极易误伤天敌和传粉昆虫。因此,研发特定波长光源是实现害虫“精准诱杀”和“控害保益”的重要手段。利用滤光片、发光二极管(light emitting diode,LED)等方式产生特定波长的光,可以比较昆虫对不同波长光的趋向反应,进而探明昆虫的敏感波段[11-13]。不同昆虫感受到的光谱范围和敏感波峰具有显著差异[14],如在340~605 nm的单色光下,草地螟Loxostege sticticalis成虫对360 nm紫外光和400 nm近紫外光的趋性最高[15];在350、375、405、383 nm及333 nm的光波条件下,桃小食心虫Carposina sasakii对350 nm单色光的反应比较明显[16];棉铃虫Helicoverpa armigera对 385 nm LED灯的趋性显著高于其他波长的LED灯,且显著高于黑光灯[17];在室内条件下,梨小食心虫Grapholita molesta成虫对520 nm的绿光趋光性最强[18];405 nm的蓝紫色LED灯对印度谷螟Plodia interpunctella雄虫和雌虫的引诱效果好于435、450 nm 和470 nm的LED蓝光[19]。
此外,性别、日龄等生理因素也会影响昆虫的趋光行为,多种害虫雌雄成虫的上灯比例存在差异,宽光谱杀虫灯诱集的51种夜蛾科昆虫中,多数昆虫的雄虫上灯数量显著高于雌虫[20];而且雌雄成虫对不同波长光的趋性反应也存在差异,如黑腹果蝇Drosophila melanogaster雌成虫对360 nm紫光以及440 nm蓝光有较高的趋光率,而雄成虫则对524 nm的绿光有极强的趋光性[21]。不同日龄成虫的趋光性也存在差异,如1~5日龄的梨小食心虫成虫以3日龄趋光性最强[18];张艳红等也发现棉铃虫3日龄成虫的趋光率最高[22]。
昆虫复眼中的视蛋白是光传导过程中的重要蛋白分子[23],介导昆虫对光的感知,与其趋光反应密切相关[24-25]。根据感受的光谱范围,可将昆虫视蛋白划分为:紫外光敏感型(ultraviolet-sensitive opsin, UV-opsin),光谱吸收范围一般在325~400 nm;蓝光敏感型(blue-light-sensitive opsin, B-opsin),光谱吸收范围一般在401~500 nm;长波敏感型(long-wavelength-sensitive opsin, LW-opsin),光谱吸收范围一般>500 nm[26]。虽然大多数昆虫都有LW-opsin、B-opsin、UV-opsin型视蛋白基因,但其表达量因昆虫种类、生存环境与生理状态等不同而有所差别,不同波长、光照强度照射下视蛋白表达的变化也不相同,如Xu等研究明确了长波敏感型视蛋白基因2(LW-opsin2)在夜蛾科昆虫体内表达水平极低[27];在0~1 000 lx光照强度刺激下韭菜迟眼蕈蚊Bradysia odoriphaga 成虫的Bo-uv基因表达量与光照强度呈正比,超过1 000 lx后Bo-uv基因表达量急速下降[28];绿光照射下赤拟谷盗Tribolium castaneum 的LW-opsin、UV-opsin视蛋白表达量显著降低[29]。
已有研究结果表明,利用黑光灯可对草地贪夜蛾进行种群监测和诱集防治[30]。草地贪夜蛾成虫有4种视蛋白基因,分别为长波敏感型视蛋白1(LW-opsin1)基因,长波敏感型视蛋白2(LW-opsin2)基因,蓝光敏感型视蛋白(B-opsin)基因和紫外光敏感型视蛋白(UV-opsin)基因。虽然草地贪夜蛾和棉铃虫一样具有显著的趋光性,但草地贪夜蛾的上灯率(约50%)显著低于棉铃虫(约80%),造成这种差异的主要原因是其视蛋白基因表达水平显著低于棉铃虫[31]。为了进一步明确吸引草地贪夜蛾的特异性波长,提高诱捕效率,同时减少对天敌和传粉昆虫的伤害,减少化学农药的使用、提高农田生态系统的稳定性,本研究通过行为反应箱,测定了草地贪夜蛾雌、雄成虫对390~550 nm波长范围内光的趋光行为反应,同时测定了不同波长光刺激对草地贪夜蛾视蛋白表达量的影响。研究结果可为进一步合理应用杀虫灯对草地贪夜蛾进行监测和防治奠定基础。
1 材料与方法
1.1 供试虫源
草地贪夜蛾初始虫源于2019年1月采自云南省江城县玉米田,之后一直在中国农业科学院新乡有害生物防治野外科学观测试验站饲养。幼虫用玉米叶粉和麦胚粉制成的人工饲料饲喂,成虫羽化后放入养虫笼(边长30 cm)内,使用5%的蜂蜜水饲喂。饲养条件均为温度(26±1)℃、相对湿度(70±5)%、光周期L∥D=16 h∥8 h。
1.2 测试方法
1.2.1 草地贪夜蛾成虫对不同波长光源的趋性反应
选取3日龄健康雌雄成虫供试。试验开始前将相同性别的20头成虫放入暗室暗处理1 h,使待测个体的复眼状态保持一致。参照杨心月等的方法[32],利用行为反应箱测定成虫对390~550 nm不同波长光的趋性反应。
将趋光反应室开口处对准单色仪出光口,用黑布遮盖反应箱,使其不受其他光线影响。打开高压氙气灯,移开暗室与趋光通道之间的挡板,处理20 min 后掀开黑布,在红光灯下分别观察统计趋光通道和避光通道内的成虫数量。每个处理均重复10次,每次均随机抽取新的待测试虫。计算草地贪夜蛾雌雄成虫对不同波长的趋光率:趋光率=趋光通道虫数/试虫数×100%。
1.2.2 日龄对草地贪夜蛾趋光率的影响
在上述1.2.1测试结果的基础上,选择雌、雄成虫都较为敏感的520 nm作为测试光源,设置羽化后1、3、5 日龄的雌雄成虫共计 6个处理,每个处理20头成虫;每处理10次重复。比较不同日龄和不同性别成虫的趋光率。测试方法同1.2.1。
1.2.3 光刺激对草地贪夜蛾视蛋白基因表达量的影响
在上述1.2.1测试结果的基础上,选择雌、雄成虫都较为敏感的520 nm作为测试光源,将20头3日龄草地贪夜蛾成虫暗处理1 h后,用520 nm波长的光源照射20 min,将成虫头部迅速切下并放入含TRIzol的1.5 mL无RNA酶的离心管中,然后放入液氮中快速冷冻。以在暗室中放置80 min的成虫作为对照。试验设置3个生物学重复。所有样品均保存在-80℃冰箱中供后续使用。
用TRIzol法提取样本总RNA,用1%琼脂糖凝胶电泳及Nano Drop紫外分光光度计检测RNA浓度及纯度后采用FastKing gDNA Dispelling RT SuperMix反转录试剂盒(天根生化科技北京有限公司)合成cDNA第一链,于-20℃冰箱保存。基于NCBI中草地贪夜蛾蓝光敏感型视蛋白(B-opsin)基因、长波敏感型视蛋白1 (LW-opsin1) 基因、紫外光敏感型视蛋白(UV-opsin)基因及长波敏感型视蛋白2 (LW-opsin2) 基因(GenBank登录号分别为:MN442089~MN442092)序列,利用Primer 5.0软件设计特异性引物(表1)。引物由生工生物工程(上海)股份有限公司合成。通过PCR(反应体系:5×FastKing-RT SuperMix 10 μL, 正、反向引物各1 μL,cDNA模板1 μL,RNase-free ddH2O 7 μL)、胶回收、连接转化及菌落PCR检测,验证基因序列的正确性,然后交由生工生物工程(上海)股份有限公司进行序列测定。
基于4种视蛋白的基因序列,设计qPCR的特异性引物,选用核糖体蛋白 L27(ribosomal protein L27, RPL27)基因为内参基因[33](表1)。用SuperReal PreMix Plus (SYBP Green) 试剂盒(天根生化科技北京有限公司)进行荧光定量PCR,采用20 μL PCR反应体系:2×SuperReal PreMix Plus 10 μL,正向和反向引物各0.6 μL,cDNA模板1 μL,50×ROX Reference Dye 0.4 μL,RNase-free ddH2O 7.4 μL。反應条件为:95℃预变性15 min;95℃变性10 s,60℃退火32 s,共40个循环。3个技术重复。
1.3 数据分析
各处理原始数据在统计分析前均进行正态性检验。如果原始数据不符合正态分布,则进行相应的数据转换(计数类进行平方根转换,百分率进行反正弦平方根转换)。
草地贪夜蛾雌雄成虫对不同波长光源的趋光率、不同日龄成虫的趋光率采用单因素方差分析(One-way ANOVA)。在进行方差分析前先对各处理进行方差齐性检验(OBrien法),如果方差同质,且方差分析结果表明处理间存在显著差异,则进行Tukeys HSD多重比较;若方差不齐,则采用Dunnett T3进行多重比较。雌雄成虫间趋光率的差异显著性采用独立样本t测验进行分析。荧光定量测定的基因表达量数据,首先根据等比稀释法制作标准曲线,计算出各引物的扩增效率,采用2-ΔΔCt法计算相对表达量[34]。光刺激对草地贪夜蛾雌雄成虫视蛋白表达量的影响,采用独立样本t测验进行差异显著性分析。
2 结果与分析
2.1 草地贪夜蛾成虫对不同波长光的趋性
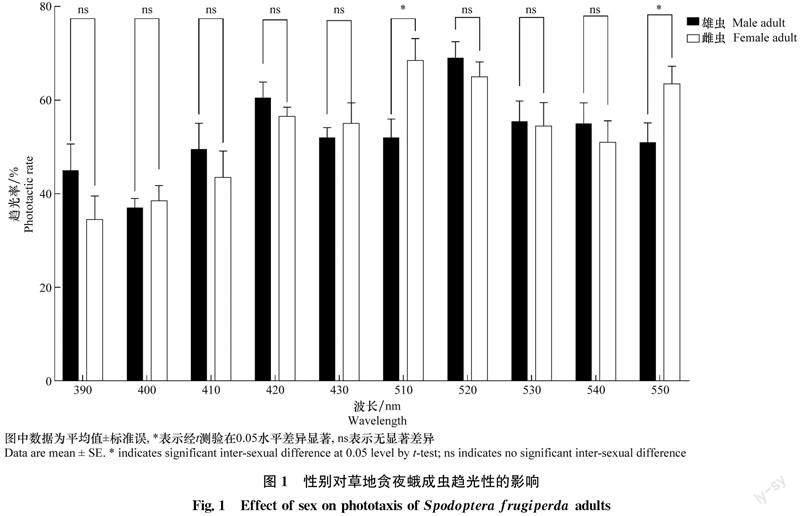
草地贪夜蛾雌蛾(F=4.433 0,df=9,90,P=0.000 1)和雄蛾(F=6.069 0,df=9,90,P=0.000 1)对不同波长光源的趋光率均存在显著差异(表2)。其中:雌蛾对绿光区510、 520 nm和550 nm波长的趋性较高,趋光率分别为(68.50±4.66)%、(65.00±3.16)%和(63.50±3.73)%;对紫外光区390 nm的趋性最低,趋光率仅为(34.50±5.02)%。雄蛾对绿光区520 nm和紫光区420 nm的趋性较高,趋光率分别为(69.0±11.01)%和(60.50±10.66)%;对紫外光区400 nm的趋性最低,趋光率为(37.00±6.32)%。
2.2 性别对草地贪夜蛾成虫趋光性的影响
草地贪夜蛾雌蛾和雄蛾对10种不同波长的趋光率比较结果如图1所示:雌蛾对510 nm(t=2.669,df=18,P=0.015)和550 nm(t=2.244,df=18,P=0.038)的趋光率显著高于雄蛾。其他波长处理下,雌雄成虫的趋光率均无显著的性别差异。
2.3 日龄对草地贪夜蛾成虫趋光性的影响
在确定草地贪夜蛾雌雄成虫都对520 nm光具有较高趋性的基础上,进一步比较了不同日龄雌雄成虫的趋光性差异,结果表明:不同日龄的雌蛾(F=2.069 8,df=2,27,P=0.145 8)和雄蛾(F=0.030 7,df=2,27,P=0.096 9)对520 nm波长光源的趋光率均存在显著差异,均表现为3日龄[雌:(65.0±3.2)%;雄:(69.0±3.5)%]和5日龄[雌:(59.1±6.0)%;雄: (61.4±4.2)%]的趋光率高,而1日龄[雌:(41.4±5.6)%;雄:(24.1±4.0)%]的趋光率最低(图2)。
2.4 光刺激对草地贪夜蛾视蛋白基因表达量的影响
首先利用PCR技术克隆得到草地贪夜蛾4种视蛋白的完整序列,通过BLAST在线对比,发现LW-opsin1、LW-opsin2、B-opsin、UV-opsin等4种视蛋白序列与GenBank上登录的草地贪夜蛾4种视蛋白序列相似度很高,分别为99.65%、98.86%、96.78%、99.74%。
进一步比较草地贪夜蛾成虫受到光刺激后视觉蛋白表达量的变化,结果(图3)表明:雄蛾UV-opsin蛋白基因在520 nm光刺激后的相对表达量显著升高(t=8.687,df=4,P=0.012),是对照组的3.61倍;但LW-opsin1(t=2.068,df=4,P=0.151)、LW-opsin2(t=0.461,df=4,P=0.118)和B-opsin (t=0.042,df=4,P=0.095)基因的相对表达量与对照组均无显著差异。雌蛾在520 nm 光刺激后,LW-opsin1(t=1.463,df=4,P=0.097)、LW-opsin2(t=0.746,df=4,P=0.112)、B-opsin(t=2.595,df=4,P=0.188)、UV-opsin(t=2.494,df=4,P=0.732)这4种视觉蛋白基因的相对表达量均与对照组无显著差异。
3 结论与讨论
前人研究表明多种昆虫具有趋光性,不同种类昆虫对不同波长呈现出不同程度的趋性,但室内与田间测定的结果常常存在差异,不同年份、不同环境下田间昆虫的趋光性也有差异,温度、湿度、降雨等气候因子都会影响昆虫的上灯率[35]。如在室内测定结果显示斜纹夜蛾 Spodoptera litura对380 nm及500 nm光源具有较高的趋光率[36];但利用不同波长LED诱虫灯对菜地昆虫的诱集效果却发现390~400 nm的LED诱虫灯对斜纹夜蛾的诱捕效果最好[37]。刘彦飞等的研究结果表明室内条件下梨小食心虫对520 nm的绿光趋性最高,趋光率为41.67%,在田间624 nm红光、590 nm黄光、520 nm绿光和465 nm蓝光等4种LED灯的诱集效果都不理想[18];但田奥等的田间试验却发现桃园梨小食心虫对紫色LED灯的趋光率最高,对黄色LED灯的趋性最弱[38]。蒋月丽等在室内评估铜绿丽金龟Anomala corpulenta 对14种单一波长的趋光反应,发现其对405、460、505、570 nm 具有明显趋向反应,对380、440、492、505 nm表现避光反应[39];但许亚岚等比较了375~748 nm LED灯对铜绿丽金龟和暗黑鳃金龟 Holotrichia parallela的田间诱捕效果,发现2015年-2019年395 nm的LED灯对两种金龟子的诱捕效果都很好;但2019年的诱捕结果却显示385 nm的LED灯诱捕到的两种金龟子数量均显著高于395 nm的LED灯[40],说明不仅室内外测定结果差异较大,而且不同年份田间害虫种群的发生数量对灯诱效果也存在显著影响。本研究通过比较草地贪夜蛾对10种不同波长的趋性,发现雌蛾对绿光区510、520 nm和550 nm的趋性较强,趋光率在63.5%~68.5%;雄蛾对紫光区420 nm和綠光区520 nm的趋性较强,趋光率分别为60.5%和69.0%。与Xu等用宽光谱365~550 nm的LED灯诱集效果(约50%)相比[27],我们筛选的几个波长,趋光率显著提高,但试验数据是在室内条件下获得的,所选波长LED灯在田间诱集况如何,需结合田间试验做进一步验证。
昆虫对不同波长的趋光性选择,除受环境因子的影响外,还受自身生理状态的影响,如食料、性别、日龄等。梨小食心虫雌蛾对绿光区520 nm的趋性(63.34%)显著高于雄蛾(36.66%),而且1~5日龄成虫中3日龄成虫的趋光性最强[18]。黑绒鳃金龟Serica orientalis雌虫的趋光率整体高于雄虫,在460、483、524 nm和605 nm单色光处理下雌虫趋光率显著高于雄虫[41];东亚小花蝽 Orius sauteri雌虫趋光率也显著高于雄虫[42]。与之不同的是相同波长处理下以烟草和白菜为食料的斜纹夜蛾趋光率无显著差异;但性别对斜纹夜蛾趋光行为影响较大,除取食烟草的斜纹夜蛾雌蛾在450 nm的趋光率高于雄蛾外,其他波长下取食烟草和白菜的雄蛾的趋光率均高于雌蛾[36]。江幸福等研究发现在一定龄期内草地螟成虫的趋光率与日龄成正比[15];而通过比较1、3、5日龄棉铃虫成虫的趋光率,发现3日龄棉铃虫对光照最为敏感[43]。这些结果表明,昆虫的趋光率受多种因素的影响,且不同昆虫间存在较大差异。本研究结果表明,510、550 nm处理下草地贪夜蛾雌蛾的趋光率显著高于雄蛾,其他波长处理下均无显著的性别间差异;3日龄和5日龄成虫的趋光性显著高于1日龄。已有研究结果表明3日龄草地贪夜蛾成虫飞行能力达到最高峰,而后随着日龄的增大飞行能力减弱[44]。我们推测草地贪夜蛾上灯率高低可能与其飞行能力有关,但具体关系还有待进一步研究。
昆虫对光线的感知主要依靠复眼,复眼内色素颗粒接收到光信号产生生物电位,最终抵达中枢神经引起视觉反应,视觉色素包含视蛋白,视蛋白在昆虫感受光刺激过程中有着不可替代的作用。视觉基因不仅在昆虫的复眼中表达,而且在棉铃虫等昆虫的脑中也高表达,说明视觉基因不仅行使视觉功能,可能还有其他非视觉功能[45]。研究发现视觉基因的表达除受到生物自身节律的影响外,也会受到昆虫发育历期和环境的影响[46-47],光照对视蛋白的表达量也具有一定的影响[48]。黏虫 Mythimna separata雌蛾LW-opsin1基因在蓝光照射1 h后表达量显著升高,UV-opsin基因在白光照射0.5 h后表达量显著升高;且雌雄成虫视蛋白基因表达量在蓝光照射下存在显著的性别差异,但白光照射下却无显著的性别差异[49];棉铃虫在绿光和紫外光照射6 h后,UV-opsin基因表达量升高[45]。光照引起昆虫视蛋白基因表达量的变化,是否与昆虫对光刺激产生反应和趋光行为反应相关,具体功能目前还没有明确。本研究发现在绿光区520 nm光刺激后,雄蛾UV-opsin蛋白基因的相对表达量显著升高,是对照组的3.61倍,但其他3种视蛋白基因的相对表达量与对照组均无显著差异,我们推测光刺激后草地贪夜蛾视觉蛋白基因表达量的变化可能是其感受光刺激、并产生相应趋性反应的关键因素。
参考文献
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738.
[2] DUMAS P, LEGEAI F, LEMAITRE C, et al. Spodoptera frugiperda (Lepidoptera: Noctuidae) host-plant variants: two host strains or two distinct species? [J]. Genetica, 2015, 143(3): 305-316.
[3] MONTEZANO D G, SOSA-GMEZ D R, SPECHT A, et al. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas [J]. African Entomology, 2018, 26(2): 286-300.
[4] EARLY R, GONZLEZ-MORENO P, MURPHY S T, et al. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm [J]. NeoBiota, 2018, 40: 25-50.
[5] 吴孔明. 中国草地贪夜蛾的防控策略[J]. 植物保护, 2020, 46(2): 1-5.
[6] 姜玉英, 刘杰, 谢茂昌, 等. 2019年我国草地贪夜蛾扩散为害规律观测[J]. 植物保护, 2019, 45(6): 10-19.
[7] 姜玉英, 刘杰, 朱晓明. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 33-35.
[8] 楊学礼, 刘永昌, 罗茗钟, 等. 云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J]. 云南农业, 2019(1): 72.
[9] HAY D A, CROSSLEY S A. The design of mazes to study Drosophila behavior [J]. Behavior Genetics, 1977, 7(5): 389-402.
[10]沈颖, 尉吉乾, 莫建初, 等. 昆虫趋光行为研究进展[J]. 河南科技学院学报(自然科学版), 2012, 40(5): 19-23.
[11]郭炳群, 李世文. 亚洲玉米螟蛾趋光行为及复眼结构节律性研究[J]. 昆虫学报, 1997(1): 58-61.
[12]OPRIAN D D. Phototaxis, chemotaxis and the missing link [J]. Trends in Biochemical Sciences, 2003, 28(4): 167-169.
[13]SHIMIZU I, KATO M. Loss of phototaxis in silkworm larvae after smelling mulberry leaves and recovery after electroconvulsive shock [J]. Nature, 1978, 272(5650): 248-249.
[14]桑文, 黄求应, 王小平,等. 中国昆虫趋光性及灯光诱虫技术的发展、成就与展望[J]. 应用昆虫学报, 2019, 56(5): 907-916.
[15]江幸福, 张总泽, 罗礼智. 草地螟成虫对不同光波和光强的趋光性[J]. 植物保护, 2010, 36(6):69-73.
[16]侯无危, 马幼飞, 高慰曾, 等. 桃小食心虫蛾的趋光性[J]. 昆虫学报, 1994(2): 165-170.
[17]PAN Hongsheng, XU Yalan, LIANG Gemei, et al. Field evaluation of light-emitting diodes to trap the cotton bollworm, Helicoverpa armigera [J/OL]. Crop Protection, 2020, 137: 105267. DOI: 10.1016/j.cropro.2020.105267.
[18]刘彦飞, 于海利, 仵均祥. 梨小食心虫对LED光的趋性及影响因素的研究[J].应用昆虫学报, 2013, 50(3): 735-741.
[19]COWAN T, GRIES G. Ultraviolet and violet light: attractive orientation cues for the Indian meal moth, Plodia interpunctella [J]. Entomologia Experimentalis et Applicata, 2009, 131(2): 148-158.
[20]WILLIAMS W E, MIDDLETON A. Vacuum wave-length measurements in the iron spectrum by means of the reflection echelon grating [J]. Proceedings of the Royal Society of London. Series A. Mathematical and Physical Sciences, 1939, 172(949): 159-172.
[21]刘倩, 高欢欢, 刘莎, 等. 光谱对斑翅果蝇和黑腹果蝇成虫趋光行为的影响[J]. 植物保护学报, 2019, 46(2): 499-500.
[22]张艳红, 刘小侠, 张青文, 等. 不同光源对棉铃虫蛾趋光率的影响[J]. 河北农业大学学报, 2009, 32(5): 69-72.
[23]段云, 吴仁海, 苗进, 等. 昆虫视蛋白的研究进展[J]. 植物保护, 2020, 46(1): 93-100.
[24]闫硕, 秦萌, 赵守歧, 等. 夜蛾复眼视觉与灯诱反应[J]. 中国植保导刊, 2017, 37(6): 30-35.
[25]LIU Yanjun, YAN Shuo, SHEN Zhongjian, et al. The expression of three opsin genes and phototactic behavior of Spodoptera exigua (Lepidoptera: Noctuidae): Evidence for visual function of opsin in phototaxis [J]. Insect Biochemistry and Molecular Biology, 2018, 96: 27-35.
[26]HENZE M J, OAKLEY T H. The dynamic evolutionary history of pancrustacean eyes and opsins [J]. Integrative and Comparative Biology, 2015, 55(5): 830-842.
[27]XU Pengjun, FEUDA R, LU Bin, et al. Functional opsin retrogene in nocturnal moth [J/OL]. Mobile DNA, 2016, 7(1): 18. DOI: 10.1186/s13100-016-0074-8.
[28]安立娜, 范凡, 杨小凡, 等. 韭菜迟眼蕈蚊紫外敏感视蛋白基因的克隆及光强度对其表达量影响[J]. 植物保护学报, 2019, 46(5): 971-979.
[29]劉芬, 毛莹, 李琪诗, 等. 赤拟谷盗视蛋白基因克隆及其对不同光源的响应研究[J]. 华中昆虫研究, 2018, 14: 291-299.
[30]MITCHELL E R. Monitoring adult populations of the fall armyworm [J]. Florida Entomologist, 1979, 66(2): 91-98.
[31]LIU Yingjie, ZHANG Dandan, YANG Liyu, et al. Analysis of phototactic responses in Spodoptera frugiperda using Helicoverpa armigera as control [J]. Journal of Integrative Agriculture, 2021, 20(3): 821-828.
[32]杨心月, 范凡, 陈洁, 等. 光谱对二点委夜蛾成虫趋光行为的影响[J]. 植物保护学报, 2015, 42(6): 1009-1013.
[33]KARAMIPOUR N, FATHIPOUR Y, TALEBI A A, et al. Small interfering RNA pathway contributes to antiviral immunity in Spodoptera frugiperda (Sf9) cells following Autographa californica multiple nucleopolyhedrovirus infection [J]. Insect Biochemistry and Molecular Biology, 2018, 101: 24-31.
[34]LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method [J]. Methods, 2001, 25(4): 402-408.
[35]陈坤, 文礼章, 龚碧涯, 等. 重要气象因子对农田灯光诱虫效率的影响[J]. 中国农学通报, 2013, 29(4): 44-50.
[36]田太安, 商胜华, 王小彦, 等. 不同食料斜纹夜蛾成虫的趋光性[J]. 贵州农业科学, 2019, 47(8): 71-74.
[37]吴霜, 张谊模, 郭萧, 等. 不同波长LED诱虫灯对菜地昆虫的诱集效果[J]. 应用昆虫学报, 2021, 58(1): 172-180.
[38]田奥, 顾鹏, 赵利鼎, 等. 桃园梨小食心虫的趋光性初探[J]. 农业技术与装备, 2012(10): 74-76.
[39]蒋月丽, 武予清, 李彤, 等. 铜绿丽金龟对不同光谱的行为反应[J]. 昆虫学报, 2015, 58(10): 1146-1150.
[40]许亚岚, 潘洪生, 梁革梅, 等. 不同波长LED灯对铜绿丽金龟和暗黑鳃金龟的田间诱捕效果[J]. 新疆农业科学, 2020, 58(11): 2028-2033.
[41]吕飞, 海小霞, 范凡, 等. 黑绒鳃金龟甲成虫对不同单色光和光强的趋光行为[J]. 植物保护学报, 2016, 43(4): 656-661.
[42]冯娜, 范凡, 陶晡, 等. 光谱对东亚小花蝽趋光行为的影响[J]. 生态学报, 2015, 35(14): 4810-4815.
[43]魏国树, 张青文, 周明牂, 等. 不同光波及光强度下棉铃虫(Helicoverpa armigera)成虫的行为反应[J]. 生物物理学报, 2000(1): 89-95.
[44]葛世帅, 何莉梅, 和伟, 等. 草地贪夜蛾的飞行能力测定[J]. 植物保护, 2019, 45(4): 28-33.
[45]YAN Shuo, ZHU JiaLin, ZHU Weilong, et al. The expression of three opsin genes from the compound eye of Helicoverpa armigera (Lepidoptera: Noctuidae) is regulated by a circadian clock, light conditions and nutritional status [J/OL]. PLoS ONE, 2014, 9(10): e111683. DOI: 10.1371/journal.pone.0111683.
[46]LICHTENSTEIN L, GRBEL K, SPAETHE J. Opsin expression patterns coincide with photoreceptor development during pupal development in the honey bee, Apismellifera [J/OL]. BMC Developmental Biology, 2018, 18(1): 1. DOI: 10.1186/s12861-0162-8.
[47]FRENTIU F D, YUAN F, SAVAGE W K, et al. Opsin clines in butterflies suggest novel roles for insect photopigments [J]. Molecular Biology and Evolution, 2015, 32(2): 368-379.
[48]ALLADA R, CHUNG B Y. Circadian organization of behavior and physiology in Drosophila [J]. Annual Review of Physiology, 2010, 72(1): 605-624.
[49]薛彧媛, 彭文菊, 劉芬, 等. 不同光照对黏虫视蛋白基因表达的影响[J]. 华中昆虫研究, 2018, 14(00): 282-287.
(责任编辑:杨明丽)
猜你喜欢
特产研究(2022年6期)2023-01-17
环境昆虫学报(2020年3期)2020-07-07
农家科技中旬版(2019年8期)2019-09-19
南方农业·下旬(2019年4期)2019-08-26
山东农业科学(2019年6期)2019-08-16
现代农业科技(2019年12期)2019-08-07
现代农村科技(2018年8期)2018-08-23
实用口腔医学杂志(2017年6期)2017-09-19
金色少年(奇趣科普)(2017年6期)2017-07-25
中国照明(2016年4期)2016-05-17
