
外源海藻糖对灌浆期高温胁迫下小麦旗叶生理特性和产量的影响
2023-06-09 09:22李宇星马亮亮秦博雅张文静马尚宇黄正来樊永惠
作物学报 2023年8期
李宇星 马亮亮 张 月 秦博雅 张文静 马尚宇 黄正来 樊永惠
外源海藻糖对灌浆期高温胁迫下小麦旗叶生理特性和产量的影响
李宇星 马亮亮 张 月 秦博雅 张文静 马尚宇 黄正来*樊永惠*
安徽农业大学农学院 / 农业农村部黄淮南部小麦生物学与遗传育种重点实验室, 安徽合肥 230036
为探究外源海藻糖(TRE)对高温胁迫下灌浆期不同耐性小麦品种旗叶生理特性和产量的影响作用, 于2020—2022年在安徽农业大学高新技术农业园进行试验, 选用前期筛选得的耐热性差异显著的敏感型小麦品种泛麦5号(Fanmai 5, FM5)和耐热型小麦品种淮麦33 (Huaimai 33, HM33)作为试验材料, 设置叶面喷施清水+不高温(CK1)、清水+灌浆期高温胁迫(CK2)、10 mmol L–1海藻糖+灌浆期高温胁迫(T10H)、15 mmol L–1海藻糖+灌浆期高温胁迫(T15H)和20 mmol L–1海藻糖+灌浆期高温胁迫(T20H)共5个处理。结果表明, 在高温胁迫条件下, 绿叶面积、叶绿素相对含量(SPAD)和干物质积累量均显著下降, 与非高温逆境相比, 高温胁迫下小麦产量显著下降, 穗数和穗粒数无显著变化, 千粒重是减产的主导因素。与喷清水相比, 喷施海藻糖后产量较高温胁迫处理有所提升, 各器官干物质积累量提高, 丙二醛(MDA)含量降低, 超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性提高, 变化幅度在品种间存在差异, 耐热品种的SPAD值降幅与丙二醛的增幅较小, 但CAT活性的增幅较大, 因而减产幅度较小。进一步的分析表明, 净光合速率(n)、气孔导度(s)、蒸腾速率(r)、SPAD、SOD及POD和千粒重呈极显著正相关关系, MDA和产量呈极显著负相关关系。这些结果表明高温胁迫通过降低n、s和r抑制小麦的光合作用, 减少光合产物的生成, 造成最终的减产。喷施TRE,n、s、r、SPAD和CAT是降低高温胁迫伤害的主要指标, 敏感品种缓解效果明显。此外, 开花期喷施浓度15 mmol L–1海藻糖效果最好。该研究结果可为海藻糖对灌浆期高温胁迫下小麦光合响应差异机制和小麦抗氧化代谢提供理论依据。
小麦; 高温胁迫; 海藻糖; 旗叶; 产量
小麦作为我国主产作物, 在人们的日常生活中占据重要地位[1]。提高小麦产量, 保障我国口粮绝对安全的根基更为牢固[2]。温度是作物生长发育的主要因素之一, 适宜的环境温度才能保证作物的稳产高产[3]。随着全球气候变暖的不断加剧, 灌浆期遭受高温胁迫的风险增加[4]。研究显示, 小麦灌浆期的适宜温度是20~24℃, 每比适宜温度高1℃, 小麦就会减产5%~10%, 情况严重会导致绝收[5]。灌浆期高温胁迫, 导致小麦叶片光合器官受到破坏, 抗氧化酶活性降低, 过氧化物积累, 膜脂过氧化, 光合能力下降, 营养器官提前衰老, 导致光合同化物供给减少, 进而导致籽粒灌浆过程的提前结束, 最终使达到最大籽粒干重的时间缩短, 产量降低[6-7]。黄淮海地区是我国重要的粮食主产区和冬小麦种植区, 其冬小麦产量对气候变化的响应对中国的粮食安全具有重要意义[8]。黄淮海地区在小麦灌浆期经常出现高温天气, 对小麦产量造成严重影响[9]。海藻糖(TRE)是非还原性糖, 具有较强的亲水保水能力、对高温不敏感, 稳定性很强, 在植物体内痕量级存在[10-11]。但在逆境条件下, TRE含量会显著上升。研究显示, TRE可以保护细胞生物膜系统、蛋白质等免受高温伤害[12]。外源TRE的添加有助于缓解高温胁迫, 维持植物较强的呼吸作用, 并且降低营养器官中淀粉含量的积累, 缓解光合作用负反馈机制, 并且有助于淀粉在源-库之间的转运和分配, 从而增强光合作用, 使光合同化物积累[13]。同时可以作为信号分子, 调节抗氧化酶相关基因的表达和酶的活性, 缓解植物离子胁迫, 维持细胞稳定, 增强植物的耐性[14]。然而目前有关外源TRE对高温胁迫下作物影响的研究大多集中在园艺作物[15-16], 此外, 有关小麦在高温胁迫下的缓解措施多集中于水杨酸(SA)[17]、磷酸二氢钾(KH2PO4)[18]和一氧化氮(NO)[19], 而外源TRE对灌浆期高温胁迫下小麦旗叶生理特性和产量的影响结果尚未明确。
为此, 本研究以小麦品种泛麦5号和淮麦33为试验材料, 探讨TRE对高温胁迫下冬小麦光合能力和抗氧化生理特性的影响, 阐明其生理机制, 以期为冬小麦抗高温生产提供理论依据和技术指导。
1 材料与方法
1.1 试验地点
2020—2022年, 本试验在安徽农业大学高新技术农业园(31.85°N, 117.27°E)进行。前茬作物为玉米, 土壤类型为棕壤土, 播种前试验田土壤养分含量如表1所示。

表1 播种前试验田0~20 cm土壤养分含量
1.2 试验设计
采用裂区设计, 其中品种为主区, 选用敏感型冬小麦品种泛麦5号(Fanmai 5, FM5)和耐热型冬小麦品种淮麦33 (Huaimai 33, HM33)作为试验材料, 两个品种的生育期均在228 d左右。喷施浓度处理为副区, 温度处理为裂区, 共设置叶面喷等量清水+不高温(CK1)、喷等量清水+高温胁迫(CK2)、喷10 mmol L–1海藻糖+灌浆期高温胁迫(T10H)、喷15 mmol L–1海藻糖+灌浆期高温胁迫(T15H)和喷20 mmol L–1海藻糖+灌浆期高温胁迫(T20H)共5个处理。于开花期使用清水和海藻糖(trehalose, TRE, 纯度为99%)对小麦进行叶面喷雾3 d, 喷施用量为100 mL m–2, 喷施至叶片正反两面均湿润为止(有一层小水珠欲落), 每天1次(傍晚), 对照组(control check 1, CK1)喷施等量的溶剂(清水), 每100 mL溶液含有0.02 mL吐温-20 (Tween-20), 能提高外源物的吸附能力[20-23]。小区面积为9 m2(3 m × 3 m), 每处理重复3次, 随机排列, 人工开沟播种。基本苗为375万株 hm–2。全生育期施纯氮240 kg hm–2, 氮肥分3次施入, 基追比(基肥∶拔节肥∶孕穗肥)为5∶3∶2。小麦全生育期施用肥料(P2O5100 kg hm–2、K2O 150 kg hm–2), 磷、钾肥全部作为底肥一次性施入。其余管理措施同大田高产栽培。
试验通过被动式增温棚模拟高温环境, 采用钢管搭建大棚框架(6.0 m×6.0 m×2.5 m), 表面覆盖透明聚乙烯膜(厚度为0.07 mm), 其透光率达到90%以上。2021年各品种的开花时间为4月13日, 2022年各品种的开花时间为4月11日。两年增温时间均是从小区小麦50%开花算起, 第15天开始增温, 连续增温5 d, 每天增温时间段为09:00–17:00 (图1-A, B)。以Elitech (RC-4hc)智能温湿度记录仪(徐州江苏精创电器股份有限公司)每10 min 1次连续记录小麦冠层空气温湿度[17]。两年增温时段未遇到降雨天气, 处理结束后将大棚移走, 让小麦继续在自然环境下生长。2020— 2021年灌浆期高温胁迫平均增温6.18℃, 2021—2022年灌浆期高温胁迫平均增温5.13℃ (图1-C, D)。

图1 2021–2022试验期间试验地棚内/棚外平均温度(A, B)和棚内/棚外温度变化动态图(C, D)
1.3 测定项目与方法
1.3.1 收获、考种和测产 成熟期调查一米双行穗数, 收获后进行室内考种, 统计穗粒数和千粒重, 每个小区收获1 m2, 风干后脱粒, 称重并合成13%含水量进行产量测定。
干物质相关指标取样方法: 在成熟期每个处理取30株单茎, 3次重复, 分为穗、旗叶、倒二叶、倒三叶、余叶、倒一节和余节, 80℃烘干至恒重, 并计算干物质积累量。
各器官干物质积累量变化率(%)=(A处理器官干物质积累量–B处理器官干物质积累量)/B处理器官干物质积累量×100; 式中, A代表CK2和TRE处理, B代表CK1处理。
1.3.2 光合参数 于花后21 d、28 d采用便携式光合速率测定仪LI-6800 (美国)测定小麦叶片净光合速率(n)、气孔导度(s)和蒸腾速率(r)。配置的人工光源将光合有效辐射(PAR)设定在1200 μmol m–2s–1,叶室温度控制在25℃, 二氧化碳(CO2)浓度控制在400 μmol mol–1左右, 测试时间选在09:30—11:00。
于花后21 d、28 d采用便携式SPAD-502型叶绿素仪(Konica Minolta, 日本)测定小麦旗叶SPAD值, 重复10次。每处理测量10株小麦旗叶绿叶的长和宽, 按系数法计算旗叶绿叶面积(绿叶面积=绿叶长×绿叶宽×0.85)[24]。
1.3.3 抗氧化特性 开花期选择长势一致的小麦植株进行标花。分别于花后21 d、28 d的09:00— 11:00, 取生长一致的旗叶, 液氮速冻后放入–80℃冰柜保存。液氮研磨成粉末, 采用硫代巴比妥酸法测定旗叶丙二醛(MDA)含量[25], 采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性[26]; 采用愈创木酚法测定旗叶过氧化物酶(POD)活性[27]; 紫外吸收法测定旗叶过氧化氢酶(CAT)活性[28]。
1.4 数据分析
采用Microsoft Excel 2016软件进行数据处理, DPS对试验数据进行显著性分析, 用Duncan’s分析方法进行多重比较, 并采用Origin 8.5软件作图。
2 结果与分析
2.1 产量
2.1.1 产量及其构成因素 由图2可知, 高温胁迫(CK2)条件下, 耐热品种淮麦33 (HM33)产量较对照(CK1)处理2年平均下降9.06%, 敏感品种泛麦5号(FM5)下降13.05%。不同浓度TRE喷施处理下, 产量均高于高温胁迫处理, 产量的增加主要源于千粒重的增加。较高温胁迫未喷施TRE处理, HM33喷施浓度10、15和20 mmol L–1TRE处理两年平均分别增产2.28%、5.47%和2.70%, FM5平均分别增产4.43%、8.09%和4.54%; HM33千粒重平均分别增加2.28%、5.46%和2.70%, FM5平均分别增加4.43%、8.00%和4.53%, 穗粒数和穗数无显著差异。方差分析结果表明, 高温胁迫和TRE处理对小麦产量及千粒重的影响大于年份和品种的影响(表2)。
2.1.2 生物量 由表3和表4可知, 2021年耐热型品种HM33干物质积累量整体高于敏感型品种FM5, 灌浆期高温胁迫处理下, 2个品种干物质积累量均显著降低, FM5总干物质量和各器官干物质量降低幅度大于HM33, 其中穗和余叶降低幅度在各器官中最为明显, FM5穗和余叶相较HM33降低幅度高56.25%和47.87%, 2022年表现一致(表5和表6)。进一步研究分析, 2022年FM5高温胁迫处理下上三叶干物质积累量降低幅度大于HM33, 其中倒二叶降低幅度最为明显。喷施TRE处理条件下干物质积累量降低均有所缓解, 2021年HM33全株总叶片的干物质积累量在T10H、T15H和T20H条件下相较CK2处理分别提高8.49%、14.31%和4.17%, FM5分别提高10.07%、28.33%和13.28%; 2022年HM33上三叶的干物质量较CK2处理分别提高2.01%、11.35%和1.93%, FM5分别提高7.43%、20.34%和2.44%。进一步研究分析, 喷施海藻糖处理条件下, 上三叶中旗叶的提升效果最为显著, HM33相较CK2处理2年平均分别提高3.39%、4.26%和2.14%, FM5分别提高11.94%、20.11%和8.22%。2个品种茎的干物质积累量变化差异显著, FM5茎在T10H、T15H和T20H条件下提高幅度相较HM332年平均分别高10.08%、26.37%和17.30%, 进一步分析, 2个品种剩余节提升幅度差异显著, 其中不同浓度处理之间提升关系表现为T15H>T10H>T20H, FM5在T15H条件下相较HM33提升幅度高38.06%, 倒一节提升幅度品种间差异不显著。

图2 外源海藻糖对灌浆期高温胁迫下小麦产量及其构成因素的影响
CK1: 叶面喷等量清水+不高温; CK2: 喷等量清水+高温; T10H: 喷10 mmol L–1海藻糖+灌浆期高温胁迫; T15H: 喷15 mmol L–1海藻糖+灌浆期高温胁迫; T20H: 喷20 mmol L–1海藻糖+灌浆期高温胁迫。不同小写字母代表同一时期不同处理间达到显著水平(< 0.05)。FM5:泛麦5号; HM33: 淮麦33。
CK1: spray equal amount of clean water on the leaf surface + not high temperature; CK2: spray equal amount of clean water + high temperature; T10H: spray 10 mmol L–1trehalose + high temperature stress during filling; T15H: spray 15 mmol L–1trehalose + high temperature stress during filling; T20H: spray 20 mmol L–1trehalose + high temperature stress during filling. Different lowercase letters represent significant difference among different treatments in the same period (< 0.05). FM5: Fanmai 5; HM33: Huaimai 33.

表2 外源海藻糖对灌浆期高温胁迫下小麦产量和产量构成因素的方差分析
*:< 0.05;**:< 0.01; ns: 无显著差异。*:< 0.05;**:< 0.01; ns: no significance.

表3 2021年外源海藻糖对灌浆期高温胁迫下小麦成熟期干物质积累量的影响

(续表3)
CK1: 叶面喷等量清水+不高温; CK2: 喷等量清水+高温; T10H: 喷10 mmol L–1海藻糖+灌浆期高温胁迫; T15H: 喷15 mmol L–1海藻糖+灌浆期高温胁迫; T20H: 喷20 mmol L–1海藻糖+灌浆期高温胁迫。同一列内不同小写字母表示在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
CK1: spray equal amount of clean water on the leaf surface + not high temperature; CK2: spray equal amount of clean water + high temperature; T10H: spray 10 mmol L–1trehalose + high temperature stress at filling stage; T15H: spray 15 mmol L–1trehalose + high temperature stress at filling stage; T20H: spray 20 mmol L–1trehalose + high temperature stress during filling. Different lowercase letters in the same column indicate significant at the 0.05 probability level. FM5: Fanmai 5; HM33: Huaimai 33.

表4 2022年外源海藻糖对灌浆期高温胁迫下小麦成熟期干物质积累量的影响
处理同表3。同一列内不同小写字母表示在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Table 3. Different lowercase letters in the same column indicate significant difference at the 0.05 probability level. FM5: Fanmai 5; HM33: Huaimai 33.

表5 2021年外源海藻糖对灌浆期高温胁迫下小麦各器官干物质积累量变化率的影响
处理同表3。同一列内不同小写字母表示在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Table 3. Different lowercase letters in the same column indicate significant difference at the 0.05 probability level. FM5: Fanmai 5; HM33: Huaimai 33.

2.2 光合参数
2.2.1 旗叶绿叶面积、叶绿素相对含量(SPAD)
灌浆期高温胁迫处理下, 2个品种旗叶绿叶面积在花后21 d和28 d平均分别降低20.42%和51.32%, 降低幅度28 d大于21 d (图3-A, B)。品种间比较, 花后21 d, 耐热型品种HM33和敏感型品种FM5高温胁迫处理相较CK1处理分别降低18.84%和21.99%; 花后28 d, 分别降低48.90%和53.75%, FM5降幅大于HM33。花后21 d, HM33在TRE浓度处理下较CK2处理显著缓解了旗叶绿叶面积的减少, T10H、T15H和T20H分别提高8.03%、9.22%和3.37%, 花后28 d, T15H显著大于T10H和T20H。FM5在花后21 d相较CK2处理分别提高5.58%、9.56%和0.27%, 缓解关系表现为T15H>T10H>T20H, 花后28 d各浓度间差异不显著。CK2处理下2个品种SPAD值均显著降低, TRE浓度处理下HM33较CK2处理SPAD值显著提高, T10H、T15H和T20H在花后21 d分别提高8.75%、12.21%和9.46%, 花后28 d提升幅度小于花后21 d, 分别提高4.01%、8.58%和3.46%, FM5与之一致, 但提升幅度均小于HM33 (图3-C, D)。同时, 从方差分析结果可以看出, 花后天数与品种和花后天数与处理交互对绿叶面积和SPAD值影响显著(表7)。

图3 外源海藻糖对灌浆期高温胁迫下小麦旗叶绿叶面积(A, B)和叶绿素相对含量(C, D)的影响
处理同图2。不同小写字母代表同一时期不同处理间在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Figure 2. Different lowercase letters represent significant levels among different treatments in the same period (< 0.05). FM5: Fanmai 5; HM33: Huaimai 33.
2.2.2 净光合速率(n)、气孔导度(s)和蒸腾速率(r) 灌浆期高温胁迫处理下, 花后21 d耐热型品种HM33和敏感型品种FM5的n分别较CK1处理降低20.19%和27.40%, 花后28 d分别降低55.27%和61.54%, 2个品种花后21 d降低幅度均大于28 d (图4-A, B)。品种间比较, FM5降低幅度大于HM33。喷施TRE处理较未喷施处理显著提高n的下降幅度, HM33在花后21 d, T10H、T15H和T20H分别较CK2处理提高2.05%、12.97%和1.44%, FM5提升幅度显著大于HM33, 分别提升13.83%、19.08%和11.69%。花后28 d, T15H显著大于T10H和T20H, T10H和T20H差异不显著。缓解关系表现为T15H>T10H>T20H,s和r与n表现一致(图4-C, D, E, F)。同时, 从方差分析结果可以看出, 花后天数和处理对n、s和r的影响大于品种的影响(表7)。
2.3 抗氧化特性
2.3.1 丙二醛(MDA)含量 由图5可知, 随着生育进程的推进, 2个品种旗叶花后28 d的MDA含量各处理均显著高于21 d, 灌浆期高温胁迫处理下, CK2处理均显著高于CK1, 耐热型品种HM33花后21 d和28 d较CK1处理分别高22.43%和26.31%, 敏感型品种FM5分别高23.65%和28.02%, HM33提高程度小于FM5。喷施TRE处理缓解了MDA的提高, HM33花后21 d, T10H、T15H和T20H分别较H处理降低6.19%、10.75%和4.24%, 花后28 d分别降低6.01%、10.65%和2.41%, 缓解关系表现为T15H> T10H>T20H, FM5花后21 d较HM33缓解效果分别高1.16%、6.43%和1.37%, 花后28 d分别高2.13%、6.19%和2.01%。

图4 外源海藻糖对灌浆期高温胁迫下小麦旗叶净光合速率(A, B)、气孔导度(C, D)和蒸腾速率(E, F)的影响
处理同图2。不同小写字母代表同一时期不同处理间在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Fig. 2. Different lowercase letters represent significant difference among the different treatments in the same period (0.05). FM5: Fanmai 5; HM33: Huaimai 33.

表7 外源海藻糖对灌浆期高温胁迫下小麦光合参数的方差分析
*:< 0.05;**:< 0.01; ns: 无显著差异。*:< 0.05;**:< 0.01; ns: no significance.

图5 外源海藻糖对灌浆期高温胁迫下小麦旗叶丙二醛含量的影响
处理同图2。不同小写字母代表同一时期不同处理间在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Fig. 2. Different lowercase letters represent significant difference among the different treatments in the same period (0.05). FM5: Fanmai 5; HM33: Huaimai 33.
2.3.2 超氧化物岐化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性 由图6可知, 旗叶花后28 d抗氧化酶活性较21 d均显著降低。灌浆期高温胁迫处理下, HM33的SOD活性在花后21 d和28 d显著降低16.76%和19.56%, FM5分别降低18.49%和21.69%。喷施TRE处理缓解了SOD活性的降低, HM33在花后21 d, T10H、T15H和T20H分别较CK2处理提高8.43%、12.41%和7.02%, 花后28 d分别提高6.89%、11.93%和3.87%。品种间比较, 花后21 d, FM5相较HM33各处理分别提高31.73%、21.97%和46.73%; 花后28 d, 分别提高19.39%、6.11%和35.71%。POD和CAT活性与SOD活性规律类似, 花后21 d, 2个品种T15H处理显著高于其他两个喷施TRE处理, HM33各处理POD活性相较花后28 d平均高1.37%, FM5高2.12%。花后21 d和28 d, FM5各处理CAT活性提升幅度均高于HM33, 相比平均分别高3.37%、13.91%和14.10%。喷施TRE处理, 各浓度抗氧化酶活性缓解关系表现为T15H>T10H> T20H。
2.4 产量、光合参数和抗氧化特性指标的相关性分析
由表8可知, 产量与花后旗叶相关性状呈极显著正相关, 与MDA呈极显著负相关, 千粒重的表现与产量类似, 与n、s、r、SPAD、SOD和POD呈极显著正相关, 与绿叶面积和CAT呈显著正相关; 穗数和穗粒数与旗叶相关性状无显著相关性。

图6 外源海藻糖对灌浆期高温胁迫下小麦旗叶SOD (A, B)和POD (C, D)和CAT (E, F)活性的影响
处理同图2。不同小写字母代表同一时期不同处理间在0.05概率水平差异显著。FM5: 泛麦5号; HM33: 淮麦33。
Treatments are the same as those given in Fig. 2. Different lowercase letters represent significant difference among the different treatments in the same period (0.05). FM5: Fanmai 5; HM33: Huaimai 33.
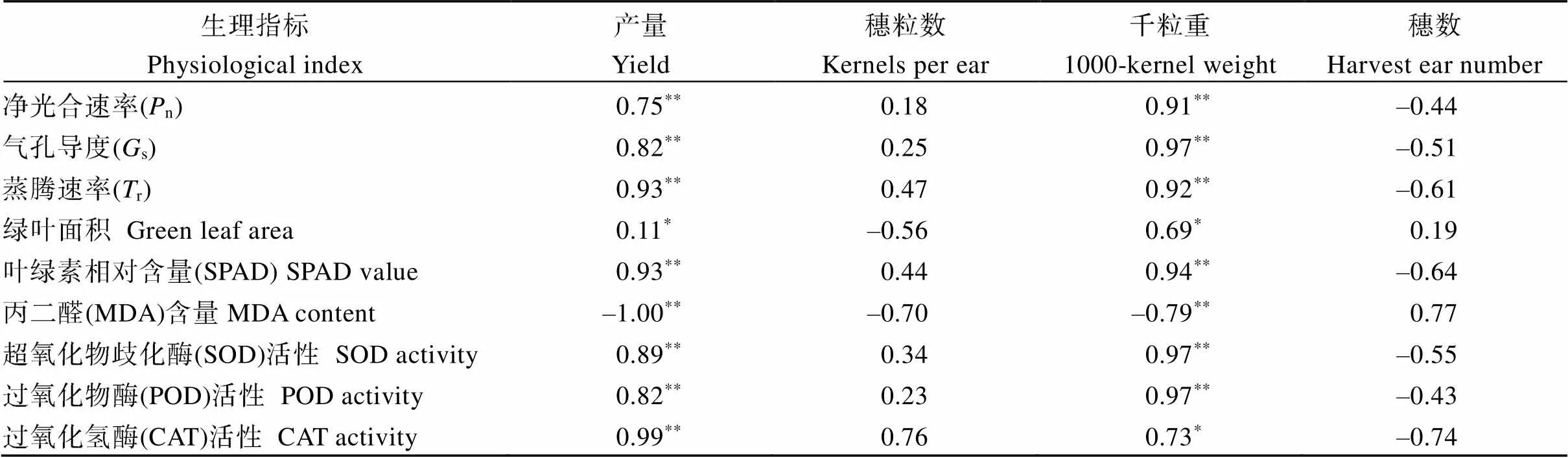
表8 产量及其构成因素和光合参数与抗氧化特性因子的相关性分析
*:< 0.05;**:< 0.01.
3 讨论
3.1 外源海藻糖对灌浆期高温胁迫下小麦产量和生物量的影响
随着全球气候变暖的加剧, 高温逆境频发, 对我国小麦产量安全造成严重的威胁, 灌浆期高温胁迫导致小麦产量减少[29]。小麦产量构成的主要因素是单位面积穗数、穗粒数和千粒重。有研究表明, 高温胁迫通过影响小麦的穗粒数, 从而影响小麦的产量[30]。也有研究认为, 高温胁迫通过影响小麦的粒重来影响小麦的产量[31]。本试验结果显示, 小麦千粒重降低是造成产量减少的主要原因, 这与岳鹏莉等[29]的研究结果相一致, 这是因为籽粒灌浆过程对温度变化非常敏感, 高温胁迫会降低小麦籽粒胚乳细胞数目, 造成胚乳细胞增殖速度变缓, 库容量降低[32]。本试验中2个品种在高温胁迫下均下降, 但降低幅度稍有差别, 这可能与小麦品种的高温耐性有关。耐热型品种HM33高温胁迫条件下, 产量降低幅度小于敏感型品种FM5, 这可能由于耐热型品种遭到高温后恢复能力较强, 受高温影响小。诸多研究结果已经验证, 利用外源物质诱导作物产生获得抗热性可能是作物耐热对策的一个重要突破口[33]。有研究证实, 外源TRE处理可以显著提高高温逆境条件下的产量[34]。外源TRE的这种抗逆效应在本研究中也得到证实, 外源TRE显著提高了高温胁迫下小麦的产量, 开花期喷施浓度15 mmol L–1TRE对灌浆期高温胁迫下小麦产量降低的缓解效果最佳。“源”与“库”是产量形成的2个方面, 产量高低既依赖于“库”的容量, 又取决于“源”的生产和光合产物的运输能力。高温胁迫抑制作为“源”器官叶片的光合作用, 灌浆物质“源”不足, 导致小麦干物质积累量下降[35]。在本研究中, 高温胁迫后, 小麦各器官质量的比例显著降低, 这表明, 灌浆期高温胁迫通过影响各器官的干物质积累量并最终影响小麦产量, 这可能是因为高温胁迫导致小麦光合器官受损, 过氧化物积累, 使叶片和茎鞘等非生殖器官中的营养物质无法由光合作用转化。本研究发现, 敏感型品种FM5相较耐热型品种HM33, 穗和上三叶中的倒二叶降幅最为明显, 喷施TRE处理, 旗叶缓解幅度最大, 因为灌浆期旗叶和倒二叶叶绿素含量影响较大, 对产量性状有影响[36]。本试验结果表明, 开花期喷施浓度15 mmol L–1TRE后增加了生殖器官干物质积累量, 这将有利于缓解高温胁迫下粒重的降低, 缓解高温胁迫下小麦减产幅度。
3.2 外源海藻糖对灌浆期高温胁迫下小麦光合参数的影响
王磊[37]认为光合效率的高低是衡量作物产量高低的重要指标。高温胁迫抑制了植株生长, 引起次生代谢产物积累, 降解叶绿素和破坏光合系统, 导致功能叶提前衰老, 最后引起光合速率下降[38]。庞椿朋等[16]指出TRE对高温下番茄幼苗光合系统起到保护作用。在本研究中, 外源TRE缓解了高温胁迫下小麦旗叶绿叶面积的降低, 缓解了旗叶捕获光能有效面积的损失, 相对延缓了旗叶的衰老。植物进行光合作用的主要色素是叶绿素和类胡萝卜素, 在植物的光合作用中叶绿素对光能的吸收、传递和转化起着极为重要的作用[39]。本研究表明, 高温胁迫下植株的叶绿素相对含量降低, 外源TRE喷施处理对高温胁迫下小麦旗叶的SPAD的降低有所缓解。TRE提高旗叶SPAD的原因可能与其参与保护叶绿体膜结构的完整性、调节叶绿体内抗氧化系统平衡有关。本研究中, TRE处理提高了n、s和r, 有效地吸收和保持水分, 并阻止水分过度蒸发, 保证叶片在高温胁迫下进行光合作用的水分供给, 防止脱水对植物造成伤害。TRE缓解气孔减少的趋势, 导致二氧化碳固定量提高, 有机物积累量提高, 产量下降幅度减缓。有研究指出在高温胁迫下, 气孔关闭, 植株叶绿体结构破坏, 叶绿素含量减少, 而TRE使植物在热胁迫下产生大量热激蛋白(HSPs), 提高抗氧化酶活性, 减轻高温诱导的氧化胁迫对植物的损伤, 确保细胞膜的稳定性, 增强叶绿体对光能的吸收和利用能力, 进而缓解高温胁迫导致的叶绿体破坏, 提高了植株的抗热能力[40]。开花期TRE喷施后, 旗叶SPAD和绿叶面积提高, 潜在的光合能力和光能利用效率提高, 保证光合作用的正常进行, 促进光合产物的形成和转运, 提高干物质的积累, 进而缓解高温胁迫下产量下降的趋势, 喷施浓度15 mmol L–1TRE效果最好。同时, 本研究发现, TRE喷施后, 敏感型品种FM5光合能力缓解效果好于耐热型品种HM33, 这可能是因为敏感型品种抵御逆境能力较差, 外源TRE的加入更好地保护光合器官生理功能。
3.3 外源海藻糖对灌浆期高温胁迫下小麦抗氧化特性的影响
植物在遭受高温胁迫时, 抑制光合作用环式电子传递链, 促进假环式电子传递链的运行, 产生大量自由基和活性氧, 从而引起膜脂质过氧化, 导致植物代谢紊乱[41]。研究表明, 高温胁迫下植物体内SOD、POD和CAT活性均会随着温度的升高而下降, 随着高温时间的延长植物体内抗氧化酶活性下降, 从而使得活性氧含量上升破坏膜系统产生MDA, 造成MDA含量呈显著上升趋势[42]。高温胁迫还会造成膜透性增加, 加剧衰老速度[43]。有研究发现TRE可以增强膜的流动性, 膜的流动性增强有利于植株对营养元素的吸收和对有害离子的排斥, 使细胞与外界的信息交流和营养运输加强, 降低对细胞自身伤害[38]。樊永惠等[17]研究发现, 开花期预喷施水杨酸(SA)缓解高温胁迫效果最好。旗叶抗氧化酶活性有所提高, 增强了旗叶清除活性氧的能力, 抗逆酶苯丙氨酸解氨酶和多酚氧化酶活性提高, 外源SA增强了小麦对高温环境的抵抗能力, 从而延缓了旗叶的衰老速率。唐秀巧等[18]研究表明, 花后3 d和11 d叶面喷施磷酸二氢钾, 通过增强高温胁迫下根系抗氧化酶活性、降低MDA含量的积累维持根系活力和叶片较高的光合能力, 从而提高成熟期植株干物质积累, 进而提高粒重, 增加小麦产量。也有研究发现, 经过0.1 mmol L–1的一氧化氮(NO)供体硝普钠(SNP)处理后, 小麦的抗氧化酶SOD、CAT活性以及脯氨酸含量得到提高, 叶片膜质过氧化产物MDA含量和相对电导率降低, 促使最大光化学效率(v/m)处于较高水平, 并维持较低的原初荧光(o), 进而减轻了活性氧(ROS)的伤害, 进而对小麦的光合机构起到保护作用, 改善小麦对高温强光胁迫的适应性[19]。本试验结果表明, 开花期喷施浓度15 mmol L–1TRE, 相较耐热型品种HM33能够显著增强敏感型品种FM5抗逆性, 降低MDA含量, 提高SOD、POD和CAT活性, 从而减缓了小麦旗叶的衰老速率, 使旗叶具有较高的光合物质生产能力, 这将有利于籽粒的灌浆, 从而缓解高温胁迫带来的产量降低。有研究指出, TRE提高植物耐热性是通过保持较高的抗氧化酶活性, 提高植物的抗氧化胁迫能力, 降低高温引起的氧化胁迫对植株的伤害, 维持细胞膜的稳定性, 从而提高耐热性[44]。也有研究指出, 胁迫状态下TRE可能为某响应机制的“开关”, 进而影响氧化酶含量, 增加叶片中蛋白质、叶绿素等生理物质的含量, 促进叶片光合作用, 进而有利于地上部叶片和地下部根系的协调生长[45]。本试验中TRE对高温胁迫条件下小麦所表现出的这些生理效应说明TRE预处理可以减弱高温胁迫下小麦植株的氧化胁迫程度, 提高了小麦对高温胁迫的抵抗能力。
4 结论
敏感型品种泛麦5号灌浆期高温胁迫下产量降低幅度大于耐热型品种淮麦33, 千粒重的降低是主要减产原因。倒二叶干物质积累量降幅最为明显, 喷施浓度15 mmol L–1海藻糖相较高温胁迫下喷施清水对照, 旗叶干物质积累量降幅缓解效果最好, 因为旗叶抗氧化酶活性提高, 进而导致丙二醛含量下降, 过氧化程度有所缓解, 光合器官得到保护, 从而使旗叶绿叶面积和叶绿素相对含量提高, 净光合速率、气孔导度和蒸腾速率有所提升, 光合能力提高。且喷施海藻糖缓解小麦灌浆期高温胁迫效果最好。本试验条件下, 开花期喷施浓度15 mmol L–1TRE能在一定程度上缓解高温影响, 生产中可以施用, 但仍不能完全弥补高温损失。
[1] 张玉. 浅析小麦种植技术推广意义及有效途径. 种子科技, 2018, 36(5): 1. Zhang Y. Analysis on the significance and effective way of popularizing wheat planting technology., 2018, 36(5): 1 (in Chinese).
[2] 缪文卿. 确保口粮绝对安全的思考. 当代农村财经, 2021, (5): 41–42. Miu W Q. Thoughts on ensuring the absolute safety of rations., 2021, (5): 41–42 (in Chinese).
[3] Chen Z, Galli M, Gallavotti A. Mechanisms of temperature regulated growth and thermotolerance in crop species., 2022, 65: 102–134.
[4] 曹彩云, 党红凯, 郑春莲, 郭丽, 李科江, 马俊永. 灌浆期高温胁迫对小麦灌浆的影响及叶面喷剂的缓解作用. 中国生态农业学报, 2016, 24: 1103–1113. Cao C Y, Dang H K, Zheng C L, Guo L, Li K J, Ma J Y. Impact of high temperature stress on grain-filling and the relief effect of foliage sprays during grain-filling stage of wheat., 2016, 24: 1103–1113 (in Chinese with English abstract).
[5] 李世平. 冬小麦耐热性及相关性状数量位点遗传剖析. 西北农林科技大学博士学位论文, 陕西西安, 2013. Li S P. Genetic Dissection of Quantitative Traits Loci Fir Heat Tolerance Index and Traits Associated with Heat Tolerance in Winter Wheat (L.). PhD Dissertation of Northwest A&F University, Xi’an, Shaanxi, China, 2013 (in Chinese with English abstract).
[6] 滕中华, 智丽, 宗学凤, 王三根, 何光华. 高温胁迫对水稻灌浆结实期叶绿素荧光,抗活性氧活力和稻米品质的影响. 作物学报, 2008, 34: 1662–1666.Teng Z H, Zhi L, Zong X F, Wang S G, He G H. Effects of high temperature on chlorophyll fluorescence, active oxygen resistance activity, and grain quality in grain-filling periods in rice plants., 2008, 34: 1662–1666 (in Chinese with English abstract).
[7] 岳鹏莉. 花后高温干旱胁迫对冬小麦淀粉合成特性的影响. 河南农业大学硕士学位论文, 河南郑州, 2017. Yue P L. Effect of High Temperature and Drought Stress on Starch Synthesis Characteristics after Flowering of Winter Wheat. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2017 (in Chinese with English abstract).
[8] 石晓丽, 史文娇. 极端高温对黄淮海平原冬小麦产量的影响. 生态与农村环境学报, 2016, 32: 259–269. Shi X L, Shi W J. Impacts of extreme high temperature on winter wheat yield in the Huang-Huai-Hai plain., 2016, 32: 259–269 (in Chinese with English abstract).
[9] 严菊芳. 干旱条件下冷型小麦的农田热量平衡及小气候特征. 西北农林科技大学硕士学位论文, 陕西西安, 2009. Yan J F. Heat balance of cold type wheat under drought conditions and microclimate characteristics. MS Thesis of Northwest A&F University, Xi’an, Shaanxi, China, 2009 (in Chinese with English abstract).
[10] Müller J, Aeschbacher R A, Sprenger N, Wiemken B A. Disaccharide- mediated regulation of sucrose: fructan-6-fructosyltransferase, a key enzyme of fructan synthesis in barley leaves., 2000, 123(1): 265–273.
[11] Guido V, Oliver F, Louis J R D B, Thomas B, Andres W, Aeschbacher R A, Astrid W. Trehalose metabolism in Arabidopsis: occurrence of trehalose and molecular cloning and characterization of trehalose-6-phosphate synthase homologues., 2001, 52: 1817–1826.
[12] Garg A K. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses., 2003, 99: 15898–15903.
[13] Schluepmann H, Dijken A V, Aghdasi M, Wobbes B, Smeekens P S. Trehalose mediated growth inhibition ofseedlings is due to trehalose-6-phosphate accumulation., 2004, 135: 879–890.
[14] Abdallah M S, Abdelgawad Z A, El-Bassiouny H. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties., 2016, 103: 275–282.
[15] 周林, 郭尚, 杨臻, 杨杰, 王华, 南晓洁. 外源海藻糖对高温胁迫下双孢蘑菇菌株生长的促进效应与耐高温株系筛选. 西南农业学报, 2019, 32(1): 154–160. Zhou L, Guo S, Yang Z, Yang J, Wang H, Nan X J. Effect of exogenous trehalose on growth ofstrain under high temperature stress and screening of high temperature resistant strains., 2019, 32(1): 154–160 (in Chinese with English abstract).
[16] 庞椿朋, 叶亮, 马健, 路涛, 杨宗艺, 齐明芳. 海藻糖对高温下番茄幼苗叶片光合作用的调控作用. 江苏农业科学, 2022, 45(21): 143–146.Pang C P, Ye L, Ma J, Lu T, Yang Z Y, Qi M F. Effects of trehalose on photosynthesis of tomato seedling leaves under high temperature., 2022, 45(21): 143–146 (in Chinese).
[17] 樊永惠, 李宇星, 马亮亮, 吕钊彦, 武倩倩, 张文静, 马尚宇, 黄正来. 灌浆期高温胁迫下外源水杨酸对小麦旗叶抗氧化生理特性的影响. 核农学报, 2022, 36: 1878–1886. Fan Y H, Li Y X, Ma L L, Lyu Z Y, Wu Q Q, Zhang W J, Ma S Y, Huang Z L. Effects of exogenous salicylic acid on antioxidant physiological characteristics of wheat flag leaves under high temperature stress at grain filling stage., 2022, 36: 1878–1886 (in Chinese with English abstract).
[18] 唐秀巧, 王静静, 龙媛媛, 翁颖, 陈浩然, 李金才, 宋有洪, 李金鹏. 花后喷施KH2PO4对灌浆期高温胁迫下小麦根系生理特性及产量的影响. 核农学报, 2022, 36: 1685–1691. Tang X Q, Wang J J, Long Y Y, Weng Y, Chen H R, Li J C, Song Y H, Li J P. Effect of spraying KH2PO4after anthesis on root physiological characteristics and yield of wheat under high temperature stress at grain filling stage., 2022, 36: 1685–1691 (in Chinese with English abstract).
[19] 杨璞, 高彩红. 外源一氧化氮对逆境胁迫下小麦的保护效应. 天津农业科学, 2020, 26(5): 27–31.Yang P, Gao C H. Protective effect of exogenous nitric oxide on wheat under adversity stress., 2020, 26(5): 27–31 (in Chinese with English abstract).
[20] 陈静, 姜敬贤, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控. 作物学报, 2022, 48: 1502–1515. Chen J, Jiang J X, Ren B C, Liu B, Liu P, Zhang J W. Regulation of leaf-spraying glycine betaine on yield formation and antioxidation of summer maize sowed in different dates., 2022, 48: 1502–1515 (in Chinese with English abstract).
[21] 杨东清, 王振林, 倪英丽, 尹燕枰, 蔡铁, 杨卫兵, 彭佃亮, 崔正勇, 江文文. 高温和外源ABA对不同持绿型小麦品种籽粒发育及内源激素含量的影响. 中国农业科学, 2014, 47: 2109–2125.Yang D Q, Wang Z L, Ni Y L, Yin Y P, Cai T, Yang W B, Peng D L, Cui Z Y, Jiang W W. Effect of high temperature stress and spraying exogenous ABA post-anthesis on grain filling and grain yield in different types of stay-green wheat., 2014, 47: 2109–2125 (in Chinese with English abstract).
[22] Lalit S, Chander S S. Tween-20 modified acacia nelotica andbiomass for enhanced biosorption of Cr(VI) in aqueous environment., 2015, 52: 41–53.
[23] Zheljazkov V D, Astatkie T, Jeliazkova E. Effect of foliar application of methyl jasmonate and extracts of juniper and sagebrush on essential oil yield and composition of ‘native’ spearmint., 2013, 48(4): 462–465.
[24] 王潭刚, 孙婷, 王冀川, 李慧琴, 高振, 石元强. 播期和密度对滴灌冬小麦群体结构与抗倒特性的影响. 浙江农业学报, 2021, 33: 193–202. Wang T G, Sun T, Wang J C, Li H Q, Gao Z, Shi Y Q. Effects of sowing date and density on population structure and lodging resistance of winter wheat under drip irrigation., 2021, 33: 193–202 (in Chinese with English abstract).
[25] Xu M Z, Liu X, Yu L Q, Liu S Q, Zeng L M. Physiological analysis on mechanisms of cold-tolerance of Dongxiang wild rice (I)., 2010, 38: 5012–5014 (in Chinese with English abstract).
[26] Giannopolitis C N, Ries S K. Superoxide dismutase: I. Occurrence in higher plants., 1972, 59: 309–314.
[27] 王伟玲, 王展, 王晶英. 植物过氧化物酶活性测定方法优化. 实验室研究与探索, 2010, 29(4): 21–23. Wang W L, Wang Z, Wang J Y. Optimization of determination method of peroxidase activity in plant., 2010, 29(4): 21–23 (in Chinese with English abstract).
[28] Hammerschmidt R, Nuckles E M, Ku J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium., 1982, 20: 73–76.
[29] 岳鹏莉, 王晨阳, 卢红芳, 刘卫星, 马耕, 王强, 胡阳阳. 灌浆期高温干旱胁迫对小麦籽粒淀粉积累的影响. 麦类作物学报, 2016, 36: 1489–1496. Yue P L, Wang C Y, Lu H F, Liu W X, Ma G, Wang Q, Hu Y Y. Effect of heat and drought stress on starch accumulation during grain filling stage., 2016, 36: 1489–1496 (in Chinese with English abstract).
[30] 曹彩云, 党红凯, 郑春莲, 李科江, 马俊永. 阶段高温对不同小麦品种产量的影响及其耐热性差异研究. 麦类作物学报, 2020, 40: 351–364. Cao C Y, Dang H K, Zheng C L, Li K J, Ma J Y. Impacts of high temperature in different periods on yield and evaluation of heat tolerance in wheat varieties., 2020, 40: 351–364 (in Chinese with English abstract).
[31] 吴翠平, 贺明荣, 张宾, 张洪华, 刘永环. 氮肥基追比与灌浆中期高温胁迫对小麦产量和品质的影响. 西北植物学报, 2007, 27: 4734–4739.Wu C P, He M R, Zhang B, Zhang H H, Liu Y H. Effects of nitrogen dressing ratios and heat stress during the middle period of grain filling on wheat grain yield and quality., 2007, 27: 4734–4739 (in Chinese with English abstract).
[32] 郭艳艳, 段巍巍. 不同冬小麦品种籽粒胚乳增殖和灌浆对粒重的影响. 麦类作物学报, 2018, 38(1): 84–89.Guo Y Y, Duan W W. Studies on endosperm development and grain filling characteristics in different winter wheat varieties., 2018, 38(1): 84–89 (in Chinese with English abstract).
[33] 刘书仁, 郭世荣, 程玉静, 刘超杰, 王丽萍, 束胜. 外源脯氨酸对高温胁迫下黄瓜幼苗叶片AsA-GSH循环和光合荧光特性的影响. 西北植物学报, 2010, 30: 309–316. Liu S R, Guo S R, Cheng Y J, Liu C J, Wang L P, Shu S. Effects of exogenous proline on the ascorbat-glutahione cycle and photosynthetic fluorescence characteristics in leaves of cucumber seedlings under high temperature stress., 2010, 30: 309–316 (in Chinese with English abstract).
[34] 李哲, 姜沣益, 代梦雪, 李宇星, 张文静, 马尚宇, 黄正来, 樊永惠. 外源海藻糖对高温胁迫小麦干物质积累和籽粒灌浆的影响. 麦类作物学报, 2022, 42: 614–622. Li Z, Jiang F Y, Dai M X, Li Y X, Zhang W J, Ma S Y, Huang Z L, Fan Y H. Effect of exogenous trehalose on dry matter accumulation and grain filling of wheat under heat stress., 2022, 42: 614–622 (in Chinese with English abstract).
[35] 罗宏海, 李俊华, 张宏芝, 何在菊, 勾玲, 张旺锋. 源库调节对新疆高产棉花产量形成期光合产物生产与分配的影响. 棉花学报, 2009, 21: 371–377. Luo H H, Li J H, Zhang H Z, He Z J, Gou L, Zhang W F. Effects of source and sink manipulation on transportation and allocation of leaf photosynthetic products during flowering and boll-setting stage in high-yield cotton of Xinjiang., 2009, 21: 371–377 (in Chinese with English abstract).
[36] 何丽香, 傅兆麟, 宫晶, 孙常玉. 小麦灌浆期上三叶叶绿素含量与产量和品质的关系. 中国农学通报, 2014, 30(15): 183–187.He L X, Fu Z L, Gong J, Sun C Y. Relationship between chlorophyll contention of top-three leaves at different times of grain filling stage and grain yield and quality traits in wheat., 2014, 30(15): 183–187 (in Chinese with English abstract).
[37] 王磊. 硫化氢信号对干旱胁迫下十字花科植物光合作用的调节. 山西大学硕士学位论文, 山西太原, 2020. Wang L. Regulation of Hydrogen Sulfide Signal on Photosynthesis of Brassicaceae Plants under Drought Stress. MS Thesis of Shanxi University, Taiyuan, Shanxi, China, 2020 (in Chinese with English abstract).
[38] 胡凡波, 刘玲, 隆小华, 刘兆普. 外源NO对NaCl胁迫下长春花幼苗生物量和叶绿素荧光的影响. 生态学杂志, 2011, 30: 1620–1626. Hu F B, Liu L, Long X H, Liu Z P. Effects of exogenous nitric oxide on biomass and chlorophyll fluorescence of catharanthus roseus seedlings under NaCl stress., 2011, 30: 1620–1626 (in Chinese with English abstract).
[39] 姜磊, 李焕勇, 张芹, 张会龙, 乔艳辉, 张华新, 杨秀艳. AM真菌对盐碱胁迫下杜梨幼苗生长与生理代谢的影响. 南京林业大学学报: 自然科学版, 2020, 44(6): 152–160. Jiang L, Li H Y, Zhang Q, Zhang H L, Qiao Y H, Zhang H X, Yang X Y. Effects of arbuscular mycorrhiza fungi on the growth and physiological metabolism ofBunge seedlings under saline-alkaline stress., 2020, 44(6): 152–160 (in Chinese with English abstract).
[40] 李金花, 张春艳, 刘浩, 李永春. 海藻糖的特性及其在植物抗逆中的应用. 江西农业学报, 2011, 23(6): 25–27. Li J H, Zhang C Y, Liu H, Li Y C. Characteristics of trehalose and its application in improving stress tolerance of plants., 2011, 23(6): 25–27 (in Chinese with English abstract).
[41] 赵文静. 高低温胁迫对紫花苜蓿幼苗光合作用和光保护机制的影响, 新疆农业大学硕士学位论文, 新疆乌鲁木齐, 2021. Zhao W J. Effects of High and Low Temperature Stress on Photosynthesis and Photoprotection Mechanism ofat seedings period. MS Thesis of Xinjiang Agricultural University,Urumqi, Xinjiang, China, 2021 (in Chinese with English abstract).
[42] 尹赜鹏, 鹿嘉智, 高振华, 齐明芳, 孟思达, 李天来. 番茄幼苗叶片光合作用、PSII电子传递及活性氧对短期高温胁迫的响应. 北方园艺, 2019, (5): 1–11. Yin Z P, Lu J Z, Gao Z H, Qi M F, Meng S D, Li T L. Effects of Photosynthetic, PSII electron transport and reactive oxygen species on short-term high temperature stress in tomato seedlings., 2019, (5): 1–11 (in Chinese with English abstract).
[43] 张桂莲, 陈立云, 张顺堂, 肖应辉, 贺治洲, 雷东阳. 高温胁迫对水稻剑叶保护酶活性和膜透性的影响. 作物学报, 2006, 32: 1306–1310. Zhang G L, Chen L Y, Zhang S T, Xiao Y H, He Z Z, Lei D Y. Effect of high temperature stress on protective enzyme activities and membrane permeability of flag leaf in rice., 2006, 32: 1306–1310 (in Chinese with English abstract).
[44] 罗音. 外源海藻糖提高小麦耐热性的生理机制研究. 山东农业大学博士学位论文, 山东泰安, 2007.Luo Y. Studies on the Physiological Mechanisms of Heat Tolerance in Wheat Improved by Exogenous Trehalose. PhD Dissertation of Shandong Agricultural University, Tai’an, Shandong, China, 2007 (in Chinese with English abstract).
[45] 柳福智, 张迎芳, 陈垣. 外源海藻糖对NaHCO3胁迫下甘草幼苗生长调节及总黄酮含量的影响. 草业学报, 2021, 30(7): 148–156.Liu F Z, Zhang Y F, Chen Y. Effect of exogenous trehalose on growth regulation and total flavonoid content ofseedlings under NaHCO3stress., 2021, 30(7): 148–156 (in Chinese with English abstract).
Effects of exogenous trehalose on physiological characteristics and yield of wheat flag leaves under high temperature stress at grain filling stage
LI Yu-Xing, MA Liang-Liang, ZHANG Yue, QIN Bo-Ya, ZHANG Wen-Jing, MA Shang-Yu, HUANG Zheng-Lai*, and FAN Yong-Hui*
College of Agriculture, Anhui Agricultural University / Key Laboratory of Wheat Biology and Genetic Breeding in the South of Huanghe-Huaihe Rivers, Ministry of Agriculture and Rural Affairs, Hefei 230036, Anhui, China
In order to investigate the effect of exogenous trehalose (TRE) on the physiological characteristics and yield of flag leaves of wheat varieties with different tolerance under high temperature stress during grain filling period, the experiments were conducted in the high-tech agricultural park of Anhui Agricultural University from 2020 to 2022. The sensitive wheat varieties Fanmai 5 (FM5) and heat-resistant wheat varieties Huaimai 33 (HM33) with significant difference in heat resistance were selected as the experimental materials, there were five treatments of spraying water + non high temperature (CK1), water + high temperature stress (CK2), 10 mmol L–1trehalose + high temperature stress (T10H), 15 mmol L–1trehalose + high temperature stress (T15H), and 20 mmol L–1trehalose + high temperature stress (T20H) on the leaf surface. The results showed that under high temperature stress conditions, compared to non-high temperature adversity, the green leaf area, chlorophyll relative content (SPAD), and dry matter accumulation were significantly decreased, wheat yield was significantly reduced, spike number, and spike grain number did not change significantly, and 1000-grain weight was the dominant factor in yield reduction. Compared to water spraying, the yield after spraying trehalose was higher than that under high temperature stress, the accumulation of dry matter in various organs was increased, the content of malondialdehyde (MDA) was decreased, the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) were increased, and the range of change was different among varieties. The decrease of SPAD value of heat-resistant varieties was smaller than that of malondialdehyde, but the increase of CAT activity was larger, indicating the production reduction was small. Further analysis showed that the net photosynthetic rate (n), stomatal conductance (s), transpiration rate (r), SPAD, SOD, and POD had an extremely significant positive correlation with 1000-grain weight, while MDA had an extremely significant negative correlation with yield. These results suggested that high temperature stress inhibited photosynthesis in wheat by reducingn,s, andr, decreasing photosynthetic product production and causing eventual yield reduction. Spraying TRE,n,s,r, SPAD, and CAT were the main indicators to reduce the injury caused by high temperature stress, and the mitigation effect of sensitive varieties was obvious. In addition, the effect of spraying 15 mmol L–1trehalose at flowering stage was the best. The results could provide a theoretical basis for the mechanism of wheat photosynthetic response difference and wheat antioxidant metabolism under high temperature stress during trehalose filling stage.
wheat; high temperature stress; trehalose; flag leaf; yield
2023-02-10;
2023-02-27.
10.3724/SP.J.1006.2023.21057
通信作者(Corresponding authors):黄正来, E-mail: xdnyyjs@163.com; 樊永惠, E-mail: yonghuifan@ahau.edu.cn
E-mail: 18702289708@163.com
2022-08-28;
本研究由安徽省科技重大专项(202003a06020014), 安徽省自然科学基金项目(2008085QC118), 国家自然科学基金联合基金项目(U19A2021), 安徽农业大学稳定和引进人才科研项目(YJ2018-38)和安徽农业大学青年基金重点项目(03087060)资助。
This study was supported by the Major Science and Technology Projects of Anhui Province (202003a06020014), the Natural Science Foundation of Anhui Province (2008085qc118), the Joint Fund Project of the National Natural Science Foundation of China (U19A2021), the Scientific Research Project for Stabilizing and Introducing Talents of Anhui Agricultural University (YJ2018-38), and the Key Project of the Youth Fund of Anhui Agricultural University (03087060).
URL: https://kns.cnki.net/kcms/detail//11.1809.S.20230223.1648.009.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年9期)2022-07-18
舰船科学技术(2022年11期)2022-07-15
农业工程学报(2022年7期)2022-07-09
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
浙江农业科学(2019年11期)2019-11-09
西藏农业科技(2019年3期)2019-11-04
今日农业(2019年11期)2019-08-15
