
Siwalik plant megafossil diversity in the Eastern Himalayas: A review
2023-07-11 08:20MhsinAliKhnSumnMhtoRoertSpicerTeresSpicerAshifAliTposhiHzrSuirBer
植物多样性 2023年3期
Mhsin Ali Khn , Sumn Mhto , Roert A.Spicer , Teres E.V.Spicer ,Ashif Ali , Tposhi Hzr , Suir Ber
a Palaeobotany and Palynology Laboratory, Department of Botany, Sidho-Kanho-Birsha University, Ranchi Road, Purulia 723104, India
b CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666303, PR China
c School of Environment, Earth and Ecosystem Sciences, The Open University, Milton Keynes, MK7 6AA, UK
d Centre of Advanced Study, Department of Botany, University of Calcutta, 35, B.C.Road, Kolkata 700019, India
Keywords:Megafossils Siwalik Miocene-Pleistocene Palaeovegetation Palaeoenvironment Eastern Himalayas
A B S T R A C T The Eastern Himalayas are renowned for their high plant diversity.To understand how this modern botanical richness formed, it is critical to investigate past plant biodiversity preserved as fossils throughout the eastern Himalayan Siwalik succession (middle Miocene-early Pleistocene).Here, we present a summary of plant diversity records that document Neogene floristic and climate changes.We do this by compiling published records of megafossil plant remains,because these offer better spatial and temporal resolution than do palynological records.Analyses of the Siwalik floral assemblages based on the distribution of the nearest living relative taxa suggest that a tropical wet evergreen forest was growing in a warm humid monsoonal climate at the deposition time.This qualitative interpretation is also corroborated by published CLAMP (Climate Leaf Analysis Multivariate Program) analyses.Here, we also reconstruct the climate by applying a new common proxy WorldClim2 calibration.This allows the detection of subtle climate differences between floral assemblages free of artefacts introduced by using different methodologies and climate calibrations.An analysis of the Siwalik floras indicates that there was a gradual change in floral composition.The lower Siwalik assemblages provide evidence of a predominance of evergreen elements.An increase in deciduous elements in the floral composition is noticed towards the close of the middle Siwalik and the beginning of the upper Siwalik formation.This change reflects a climatic difference between Miocene and Plio-Pleistocene times.This review helps us to understand under what paleoenvironmental conditions plant diversity occurred and evolved in the eastern Himalayas throughout the Cenozoic.
1.Introduction
‘Siwalik’sediments comprise a thick(about 7000 m)succession of Neogene predominantly freshwater coarsely bedded sandstone,siltstone, clay,and conglomeratic molassic deposits exposed along the length of the Himalayan foothills from the Potwar Plateau of Pakistan in the west to Assam in the east(Parkash et al.,1980;Bora and Shukla, 2005; Chakrabarti, 2016).They were deposited in a variety of fluvial environments, including piedmonts, outwash plains, channels, floodplains, and oxbow lakes, although some record marine influence (Taral et al., 2019; Debnath et al., 2021).They have accumulated close to sea level in a long but narrow foredeep to the south of the rising Himalayas since the middle Miocene time (Bora and Shukla, 2005; Chakrabarti, 2016).The Siwalik succession is generally subdivided into three subgroups,namely the lower, middle, and upper Siwaliks, and their ages are assigned to middle Miocene,early Pliocene-late Pliocene,and late Pliocene-early Pleistocene respectively (Pilgrim, 1910, 1913;Johnson et al.,1985;Ranga Rao et al.,1988;Valdiya,2002).During the latest phase of the rise of the Himalayas,in Pleistocene to recent times,‘Siwalik’sediments were uplifted,folded,and faulted to form a continuous mountain range of relatively low height ranging from 1000-1200 m a.s.l., 2400 km in length and 20-25 km in width.From west to east along their length the Siwaliks have been divided into seven sectors: Jammu, Himachal, Uttarakhand, Nepal, Darjeeling, Bhutan, and the Southeastern Himalaya (Fig.1; Karunakaran and Ranga Rao, 1976; Ranga Rao et al., 1979).Compared to their outcrops in the western and central Himalayas, the eastern Himalayan Siwaliks occur as a thinner and discontinuous belt.The Darjeeling, Bhutan, and Southeastern Himalaya Siwalik sectors of the eastern Himalayas are the focus of the present review.
Although a rich vertebrate fauna has been reported from eastern Himalayan Siwalik sediments(Medlicott,1865;Pilgrim,1910,1913;Singh, 1975, 1983; Acharyya et al., 1987), which has helped in establishing the stratigraphy, classification, and depositional environments of the Siwalik Group of rocks, comparatively little systematic work has been carried out on the plant fossils entombed in these beds.Here, we explore the floral composition within the different units of the Siwalik succession in the eastern Himalayan sectors and trace how the climate and floras have changed through time from the middle Miocene through the Pleistocene to the present using a common proxy framework that allows direct climatic comparison between fossil sites free of artefacts that might otherwise be introduced by using a range of proxies differently calibrated.
The Siwalik floras offer considerable potential for studies of Neogene vegetation and climate change, including monsoon signatures, elevation changes within the Himalayan and Tibetan region, and plant biogeography since the middle Miocene (Prasad,2008; Khan et al., 2014a, 2019a).The recovered fossil floras from the different eastern Siwalik sectors comprise mainly leaves of woody dicot angiosperms (Mehrotra et al., 1999; Prasad et al.,1999, 2004; Prasad and Tripathi, 2000; Prasad, 2008; Khan et al.,2014a; Srivastava et al., 2018).Here, we review fossil floras recovered from the Darjeeling, Bhutan, and Southeastern Himalaya Siwalik sectors of the eastern Himalayas and reconstruct the climate by applying a new common proxy and calibration to all the currently available eastern Siwalik fossil floras.The use of a common proxy with the same calibration has been lacking thus far as different authors tend to apply different methodologies and the methodologies themselves have evolved over time.This mix of approaches means detection and quantification of subtle changes in climate over time is hard to achieve and ambiguous at best.By using a common analytical framework, we provide insights into the monsoon evolution during Siwalik depositional period in the eastern Himalayas.
In the present assessment,we compile all palaeobotanical plant data from published literature to document plant diversity in the eastern Himalayas throughout the Siwalik succession.We consider only fossil taxa represented by megafossils (leaves, wood, fruits,fruiting calyx, and seeds).Potential megafossils, especially leaves,cannot travel far from their point of origin before fossilization and remain identifiable(Ferguson,1985;Spicer and Wolfe,1987;Spicer,1991) and are destroyed when the sediments hosting them are reworked.Pollen and spores (microfossils), however, can travel long distances up and down the slope prior to final burial and still appear pristine, and can be reworked many times.Moreover, they often cannot be identified to a fine enough taxonomic resolution using only a light microscope(Ferguson et al.,2007).By focusing on megafossils we ensure that we are reconstructing vegetation local to the fossil site, and so potentially are able to detect fine-scale changes in space and time.
We first briefly introduce the present-day floristic composition in the eastern Himalayas, then present known fossil families, genera,and species to depict the diversity of Siwalik plants and ecosystems,and then analyze the inferred floristic characters and changes.We also summarize the Siwalik climate evolution in the eastern Himalayas.The present review aims to facilitate access to the rich Siwalik palaeobotanical record of the eastern Himalayas.We also reexamine the eastern Himalayan Siwalik Miocene-Pleistocene climate and introduce new quantitative proxy palaeo-humidity measurements in order to characterize better the eastern Himalayan environment during a critical time of Himalayan uplift and associated evolution(Ding et al.,2017;Bhatia et al.,2021,2022).
2.Geological setting

Fig.1.Maps showing the seven sectors of the Siwalik belt (modified after Karunakaran and Ranga Rao,1976) showing the locations of the present study areas.

Table 1 A generalised lithostratigraphy of Siwalik sediments in the eastern Himalayas (modified after Khan et al., 2014a; 2019b; Taral et al., 2017).
Pilgrim (1913) divided the Siwalik succession into three units,namely the lower(middle Miocene),middle(late Miocene to early Pliocene), and upper (late Pliocene to early Pleistocene).The Siwalik sediments are characterized by alternating sandstone and mudstone facies, with the finer sediments very often containing abundant biota (Chakrabarti, 2016).In general Siwalik sediments exhibit a coarsening trend over time, but even the youngest units contain fine-grained beds that are fossiliferous.Here,we represent a generalized lithostratigraphy of the Siwalik sediments exposed in the eastern Himalayas(Table 1;Figs.1 and 2).In the Darjeeling area,this is based on Ganguly and Rao(1970),Acharya(1994),and Taral et al.(2017); in Bhutan, we refer to Coutand et al.(2016), while in the Southeastern Himalaya we rely on Anand-Prakesh and Singh(2000) and Singh (2007).The ages of the sedimentary succession in different Siwalik sectors of the eastern Himalayas have been quantified by means of magnetostratigraphy(Chirouze et al.,2012;Coutand et al., 2016).Magnetostratigraphic correlations indicate that the Bhutan Siwalik Group was deposited during the latest Miocene and the Pleistocene, between ~7 million years ago (Ma)and ~1 Ma, and that the boundary between lower and middle Siwaliks can be dated to ~6 Ma, the middle to upper Siwalik to 3.8 Ma,and the top of the section as~1 Ma(Coutand et al.,2016).In the Southeastern Himalaya, the Siwalik Group was deposited between 13 and 2.5 Ma, with the transition between the lower and middle Siwaliks dated at about 10.5 Ma and the middle to upper Siwalik transition at 2.6 Ma (Chirouze et al., 2012).The boundary age differences exhibited among the different parts of the eastern Himalayas could be due to temporal and locational changes in the various loci of deposition as well as local erosion events.
3.Modern floristic composition of the eastern Himalayas
To contextualize megafossil records from the eastern Himalayas,it is useful to consider the rich and diverse modern flora of the region.At elevations similar to those inferred for the source vegetation of the Siwalik fossil assemblages,modern vegetation close to the fossil localities in the eastern Himalayas is characterized as warm humid tropical (Champion and Seth, 1968; Biswas et al.,1976; Grierson and Long, 1983; Kaul and Haridasan, 1987; Hazra et al.,1996; Baishya et al., 2001).
The eastern Himalayas are considered ‘crisis ecoregions’and‘biodiversity hotspots’(Brooks et al., 2006).This region is also a meeting ground for the Indo-Malayan, Palearctic, and Sino-Japanese biogeographical realms, and has diverse biota as well as diverse ecological and elevational gradients(CEPF,2005,2007).The complex topography and extreme elevational gradients from less than 300 m (tropical lowlands) to more than 8000 m (high mountains) have led to the development of a variety of floristic as well as vegetation patterns.Climate-dependent vegetation is largely determined by decreasing moisture and temperature with increasing elevations (thermal and hygrometric terrestrial lapse rates), which vary through time, affording opportunities for plant migrations, novel juxtapositions, and speciation.The Himalayan range to the north acts as a barrier to the southwest monsoon from the Bay of Bengal, causing the moisture regime to decrease westwards along the Siwaliks, and comparatively more rain is received in the East.The complex mountain topography creates diverse bioclimatic zones, an exceptionally rich biodiversity assemblage,and‘sky’island conditions for many species.Broadly,vegetation in the eastern Himalayas can be categorized into tropical,sub-tropical,warm temperate, cool temperate, sub-alpine, and alpine types in ascending order based on parameters such as physiognomy, floral composition, habitat conditions, and physiography (WWF and ICIMOD 2001).
Takhtajan(1969)regarded the eastern Himalayas as the‘cradle’of flowering plants.The region is also well known for its botanically curious and rare species (e.g., Sapria himalayana Griff., Rafflesiaceae).Nearly 50% of the total flowering plants recorded in India are from the northeastern region.The genus Rhododendron(Ericaceae) is a remarkable taxon of showy plants, with most confined to this region and a substantial number of endemic species(Pradhan and Lachungpa,1990).The eastern Himalayan region is rich in endemic floras and many species have value as medicinal or edible plants (Sundriyal,1999).

Fig.2.Fossiliferous Siwalik exposures of the eastern Himalayas (a: Darjeeling; b: Bhutan; c: Southeastern Himalaya).
Several species,namely Shorea robusta C.F.Gaertn.,Mesua ferrea L., Syzygium cumini (L.) Skeels, Terminalia paniculata Roth., are dominant in northern tropical wet evergreen forests (150 m altitude); Magnolia L., Terminalia elliptica Willd., and Bombax ceiba L.in northern sub-tropical semi-evergreen forests (150-230 m);
Chukrasia tabularis A.Juss., Gmelina arborea Roxb., Rhododendron arboretum Sm., Dalbergia sissoo Roxb.in North India moist deciduous forests(230-300 m);Madhuca longifolia(J.Konig)J.F.Macbr.,
Gordonia chilaunia, T.elliptica,Gossypium L.,Toona ciliata M.Roem.and Cinnamomum glaucescens (Nees) Hand.-Mazz.in Northern sub-tropical broad-leaved wet forests (300-1650 m altitude);Senegalia catechu (L.f.) P.J.H.Hurter & Mabb., Abies densa Griff.,Tsuga canadensis Carri`ere, Acer L.in northern montane wet temperature forests(1650-3000 m);M.longifolia(J.Konig)J.F.Macbr.,
Schima wallichii (DC.) Korth., Castanopsis indica (Roxb.ex Lindl.) A.DC.,T.elliptica,Duabanga grandiflora Walp.,Cassia fistula L.,Annona muricata L.in east Himalayan moist temperature forests(1500-1800 m) and Ficus religiosa L., Acer, Exbucklandia populnea(R.Br.ex Griff.) R.W.Br., Ceiba pentandra (L.) Gaertn., Prunus undulata Buch.Ham.ex D.Don, Castanopsis (D.Don) Spach,Rhododendron L., Salix L.in sub-alpine forests (3000-3660 m)(Grierson and Long,1983; Kaul and Haridasan,1987; Hazra et al.,1996; Baishya et al., 2001).
In and around the fossil localities the principal constituents of tropical moist semi-evergreen to deciduous forests are Pongamia pinnata(L.)Pierre,Bauhinia purpurea L.,Albizia sp.L.,Dalbergia sisso(Fabaceae); D.grandiflora (D.C.) Walp., Lagerstroemia parviflora Roxb.(Lythraceae); Terminalia catappa L., T.chebula Retz., T.miocarpa F.Muell.(Combretaceae); Litsea sp.Lam., Cinnamomum bejolghota (Buch.-Ham.) Sweet, Actinodaphne angustifolia (Blume)Nees, A.obovata (Nees) Blume, Phoebe goalparensis Hutch.(Lauraceae); Gynocardia odorata R.Br.(Achariaceae); Calophyllum polyanthum Wall.ex Choisy(Calophyllaceae);Bombax malabaricum D.C.(Malvaceae); Macaranga denticulata (Blume) Müll.Arg., Mallotus Lour.(Euphorbiaceae); Knema Lour.(Myristicaceae); Elaeocarpus aristatus Roxb.(Elaeocarpaceae); Shorea robusta (Dipterocarpaceae); Gynocardia arborea Roxb.ex Sm.(Achariaceae); Vitex quinata(Lour.)F.N.Williams(Lamiaceae);Dillenia pentagyna Roxb.(Dilleniaceae);Sterculia villosa Roxb.ex Sm.,Grewia eriocarpa Juss.,
Grewia zizyphifolia Baill.(Malvaceae); Garuga pinnata Roxb.(Burseraceae); Meliosma simplicifolia (Roxb.) Walp.(Sabiaceae); Spondias axillaris Roxb.B.L.Burtt & A.W.Hill (Anacardiaceae).
4.Research history of Palaeobotany in eastern Himalayan Siwalik
The eastern Himalayas hosts many Cenozoic sedimentary basins that have yielded Siwalik deposits that bear micro and mega plant fossils (Chakrobarty et al., 2020).The Lower, Middle, and Upper Siwalik floral archives range from the middle Miocene to the early Pleistocene(Table 1).The first information about the occurrence of plant fossils in the eastern Himalaya Siwaliks dates back to 1969 when Pathak (1969) initially described a few fragmentary angiosperm leaves from the middle Siwalik sediments of the Mahanadi section of Darjeeling.In subsequent years other geologists and paleobotanists also noted the presence of plant remains in the Siwalik sediments of the eastern Himalayas(Antal and Awasthi,1993;Antal and Prasad,1995,1996a, b, c,1997,1998).During the last few decades plant fossil localities have increased and valuable contributions have been made to the knowledge of the Siwalik paleobotany of the eastern Himalayas(Singh and Prakash,1980;Mehrotra et al.,1999; Prasad and Tripathi, 2000; Joshi and Mehrotra, 2003, 2007;Mitra and Banerjee,2004;Khan et al.,2007,2008,2009,2011,2014a,b,c,2015a,b,2016,2017a,b,2018a,b,2019a,b;Tripathi et al.,2007;Srivastava and Mehrotra,2009;Khan and Bera,2010,2014a,b,2016,2017; Prasad et al., 2015; Mehrotra et al., 2018; More et al., 2018;Srivastava et al., 2018).Rich and varied assemblages of plant megafossil, including leaf impressions, compressions, fruits, seeds, and woods, are now known and provide a sufficient basis for tracking regional environmental changes through time and space.Qualitatively, the present-day distribution of the taxonomically nearest living relatives (NLRs) of the Siwalik plant fossils suggests the existence of a tropical evergreen type of forest throughout the eastern Himalaya lowlands during the period of deposition.In recent years plant-based quantitative techniques(CLAMP-Climate Leaf Analysis Multivariate Program and CoA - Coexistence Approach) have been used to derive past climate parameters(temperature,humidity,and precipitation) and chart monsoon evolution (Khan et al., 2014a,2019b; Srivastava et al.,2021; Bhatia et al.,2022).
5.Palaeofloristic composition
This review is based on Siwalik (middle Miocene-early Pleistocene)plant megafossil assemblages recovered from the road and river exposures at different locations within the eastern Himalayas.The fossiliferous localities in the Darjeeling Siwalik sector are those of Oodlabari,Sevok road sections,and the Sevok bridge,Washbari,Gish, Lish, Churanthi, and Ramthi river sections; in Bhutan the Lakshmi and Darranga river sections and in the easternmost Siwalik sector, i.e., Southeastern Himalaya localities, the river cutting sections of the East Kameng district, the Bhalukpong, East Pinjoli and South Pinjoli road cutting sections of the West Kameng district, and the Naharlagun-Banderdewa, Nirjuli-Banderdewa,Itanagar-Banderdewa and Chandernagar-Gohpur road sections of the Papumpare district (Fig.2).Most leaf impressions were preserved in grey shales.Here, we review 219 plant fossil taxa, of which 102 taxa are from Darjeeling foothills,9 taxa are from Bhutan and 108 fossil taxa are from the Southeastern Himalaya.Most of the megafossil plant specimens were identified to the species level,but for a few specimens,necessary morphological characters were not preserved.Khan et al.(2015a, 2016, 2017a, b) studied cuticular epidermal features of some compressed leaves from the Southeastern sub-Himalaya and those are the most securely identified specimens.
There have been several contributions describing plant macrofossils from the eastern Himalaya Siwaliks, but so far none has synthesized our knowledge of the past flora and environment throughout Siwalik sedimentation.The recovered megaplant remains include compressions and impressions of leaves, fruiting calyxes, fruits, seeds, petrified and carbonized woods (Figs.3-5).The Siwalik floral assemblage is rich both in fossil quality and quantity and comprises 219 species belonging to 162 genera and 42 families (Tables 2 and S1-S5).Angiosperms are grouped into 159 genera within 39 families, the most abundant being the Fabaceae(represented by seventeen genera and twenty-four species), Dipterocarpaceae(four genera and thirteen species),Annonaceae(eight genera and eleven species), Lauraceae (seven genera and eleven species), Euphorbiaceae (six genera and seven species), Anacardiaceae (six genera and six species), Flacourtiaceae (four genera and five species),Rubiaceae(four genera and five species),Apocynaceae(four genera and four species)and Meliaceae(three genera and four species) (Tables S4 and S5).The eastern Himalayan Siwalik fossil floras consist of a wide variety of mostly woody plants listed in Tables S1-S3.Arborescent taxa dominate the assemblage, grass,and ferns being in a minority.Additionally,some workers reported in-situ occurrences of characteristic epiphyllous fungi on Siwalik leaf megafossils (Table S6).Reliable identification of the fossils is crucial for reliable palaeoclimatic and palaeoecological interpretation, thus, here some fossil specimens of uncertain affinities have been excluded from the listed fossil floras.The fossil specimens are held in the repository of the Palaeobotany-Palynology Section,Department of Botany, University of Calcutta.
5.1.The Lower Siwalik flora (middle Miocene)
The Lower Siwalik assemblage recovered from sediments exposed near Gish,Ramthi,Oodlabari,Sevok of Darjeeling,East and West Kameng of Southeastern Himalaya comprises mainly angiosperm plant remains attributable to 91 species within 65 genera belonging to 32 families (Table S1; Antal and Awasthi,1993; Antal et al., 1996; Antal and Prasad, 1996a; Joshi and Mehrotra, 2007;Khan et al., 2008; Srivastava and Mehrotra, 2009; Khan and Bera,2012, 2014a; Khan et al., 2015).In this assemblage, 90 leaf fossils,one dicot fossil wood,one Thelypteridaceae fern, and one gymnosperm taxon have been reported.Of these,22 species are new to the Siwalik palaeoflora and 19 have been identified as new to the Neogene flora of India.For example, Khan and Bera (2017)described Pinus on the basis of seed remains from the Dafla Formation exposed around the West Kameng district in the Southeastern Himalaya.This report provides the first-ever fossil record of Pinus winged seeds from India.It is obvious from the list of fossil taxa (Table S1) from the lower Siwalik eastern Himalaya assemblages that the family Fabaceae,represented by ten genera(Entada Adans.,Dalbergia L.f.,Derris Lour.,Millettia Wight&Arn.,Cynometra L., Bauhinia Plum.ex L., Albizia Durazz., Pongamia Adans., Acacia Mill.,and Mastertia L.),is the most dominant,followed by Lauraceae comprising four genera, Flacourtiaceae comprising four genera,Dipterocarpaceae comprising two genera, Annonacaae comprising two genera, Euphorbiaceae comprising of two genera and Combretaceae comprising two genera.The dominance of Fabaceae and the presence of Dipterocarpaceae is very significant from both palaeoecological and phytogeographical contexts.
5.2.The Middle Siwalik flora (late Miocene to Pliocene)
The Middle Siwalik assemblages recovered from sediments exposed from the fossil localities of Bhutan, Darjeeling, and Southeastern Himalaya represent mainly angiosperm plant remains currently comprising 81 species of 56 genera within 29 families (Table S2).Assignments are based mainly on leaf impressions.In this assemblage, 67 leaf fossils, four dicot fossil wood specimens, and one Thelypteridaceae fern have been described.They show closed affinity with extant thermophilic taxa such as Mitrephora Hook.f.& Thomson, Dipterocarpus C.F.Gaertn., Combretum Loef l., Millettia, Donax Lour.(Clinogyne grandis accepted name Donax canniformis), Shorea, Meiogyne Miq., Fissistigma Griff.,

Fig.3.(a) A fossil leaflet of Dysoxylum miocostulatum Khan and Bera (2007) from Lower Siwalik sediments of Southeastern Himalaya (CUH/PPL/P26) (Scale bar = 1 cm); (b, c)Winged seeds of Pinus arunachalensis Khan and Bera (2017) from Lower Siwalik sediments of Southeastern Himalaya (CUH/PPL/P/f/61a, b) (Scale bar = 1 cm); (d) A fossil leaf Quercus cf.lamellosa Khan et al.(2011) from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/46) (Scale bar= 1 cm); (e) Dysoxylum raptiensis Khan et al.(2015a)from Upper Siwalik sediments of Southeastern Himalaya (CUH/PPL/IB7/17) - abaxial cuticle showing stomata and epidermal cells (Scale bar = 10 μm); (f) Fossil leaf of Shorea mioobtusa Khan et al.(2016) from Lower Siwalik sediments of Southeastern Himalaya (CUH/PPL/P 83) (Scale Bar = 1 cm); (g) Dipterocarpus koilabasensis Khan et al.(2015a) from Upper Siwalik sediments of Southeastern Himalaya (CUH/PPL/IB7/3) - abaxial cuticle, hyphae with opposite and pointed appressoria of epiphyllous fungi Asterina sp.(Scale bar=10 μm);(h)Calophylloxylon eoinophyllum Khan et al.(2017a)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/W1A)-transverse section(T.S.)showing diffuse vessel distribution and solitary arrangement, and parenchyma bands (Scale bar = 10 μm); (i) Fossil fruit wing of Shorea mioassamica Khan and Bera (2010) from Lower Siwalik sediments of Southeastern Himalaya (CUH/PPL/P14) (Scale bar = 1 cm).
Gynocardia R.Br.,Vatica L.,and Garcinia L.(Antal and Prasad,1996a;Mehrotra et al., 1999; Prasad and Tripathi 2000; Tripathi et al.,2007; Prasad et al., 2015; Khan et al., 2016; 2017a, 2019).Of these, 17 species are new to the Siwalik flora and 15 have been identified as new to the Neogene flora of India.
Dipterocarpaceae, represented by 13 genera, is most dominant in the assemblage, followed by Annonaceae comprising four genera, Lauraceae comprising four genera, Sterculiaceae comprising two genera,and Calophyllaceae comprising two genera.
5.3.The Upper Siwalik flora (late Pliocene to early Pleistocene)

Fig.4.(a)A fossil leaf of Actinodaphne palaeoangustifolia Khan et al.(2011)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/40)(Scale bar=1 cm);(b)A fossil leaf of Calophyllum suraikholaensis Khan et al.(2009)from Middle Siwalik sediments of Southeastern Himalaya(CUH/PPL/B1) (Scale bar=1 cm);(c)Light micrographs of Gmelina siwalika Khan et al.(2018a)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/C3/44)-tangential longitudinal sections of the secondary xylem showing 2-3 seriate ray cells(Scale bar=50 μm);(d)Gynocardia arunachalensis Khan et al.(2014c)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/f/61a)-fossil seed showing thick, leathery seed coat (Scale bar = 1 cm); (e) Fossil leaf of Glochidion siwalikum Khan et al.(2019a) from Middle Siwalik sediments of Southeastern Himalaya (CUH/PPL/B/64A)(Scale bar=1 cm);(f)Cyathea siwalika Bera et al.(2014)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/TF/1)-Cyatheoid arrangement of vascular bundles within the leaf scar(scale bar=1 cm);(g)Calophyllum suraikholaensis Khan et al.(2015a)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/19)-lower cuticle showing paracytic stomata and epidermal cells (Scale bar = 10 μm).
The upper part of the Siwalik assemblage recovered from sediments exposed near Papumpare, East and West Kameng of Southeastern Himalaya is mainly represented by dicots comprising 47 species of 31 genera belonging to 21 families(Table S3).Assignments are based on both leaf impressions and compressions.An exception is that part of a compressed tree fern axis with leaf and adventitious root scars in the unusual arrangement has been described from the Plio-Pleistocene sediments of Southeastern Himalaya (Bera et al., 2014).This was the first macroscopic record of a cyatheaceous fern from the Indian Cenozoic.Other specimens show affinity with extant angiosperm taxa such as Dipterocarpus, Calophyllum L., Actinodaphne Nees,Shorea, Mastixia Blume, Gynocardia, Millettia, Knema Lour., Macaranga Thouars, Canarium L., Quercus L., Terminalia L., Croton L.,Gmelina L., Kayea Wall., Elaeocarpus L., and Pongamia (Bera et al.,2004; Joshi et al., 2003; Joshi and Mehrotra, 2007; Khan et al.,2011, 2015, 2016, 2017a, b; Khan and Bera, 2014a; Srivastava et al., 2018; Mehrotra et al., 2018).Lauraceae, represented by four genera, is the most dominant in this assemblage, followed by Fabaceae and Calophyllaceae represented by two genera.
6.Floristic changes throughout the Siwalik succession

Fig.5.(a) A fossil leaf of Persea preglaucescens Khan and Bera (2014) from Middle Siwalik sediments of Southeastern Himalaya (CUH/PPL/B/19) (Scale bar = 1 cm); (b) A wellpreserved wing-like persistent calyx lobe of Shorea bhalukpongensis Khan et al.(2016) from Middle Siwalik sediments of Southeastern Himalaya (CUH/PPL/B/f/19) showing characteristic parallel primary veins (green arrows) (Scale bar = 1 cm); (c) Light micrographs of Meliolinites neogenicus Khan et al.(2019c) from Upper Siwalik sediments of Southeastern Himalaya (CUH/PPL/IB7/36/AS1) - Hypha of M.neogenicus showing capitate appressoria with head cells and stalk cells (Scale Bar = 20 μm); (d) Dysoxylum raptiensis Khan et al.(2015a)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/17)-SEM of the abaxial cuticle,inner surface,paracytic stomata(Scale bar=10 μm);(e)Fossil leaf of Shorea Khan et al.(2019b)from Middle Siwalik sediments of Bhutan(CUH/PPL/BH/12A)(Scale bar=1 cm);(f)Fossil fruit of Dalbergia prelatifolia Khan and Bera(2014)from Lower Siwalik sediments of Darjeeling foothill (CUH/PPL/SV/f/1) (Scale bar = 1 cm); (g) Fossil fruit of Elaeocarpus prelancaefolius Bera et al.(2004) from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/IB7/5/F1)(Scale bar=1 cm);(h)Zig-zag type leaf mining on the fossil leaf of Terminalia panandhroensis(Lakhanpal and Guleria)Khan et al.(2014b)from Middle Siwalik sediments of Southeastern Himalaya(CUH/PPL/B/54)(Scale bar=1 cm);(i)Scanning electron micrographs of Gmelina siwalika Khan et al.(2018a)from Upper Siwalik sediments of Southeastern Himalaya(CUH/PPL/C3/44)-transverse section of the secondary xylem showing vessel with a prominent tylosis(Scale bar=50 μm);(j) Dysoxylum raptiensis Khan et al.(2015a) from Upper Siwalik sediments of Southeastern Himalaya (CUH/PPL/IB7/17) - adaxial cuticle with characteristic frass-trail (Scale bar = 10 μm).
The nearest living relatives (NLR) method extrapolates the known climatic requirements of modern taxa back to comparable and related taxa in the past and presupposes that fossil plants and their modern relatives share similar physiological requirements for climate (Mosbrugger and Utescher, 1997).The megafossil assemblages recovered from the Siwalik sediments of the eastern Himalayas have yielded mainly angiosperm taxa (Tables S1-S3)that can be used effectively to interpret palaeoclimate and palaeoflora because the nearest living relatives of these fossil angiosperm taxa are known with high confidence.On the basis of NLRs,the Siwalik floral assemblages of the eastern Himalayas consist of three major forest elements: evergreen (58.60%), deciduous(26.82%),and others(14.39%)(Fig.6a-d).In the eastern Himalayan Lower Siwalik assemblage 63.73% of the taxa are evergreen, and deciduous elements make up just 23.07%of the taxa(Fig.6a).In the Middle Siwalik assemblage, 57.74% of the taxa are evergreen and 25.35%of taxa are deciduous(Fig.6b).In the Upper Siwalik 54.34%and 32.60%are evergreen and deciduous respectively(Fig.6c).With respect to the present-day distribution pattern of NLR taxa this suggests that wet evergreen forests persisted throughout the period of deposition (Tables S1-3; Fig.6d).The predominance of evergreen elements in the assemblage along with pteridophytes(ferns)indicates the prevalence of a tropical,warm,humid climate with abundant rainfall in contrast to the relatively dry present-day climate in the area.An increase in deciduous elements is evident towards the close of the Middle Siwalik and the beginning of the Upper Siwalik(Fig.6e).This change in the vegetation pattern must reflect a climatic change between the lower part (Miocene) and upper part of Siwalik (Plio-Pleistocene) deposition, and seems to indicate an increase in rainfall seasonality.We reconstruct the paleovegetation to better understand the general evolutionary history of floristic patterns in eastern Himalaya during Siwalik sedimentation (Fig.7).

Table 2 (continued)
7.Palaeoclimates reflected by the fossil floras
Investigations using different qualitative(NLR,cuticular studies)and quantitative(Co-existence Approach and CLAMP)proxies have revealed the history of past climate,and in particular the evolution of Indian summer monsoon(ISM),during the Siwalik sediments of the eastern Himalayas(Khan and Bera,2014a,b;Khan et al.,2014a,2015, 2019a, b; Prasad et al., 2015; Srivastava et al., 2021; Bhatia et al., 2022).
7.1.A qualitative NLR approach

Fig.6.(a) Diagrammatic representation of different types of forest elements of the lower Siwalik flora of the eastern Himalayas (E = Evergreen, D = Deciduous, O = Others); (b)Diagrammatic representation of different types of forest elements of middle Siwalik flora of the eastern Himalayas(E=Evergreen, D=Deciduous, O=Others); (c)Diagrammatic representation of different types of forest elements of upper Siwalik flora of the eastern Himalayas(E=Evergreen,D=Deciduous,O=Others);(d)Diagrammatic representation of different types of forest elements of entire Siwalik flora of the eastern Himalayas(E=Evergreen,D=Deciduous,O=Others);(e)Schematic sketch of the floristic pattern changes throughout the Siwalik sediments in the eastern Himalayas.
The principal basis of any study of the past is that known as‘uniformitarianism’.This, principle, often summarized as ‘the present is the key to the past’, implies that the physical and biological processes that operate in today's environment, as well as vegetation, must have functioned in a similar way in the past(Thanukos, 2012).In the case of the NLR approaches used to reconstruct past climates, this extrapolates the known climatic requirements of modern taxa to presumed ancestral taxa in the past.Of the plant fossils recovered from the eastern Himalayas that have nearest living relatives, several still exist in the area today.This suggests some degree of climatic similarity between the past and now and the persistence of a tropical warm and humid climate.
7.2.The coexistent approach (CoA)
CoA,developed by Mosbrugger and Utescher(1997),is based on the concept that the climatic requirements of fossil species are similar to those of their NLRs and reconstructs the paleoclimate parameters for a given fossil flora using climatic intervals in which all the NLRs of the fossil flora could coexist coexist(Mosbrugger and Utescher,1997;Mosbrugger,1999;Utescher et al.,2014).This is an improvement from many previous NLR analyses that have tended to choose single, or at best, just a few taxa for the analysis.By adopting a whole-population approach, outliers due to misidentification or evolutionary innovation can be isolated and removed from the analysis, thus, improving accuracy and precision.Prasad et al.(2015) first reconstructed the eastern Siwalik palaeoclimate by applying this quantitative method to the middle Siwalik flora of Darjeeling sub-Himalaya.They estimated different climatic variables, such as mean annual temperature (MAT), warmest month mean temperature (WMMT), coldest month mean temperature(CMMT), and mean annual precipitation (MAP) as 22-26.5°C,17.8-20°C,25-30°C,and 2650-3200 mm,respectively.However,their methodology differed from that codified by Mosbrugger and Utescher (1997) and Utescher et al.(2014) and did not specify the origins of their plant distribution data.Srivastava et al.(2021)subsequently reconstructed the climate of the Upper Siwalik strata of Southeastern Himalaya quantitatively, based on the more usual form of CoA specified by Mosbrugger and Utescher (1997)and Utescher et al.(2014), and reported that during the late Pliocene-early Pleistocene, the temperature seasonality between warm (27-28.1°C) and cold months (22-23.6°C) was less pronounced compared with present-day warm (27-27.7°C) and cold(14.8-15.4°C) month conditions.The reconstructed rainfall data indicated a monsoonal type of climate having a strong wet/dry seasonality during the deposition of the Upper Siwalik sediments.Recently,Bhatia et al.(2022)also applied this methodology to two Siwalik floras recovered from the Lower (middle Miocene) and Middle (late Miocene-Pliocene) Siwalik successions of Darjeeling in the eastern Himalayas.The reconstructed climate data suggested a decrease in both winter temperature and precipitation during the wettest months, and thus an overall drying, from the Lower to Middle part of the Siwalik succession.

Fig.7.Reconstruction of the palaeovegetation during Siwalik sedimentation of the eastern Himalayas.
7.3.The CLAMP approach
The principal leaf-based palaeoclimate proxy for assessing a range of climate variables is known as CLAMP (Climate Leaf Analysis Multivariate Program; http://clamp.ibcas.ac.cn) (Wolfe, 1993;Kovach and Spicer,1996;Yang et al.,2011,2015).CLAMP utilizes the universal relationships that exist between leaf form in woody dicotyledonous plants and an array of climate variables.On a global scale,aggregate leaf form in a stand of vegetation is more strongly determined by climate than by taxonomic composition(Yang et al.,2015).Using a multivariate statistical engine,CLAMP decodes these relationships and, by scoring fossil leaf traits the same way as for living vegetation growing under known climatic regimes,estimates past conditions (http://clamp.ibcas.ac.cn).Five fossil floras (one lower Siwalik mid-Miocene, one middle Siwalik Pliocene, and one upper Siwalik Plio-Pleistocene flora of Southeastern Himalaya;one lower Siwalik mid-Miocene flora of Darjeeling and one latest Miocene-Pliocene middle Siwalik Group of Bhutan sub-Himalaya)ranging in age from the mid-Miocene to the early Pleistocene from the eastern Siwalik near Bhutan, Darjeeling and in Southeastern Himalaya were also subjected to a CLAMP analysis using a calibration data set that includes sites from India, southern China,and Thailand and gridded climate data (Khan et al., 2014a, 2019b)(Tables S7-S11).CLAMP climate retrodictions derived from the PhysgAsia2 calibration for all the fossil sites of the eastern Himalayas are given in Tables 3-5.
The results of these analyses are also consistent with published quantitative climate data (Khan et al., 2014a, 2019b) using CLAMP analysis (Wolfe,1993; Teodoridis et al., 2011; Yang et al., 2011) on fossil leaf morphotypes (i.e., not assigned taxonomic affiliation)from the eastern Himalayas.Two lower Siwalik mid-Miocene floras of Darjeeling and Southeastern Himalaya yielded almost the same values suggesting mean annual temperatures (MATs) of 25.4 and 25.3±2.8°C(all uncertainties±2 sigma)with warm month mean temperatures(WMMTs)of 28.4 and 27.8±3.39°C and cold month mean temperatures(CMMTs) of 17.9 and 21.3 ± 4°C.Precipitation estimates have high uncertainties but suggest a weak monsoon with growing season precipitations of 181 ± 91 cm for Bhutan,242 ± 92 cm for Darjeeling, and 174 ± 92 cm for Southeastern Himalaya.Leaves from the middle Siwalik (Pliocene) sediments of Southeastern Himalaya indicate a lowering of the MAT to 23.7°C,which appears to be largely a function of cooler winter months(CMMT 16.9°C).Southeastern Himalaya's early Pleistocene temperatures and rainfall were similar to those of the mid-Miocene.Khan et al.(2019b) compared palaeoclimate estimates of the latest Miocene-Pliocene Siwalik(ca.6 to 3.8 Ma)flora of Bhutan with those of Siwalik floras from the Miocene-Pleistocene of Southeastern Himalaya and the Miocene Siwalik flora of Darjeeling.Because all the Siwalik floras of the eastern Himalayas spanning the mid-Miocene to Pleistocene yield almost the same values, they suggested that overall, the eastern Himalayan Siwalik climate appears to have been remarkably uniform over the past 15 million years.The MAT result of the Bhutan Siwalik palaeoflora differs by just 0.6°C from the Southeastern Himalaya, and 1.2°C from the Darjeeling palaeoflora.For all Siwalik fossil assemblages, WMMTs,CMMTs, LGSs (length of the growing season), RH (mean annual relative humidity), and SH (mean annual specific humidity) aresimilar and consistent (WMMTs around 28°C, CMMTs around 18°C, LGSs around 12 months, RHs around 80% and SHs around 14 g/kg).Hence, palaeoclimate estimates of the Southeastern Himalaya, Darjeeling, and Bhutan Siwalik flora provide valuable insights into monsoon climatic evolution throughout the eastern Himalayan Siwalik belt during late Cenozoic time and indicate that the Siwalik floras experienced a persistent monsoonal tropical warm humid climate.Changes in the Monsoon index suggest that in both the Bhutan and Southeastern sub-Himalaya,there has been little change in the intensity of the monsoon since mid-Miocene time, while further west in the Darjeeling area precipitation seasonality has increased since the mid-Miocene.
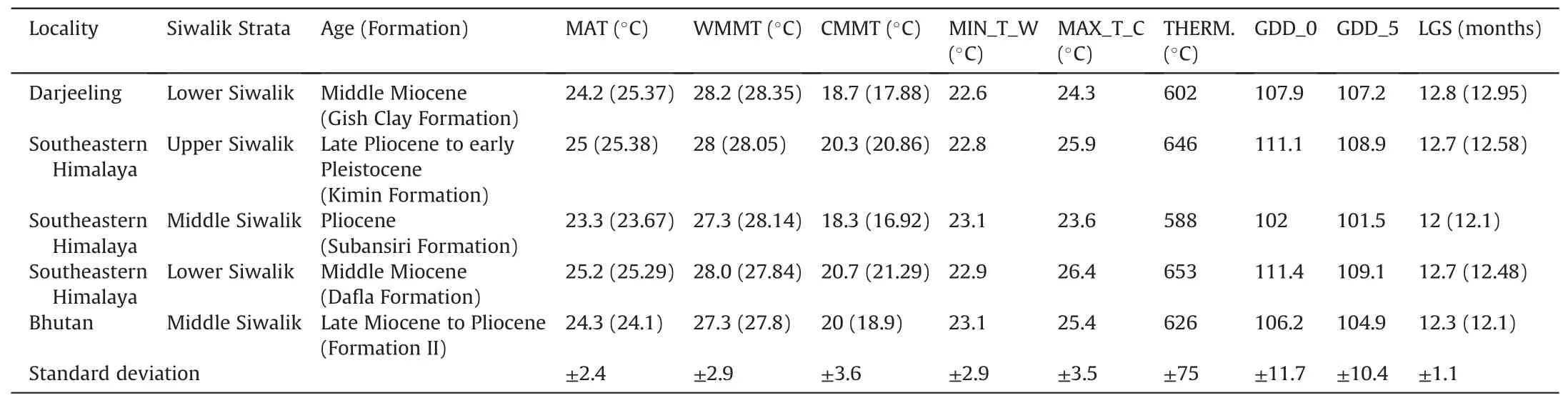
Table 3 Summary of temperature-related CLAMP-derived metrics for Siwalik leaf assemblages from the eastern Himalayas.Values obtained by a CLAMP calibration based on PhysgAsia2 trait scores and WorldClim2 climate data as well as HiResGridMetAsia2 (in parentheses)gridded climate data.

Table 4 Summary of precipitation and moist enthalpy CLAMP-derived metrics for Siwalik leaf assemblages from the eastern Himalayas.Values obtained by a CLAMP calibration based on PhysgAsia2 trait scores and WorldClim2 climate data as well as HiResGridMetAsia2 gridded climate data, in parentheses.

Table 5 Summary of humidity CLAMP-derived metrics for Siwalik leaf assemblages from the eastern Himalayas.Values obtained by a CLAMP calibration based on PhysgAsia2 trait scores and WorldClim2 climate data as well as HiResGridMetAsia2 (in parentheses) gridded climate data.
7.3.1.New insights into the thermal and hydrological regime of the eastern Himalayan Siwalik
Here, we re-analyze the five well-documented fossil leaf assemblages from across the eastern Himalayas spanning Siwalik time (middle Miocene to early Pleistocene).All have been previously analyzed for the eleven standard CLAMP climate variables(mean annual temperature - MAT; warm month mean temperature-WMMT;cold month mean temperature-CMMT;length of the growing season - LGS; growing season precipitation - GSP;mean monthly growing season precipitation - MMGSP; precipitation during the three consecutive wettest months - 3WET; precipitation during the three consecutive driest months - 3DRY;mean annual relative humidity - RH.ANN; mean annual specific humidity - SH.ANN; and mean annual moist enthalpy - ENTH),calibrated using modern gridded climate data at 10'spatial resolution (HiResGridMetAsia2) and physiognomic PhysgAsia2 calibration(Table S12).Here,fossil leaf assemblages are subjected to a CLAMP analysis using a new high spatial resolution 30" (~1 km2)WorldClim2 gridded climate data (Fick and Hijmans 2017; http://worldclim.org/version2) (Table S13) with 15 new climate variables (Tables 2-4; Figs.8 and S1-S4).However, we use the same modern vegetation trait scores as used previously (PhysgAsia2)(Table S12).This calibration data set interpolates average meteorological observations between 1970 and 2000 onto a spatial grid approximating 1 km2.CLAMP climate retrodictions for all the fossil sites of the eastern Himalayas are given in Tables 3-5.
One advantage of using WorldClim2 for calibration is that numerous environmental variables have been mapped onto the same grid, so for CLAMP, the range of environmental signals decoded from leaf form can be extended.The new temperaturerelated environmental variables that correlate strongly with leaf form are (1) the compensated thermicity index -THERM (sum of mean annual temperature, minimum temperature of the coldest month,the maximum temperature of the coldest month,×10,with compensations for better global comparability),(2)growing degree days above 0°C - GDD_0 (sum of mean monthly temperature for months with mean temperature >0°C multiplied by the number of days this occurs), (3) growing degree days above 5°C - GDD_5(Sum of mean monthly temperature for months with mean temperature >5°C multiplied by the number of days this occurs), (4)minimum temperature of the warmest month-MIN_T_W(lowest daily temperature during the warmest month) and (5) maximum temperature of the coldest month - MAX_T_C (warmest daily temperature during the coldest month).The new humidity-related variables are(6) mean annual vapour pressure deficit - VPD.ANN,(7) mean summer vapour pressure deficit - VPD.SUM (average vapour pressure deficit during the three summer months), (8)mean winter vapour pressure deficit - VPD.WIN (average vapour pressure deficit during the three winter months), (9) mean spring vapour pressure deficit-VPD.SPR(average vapour pressure deficit during the three spring months), (10) mean autumn vapour pressure deficit-VPD.AUT(average vapour pressure deficit during the three autumn months), (11) mean annual potential evapotranspiration - PET.ANN (the ability of the atmosphere to remove water through evapotranspiration, given unlimited water supply- no limits on plant water supply- averaged over the year), (12) mean monthly potential evapotranspiration during the warmest quarter-PET.WARM and(13)mean monthly potential evapotranspiration during the coldest quarter - PET.COLD.
Tables 3-5 present results obtained for the fossil assemblages using the new WorldClim2 CLAMP calibration, as well as (for comparison)previously obtained results(in parentheses)that used low spatial resolution HiResGridMetAsia2CLAMP calibration.Figs.8 and S1-S4 illustrate the CLAMP regression models for each of the climate variables to show not only the relative position on the regression of the Siwalik fossil locations but also the scatter of the modern training data and thus the precision of the CLAMP predictions.As used in earlier CLAMP analyses, all regression models are derived from the leaf physiognomy/climate relationships in four-dimensional space (Spicer and Herman, 2010).CLAMP scoresheets for all eastern Himalayan Siwalik fossil assemblages are given in the Tables S7-S11.The new WorldClim2-based climate training set(WorldClim2_3br) and the accompanying modern leaf physiognomic (PhysgAsia2) data files are given in the Tables S12 and S13.
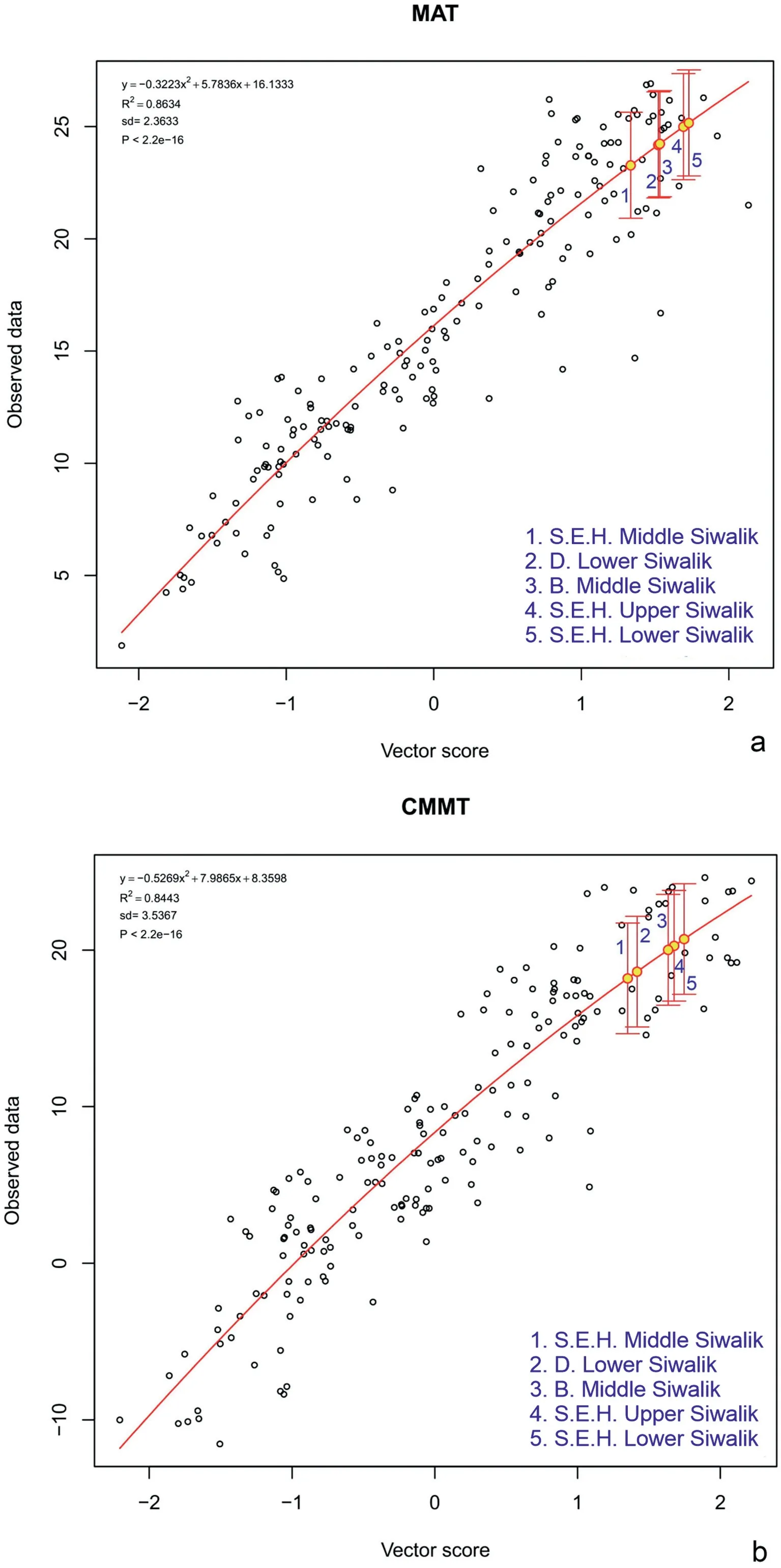
Fig.8.CLAMP WorldClim2 regression models for(a)mean annual temperature(MAT)and(b)cold month mean temperature(CMMT).The position of the eastern Himalayan fossil flora along the second-order polynomial regression relating the MAT and CMMT vector scores for modern vegetation against the observed MATs and CMMTs for those sites is shown as a red-rimmed circle with a yellow center, with uncertainty bars (1 s.d.) reflecting the scatter of the residuals about the regression line.The vector score represents the relative position of the sites, modern and fossil, along with a vector representing the primary trend of the climate variable in axes 1-4 space.See the CLAMP website (http://clamp.ibcas.ac.cn) for details.
The new calibration and range of climate variables allow us to explore new insights into the hydrological regime.We examine not only precipitation but humidity in terms of specific humidity(SH),relative humidity(RH),vapour pressure deficit(VPD),and potential evapotranspiration (PET).Both VPD and PET are investigated in respect of annual average values and seasonal variations.
Using leaf form (physiognomy) we reconstruct middle Miocene-early Pleistocene thermal and hydrological regimes at five locations in the eastern Himalayas.The new high spatial resolution (~1 km) WorldClim2 calibration yields result similar to previous analyses,but also provides more detailed insights into the hydrological regime through the return of annual and seasonal vapour pressure deficit (VPD), potential evapotranspiration (PET)estimates, as well as new thermal overviews through measures of thermicity and growing degree days.The new results confirm the overall warmth of the region.Palaeo-rainfall estimates have large uncertainties due to moisture not being limiting in the context of the Siwalik assemblages and because fossils are usually preserved in water-lain deposits, suggesting the parent plants were growing in or near year-round wet soils.The new measures of VPD and PET show the persistent high humidity to which the leaves were exposed and adapted,but with notably lower humidity during the summers at all the eastern Himalayan locations.
7.4.Cuticular approach
No proxy is perfect, so a multiproxy approach is always desirable.The examination of fossil leaf cuticles found on compressed leaves can also afford an estimation of past climate.Several cuticular characters indicate a warm, humid tropical climate with nonlimiting rainfall, including thin cuticles, undulate to sinuous epidermal lateral walls, non-papillate or smooth leaf external surfaces, few epidermal hairs, unspecialized stomata, and subsidiary cells, all of which are commonly found in the Siwalik assemblages(Khan et al., 2015a).The hypostomatic nature of many Siwalik stomata also reflects heavy precipitation, humidity, and shade.Cuticular micro-morphological features have also helped to confirm the identification of some leaf compressions to the species level,and are clearly indicative of mesophytic ecological conditions that reflect a tropical climate with high precipitation (Khan et al.,2015a, 2016, 2017a, b).
Some workers(Mitra and Banerjee,2000;Mitra et al.,2002;Das et al.,2007;Mandal et al.,2009,2011;Vishnu et al.,2017,2019;Bera et al., 2018, 2019, 2022a, b; Khan et al., 2018b, 2019c) reported insitu occurrences on leaf megafossils of characteristic epiphyllous fungi such as Meliolinites (fossil Meliolaceae) (comparable to the modern genus Meliola Fr.), Phomites (comparable to the modern genus Phoma Sacc.), Palaeocercospora (comparable to the modern genus Cercospora Fresen.ex Fuckel), Palaeocolletotrichum (comparable to the modern genus Colletotrichum Corda), Palaeoasterina(comparable to the modern genus Asterina L´ev.) and Vizellopsidites(comparable to modern genus Vizella Sacc.) on the cuticular surfaces of fossilized leaf cuticle fragments of the different angiosperm taxa recovered from the Siwalik sediments (middle Miocene to early Pleistocene) of Darjeeling, Bhutan, and Southeastern Himalaya(Table S6).They described fossil fungi on the basis of vegetative and reproductive structures.The Siwalik host leaves harboring the fossil fungi so far identified are Shorea, Dipterocarpus (Dipterocarpaceae),Breonia A.Rich.ex D.C.(Rubiaceae),Dysoxylum Blume(Meliaceae), Combretum (Combretaceae), Xylopia L.(Annonaceae),Amherstia Wall.(Fabaceae), Actinodaphne Nees, Lindera Thunb.,Persea(Lauraceae),Macaranga Thouars(Euphorbiaceae),Lauraceae,and Poaceae.Based on earlier records, it is also evident that Lauraceae has been a common host for meliolaceous fungi since the early Cenozoic (Khan et al., 2019c).The reported appreciable numbers of foliicolous fungal remains indicate the prevalence of a warm, humid, climate favored by the high rate of precipitation in the eastern Himalayas during the Plio-Pleistocene (Mitra and Banerjee, 2000; Mitra et al., 2002; Das et al., 2007; Mandal et al.,2009, 2011; Vishnu et al., 2017, 2019; Bera et al., 2018, 2019,2022a, b; Khan et al., 2018b, 2019c).These climatic data are also consistent with published climatic data obtained from the study of the macroscopic plant remains using qualitative and quantitative methods.Thus, for the eastern Himalayan Siwalik, all approaches(CLAMP, NLR, CoA, and cuticle) give broadly similar palaeoclimate outcomes.The in-situ evidence of epiphyllous fungal remains on host leaf cuticles also indicate the possible existence of a hostectoparasite relationship in the ancient warm and humid tropical evergreen forest of this area during Siwalik sedimentation(Vishnu et al.,2017,2019;Bera et al.,2018,2019,2022a,b;Khan et al.,2018b,2019c).
8.Comparisons
8.1.Comparisons with other Siwalik floras
To reveal the degree of resemblance to other Siwalik floras(western and central), we make the following comparisons.
8.1.1.Western Siwalik flora
This includes the floras of Jammu and Kashmir, Uttarakhand,and Himachal Pradesh and comprises a large number of fossil woods and leaves (Sahni,1964a, b, Lakhanpal,1965,1967; Varma,1968; Lakhanpal and Awasthi, 1992; Prasad, 1994a, 2006; Prasad et al.,1997; Shashi et al., 2006, 2008; Srivastava et al., 2015).The NLRs of common fossil taxa are Millettia, Ziziphus Mill., Pongamia,
Dalbergia, Diospyros, Fissistigma, Bambusa Schreb., Dipterocarpus,Lagerstroemia, Marantochloa Brongn.ex Gris, Calophyllum, Shorea,Gynocardia, Grewia, Cynometra, Persea, Sterculia, Mallotus, Terminalia, and Hopea Roxb.This indicates that these taxa were widely distributed in both eastern and western Siwalik strata and flourished under a generally equitable climate, at least in terms of temperature.
8.1.2.Central (Nepal) Siwalik flora
Plant megafossils(mainly fossil leaves)are known from various localities in Nepal such as Koilabas,Arung Khola,Surai Khola,Tinau Khola,Babai and Surkhet Valleys,Mahendra Highway,Arjun Khola,and Sindhuli (Prakash and Prasad, 1984; Prasad, 1990, 1994b;Prasad et al.,1997,1999;Tripathi et al.,2002;Dwivedi et al.,2006;Prasad and Dwivedi, 2007).Comparison of the eastern Himalaya fossil floras with those of the central Siwalik fossil flora assemblages shows that most NLRs of the fossil genera Mesua L., Mangifera L.,Bouea Meisn.,Garcinia,Albizia,Cassia L.,Millettia,Ziziphus,Pongamia,Dalbergia,Diospyros,Fissistigma,Bambusa,Dipterocarpus,Lagerstroemia, Marantochloa, Calophyllum, Shorea, Gynocardia, Grewia, Cynometra, Persea, Sterculia, Melilotus, Cinnamomum Schaeff.,Mitrephora Hook.f.&Thomson,Hopea,Polyalthia Blume,Uvaria L.,Sabia Colebr.,Miliusa Lesch.ex A.D.C.,Swintonia Griff.,Euphorbia L.,Entada, Combretum, Dillenia, Randia L., and Flacourtia Comm.ex L'H´er.are common to both regions.
8.2.Comparisons among Siwalik CLAMP data
8.2.1.Methodology
The CLAMP methodology, its limitations, and its evolution are detailed in Spicer et al.(2021),but in summary,CLAMP reconstructs past climate based on an array of macroscopic leaf traits preserved in leaf megafossils.Organic remains are not required and CLAMP can be applied to mere leaf impressions provided that the leaves retain sufficient trait data across a minimum of 20 taxa or morphotypes in any one fossil assemblage.Also, leaf identification is not required,only an ability to distinguish one taxon(morphotype)from another.Morphotype partitioning is based not only on the CLAMP traits (31 trait states encompassing leaf lobing, margin features size,apex,base forms,and overall shape,but also venation and other taxonomically useful features.
To calibrate CLAMP, a database of trait spectra from modern vegetation stands growing under a wide range of known climate conditions provides a multidimensional framework for identifying correlations between leaf trait combinations and individual climate variables such as temperature and moisture metrics.This physiognomic data set is accompanied by a suite of climate data derived in most cases from gridded observations.There are several such modern gridded data sets available at different spatial resolutions(e.g., New et al.,1999, 2002; Harris et al., 2014; Fick and Hijmans,2017) and each are slightly different, largely as a function of interpolation artifacts and different observation periods, and so return slightly different retrodictions of past climate.This is shown in Tables 3 and 4 where WorldClim2 calibration results are compared with those based on New et al.(2002), shown in parentheses.These climate calibration anomalies apply to any climate proxy,so it is important when comparing proxy-reconstructed past climates that uncertainties introduced by using different types of climate calibration data are fully appreciated.Here we use a common proxy CLAMP calibration combining the PhysgAsia2 leaf trait data set (Spicer et al., 2021) with the WorldClim2 climate data gridded at ~1 km resolution (Fick and Hijmans, 2017).The trait/climate relationships are decoded using the multivariate statistical engine known as Canonical Correspondence Analysis (ter Braak,1986; http://clamp.ibcas.ac.cn).
Recently, fossil leaf assemblages from the lower (middle Miocene) and middle (late Miocene-Pliocene) Siwalik sediments exposed in Nepal were subjected to a CLAMP analysis by Bhatia et al.(2021) using the new high spatial resolution (~1 km2)WorldClim2 gridded climate data and PhysgAsia2 calibration.Their analysis indicates a mean annual temperature (MAT) of 22.2 ± 2.3°C and 24.7 ± 2.3°C for the Lower Siwalik and Middle Siwalik assemblages,respectively.Cold month mean temperatures(CMMTs) were 14.7 and 19 ± 3.5°C and warm month mean temperatures (WMMTs) were 28.3 and 28.5 ± 3°C for the Lower and Middle Siwalik assemblages,respectively,showing warming of the cold months being mostly responsible for a slight increase in the MAT over that time interval.Here, we compare palaeoclimate estimates of the middle Miocene to Plio-Pleistocene Siwalik flora from the eastern Himalayas with those of previously investigated Siwalik middle Miocene-Pliocene floras of Nepal.
Tables 3-5 summarize the CLAMP results for the eastern Himalaya Siwaliks.Table 3 focuses on temperature-related metrics while Table 4 provides an overview of rainfall-related metrics and Table 5 presents humidity-related metrics.All middle Miocene(Lower Siwalik of Southeastern Himalaya and Darjeeling) thermal metrics are identical within 1 sigma uncertainty,showing frost-free year-round growth in a climate where the CMMT is >18°C and thus‘tropical’.The CMMT of the Lower Siwalik of Nepal is somewhat lower at 14°C and just on the margins of the 1 sigma uncertainty difference from the eastern Himalaya value reported here.This marginal difference is also evident in the cooler (21°C for Nepal versus 24.3 and 26.4°C for Darjeeling and Southeastern Himalaya,respectively) maximum temperature of the coldest month metric,but again it is debatable whether these differences can be regarded as real.For the Middle Siwalik assemblages (late Miocene to Pliocene of Southeastern Himalaya and Bhutan) a similar pattern is evident with all thermal metrics showing no discernible differences between assemblages (Table 5).These values are also more or less identical to those from the Middle Siwalik of Nepal reported by Bhatia et al.(2021).There is only one Upper Siwalik(late Pliocene to Pleistocene)assemblage reported here from the eastern Himalayas and that is from the Southeastern Himalaya section.This also shows thermal metrics apparently unchanged from those of earlier times.
Regarding rainfall-related metrics (Table 4), the Lower Siwalik assemblages of Darjeeling and Southeastern Himalaya are identical within uncertainty except for precipitation during the three consecutive driest months, which indicates Darjeeling was markedly wetter than Southeastern Himalaya.This difference is independent of which meteorological data set is used for calibration.The Middle Siwalik sites similarly are indistinguishable from one another,this time across all moisture metrics.The Darjeeling Lower Siwalik assemblage appears to be notably wetter than the other sites, particularly in respect of precipitation during the three consecutive driest months, but apart from that, all assemblages appear very similar in terms of their reconstructed climate metrics.
It is often assumed that rainfall is an important environmental constraint on plant growth, but this need not be the case.What is critical is soil moisture combined with transpirational stresses imposed by atmospheric humidity and wind strength.In situations where leaf fossils are preserved (i.e., near water bodies), soil moisture reflects the proximity to that water body,not necessarily local rainfall, and because of this, the soil moisture in these situations is rarely limiting.It follows that a more useful measure of atmospheric conditions can be found in the way that leaf traits code for humidity metrics, especially vapour pressure deficit (VPD) and potential evapotranspiration(PET)(Spicer et al.,2019,2021).VPD is a measure of the ease by which a plant can lose moisture to the atmosphere, with low VPDs found when the air is near saturation and there is strong resistance to transpiration, whereas at high VPDs there is no atmospheric constraint on transpirational water loss from the plant.Unlike relative humidity(RH),VPD has a nearly straight-line relationship to the rate of evapotranspiration, and plant distribution (Huffaker, 1942) and leaf physiognomy (Spicer et al., 2019) are more reflective of VPD than RH.PET is similar but is a measure of the ability of the atmosphere to remove water through evapotranspirational processes provided the water supply to the roots is not limiting.Unlike with VPD,atmospheric dynamics(convection and wind) play a role in determining PET.
Table 5 suggests very similar values of humidity metrics among the different assemblages through time.That is to say,there are no discernible step changes in climate features from the mid-Miocene to the Pleistocene.However, there are some important seasonal differences in VPD and PET metrics consistent across all assemblages, with summer being consistently more humid than spring,as with the modern SAM, but Darjeeling shows less seasonal variation in VPD than at Bhutan or Southeastern Himalaya.This reflects the relatively wet dry season in Darjeeling as indicated by rainfall metrics (Table 4).Similarly, the cold month (winter) PET value is more than 1 standard deviation lower than the other cold month PET values, suggesting a markedly wetter dry season.
Because all the Siwalik floras of the eastern Himalayas and Central Himalayas yield almost the same values, we suggest that overall,the eastern and central Himalayan Siwalik climate appears to have remained remarkably uniform from the mid-Miocene to Pleistocene.However, while the modern temperature regime for the eastern Himalayan Siwaliks exhibits cooler winters than evident from the fossil data, the overall temperature regime is similar over time.The most marked differences are in the precipitation regime, with the modern being wetter with a greater seasonality in rainfall (wetter wet seasons and drier dry seasons)(Table S14).However,because uncertainties are also quite large for these metrics, the differences between the past and present may not be genuinely significant.It is unlikely that this consistency over time is representative of the whole SAM region, but it does have important implications for ecosystem evolution in the eastern Himalayas at low elevations, as discussed below.
9.Plant-arthropod associations from Siwalik forests
Fossil leaf impressions and compressions from the Siwalik sedimentary strata of the eastern Himalayas provide evidence of a variety of plant-insect interactions that have operated throughout the evolution of monsoon-influenced forests since middle Miocene times (Khan et al., 2014b, 2015b).Five functional feeding groups(FFGs) were identified in this study, namely leaf mining, hole feeding, skeletonizing, galling, and margin feeding.Furthermore,these morphotraces are similar to those found in leaves of extant plant species such as Millettia,Canarium L.,Glochidion J.R.Forst.&G.Forst., Callicarpa L., Chonemorpha G.Don, Actinodaphne, Persea,Woodfordia Salisb., Shorea, Artocarpus J.R.Forst.& G.Forst., Albizia,Lagerstroemia and others, suggesting similar interactions have existed in the eastern Himalaya region for at least 15 million years.On the basis of comparison with extant taxa, possible leaf feeders could have belonged to the insect orders Orthoptera, Coleoptera,Lepidoptera, and Diptera, and those plant-arthropod relationships were established by the mid-Miocene and continue to the present,shaping both the present-day flora and fauna.Khan et al.(2014b,2015b) also compared insect herbivory evident in the Darjeeling Lower Siwalik flora to that of the similarly aged Southeastern Lower Siwalik flora,as well as two younger floras from that area and noted a similar range of FFGs and damage types among all four fossil floras of the eastern Himalayas.They concluded that compared to biotic factors climate had little influence on determining the evolution of plant-insect interactions in the eastern Himalayan region.
10.Phytogeographic patterns
The Siwalik plant fossils date from the late Neogene time(middle Miocene to early Pleistocene), and close relatives of the fossil forms still exist in the tropical forests of India and Southeast Asia today.This allows direct tracking of changes in phytogeographic distributions of these taxa over time.The present-day distributions of NLRs of 219 fossil taxa recovered from the Siwalik sediments(Mio-Pleistocene)of the eastern Himalayas indicate that today they grow in a variety of locations all over India and other adjoining countries (Fig.9).In India, they are distributed in the northeastern(14.75%)and southern regions(4.35%).In this Siwalik assemblage, the NLRs of 49.04% taxa (Shorea roxburghii G.Don,
Uvaria hirsuta A.St.Hil., Dipterocarpus tuberculatus Roxb., D.turbinatus Roxb., Ventilago calyculata Tul., Syzygium cuminii (Gamble)Tenjarla & Kashyapa, Homonoia riparia Lour., Xanthophyllum flavescens Roxb., Mitrephora maingayi Hook.f.& Thomson, Hopea wightiana Roxb., Bauhinia accrescens Killip & J.F.Macbr., Randia wallichii Hook.f., R.densiflora Lou., Fissistigma bicolor Merr.,B.malabaricum D.C., Sterculia parviflora Roxb., Buchanania sessilifolia Blume,Cynometra iripa Kostel.,C.polyanthum Wall.ex Choisy,Glochidion zeylanicum (Gaertn.) A.Juss., P.pinnata (L.) Merr.occur both in India and the Malaya Peninsula and 22.7% taxa (Pterospermum yunnanense H.H.Hsue, Parashorea buchananii (C.E.C.Fisch.) Symington, Flacourtia inermis Wall., Fordia albiflora (Prain)Dasuki & Schot, Paranephelium macrophyllum King, Diospyros argentea Griff.) are found to generally grow in Malaya.This clearly indicates a free exchange of floral elements during the late Neogene across Southeastern Asia (Fig.10).
The late Neogene floral assemblages covering the eastern Himalayas and Southeast Asian countries (Sumatra, Borneo,Indonesia, Vietnam, Malaysia, and Java) not only reflect great diversity with respect to the angiosperm families and variety of taxa but also uniformity and similarity in floristic composition.Thus,the late Neogene may be regarded as the time of maximum proliferation and diversification of tropical vegetation, particularly in the evergreen forests in Southeast Asia (Bande and Prakash, 1986;Awasthi and Mehrotra,1990,1997;Guleria,1992).This floral spread was not unidirectional but suggests cross-migration between the regions as some Southeast Asian taxa must have migrated from the Indian landmass due to the widespread distribution of similar edaphic and climatic factors.Evidence of many southeast Asian elements recovered from the Siwalik exposures of the eastern Himalayas suggest migration into the Indian subcontinent and intermixing and merging with the existing eastern Himalayan flora before expanding further westwards along the entire Himalayan lowlands.Hence, the late Neogene period may be considered an important time in the evolution and speciation of flowering plants in India and Southeast Asia through the introduction and proliferation of new floral elements and the opportunity for subsequent interbreeding.
11.Disappearance of some plant taxa
Gradual changes in climate and monsoon amplification across Asia during the Neogene (Su et al., 2013; Tang et al., 2013, 2015)have been attributed to a general global cooling(Zachos et al.,2001,2008),closure of the Tethys,loss of littoral environments along the Himalayan front(Lakhanpal,1970)and rapid uplift of the Himalayas(Chatterjee and Scotese,1999;Boos and Kuang,2010;Molnar et al.,2010;Ding et al.,2017;Farnsworth et al.,2019).These factors are all likely to have played an important role in altering and diversifying floral patterns (Morley, 2000; Huggett 2004; Mosbrugger et al.,2005).Some plant taxa were able to adapt to the changed circumstances or already had wide environmental tolerances.These have continued to flourish to the present-day in the areas surrounding the fossil localities, whereas others that could not adapt to the new environment either suffered local extinction or shifted to suitable areas with more favourable eco-climatic conditions.

Fig.9.(a) Diagrammatic representation of Siwalik flora of the eastern Himalayas represented in the present-day flora of different geographical regions(NE=Northeast India; SI = South India; IM = India and Malaya Peninsula; M = Malaya; C = Cosmopolitan; O = Others); (b) Diagrammatic representation of the Siwalik flora of the eastern Himalayas in three different categories (EL = Extant local taxa; ELE = Extant but locally extinct taxa; ERE = Extant but regionally extinct taxa).
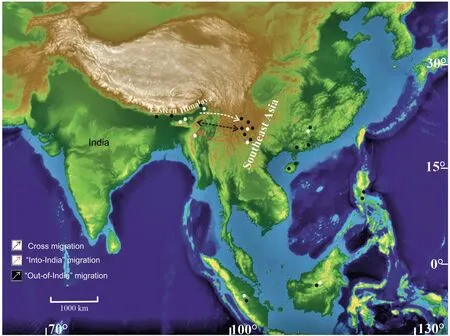
Fig.10.Map showing hypothetical migratory routes of Siwalik and Southeast Asian tropical elements.
Based on the present-day distribution of NLRs of the eastern Himalayan Siwalik plant fossil species, we categorize the Siwalik species into three main groups:extant local species(65%),extant but locally extinct species(12%),and extant but regionally extinct species (23%) (Fig.9b).Extant local species have their NLRs (for example,Guatteria australis A.St.Hil.,D.tuberculatus Roxb.,D.turbinatus C.F.Gaertn., S.cuminii (Gamble) Tenjarla & Kashyapa, Tarennoidea wallichii(Hook.f.)Tirveng.&Sastre,F.bicolor Merr.,B.ceiba
L.,S.parviflora Roxb.,C.iripa Kostel.,C.polyanthum Wall.ex Choisy,G.zeylanicum(Gaertn.)A.Juss.,P.pinnata(L.)Merr.)growing today in or near the fossil localities.Extant but locally extinct species (for example, Mastixia arborea (Wight) C.B.Clarke, Shorea tumbuggaia Roxb., Persea parviflora (Meisn.) Harid.& R.R.Rao, Callerya cinerea(Benth.) Schot, Phyllanthus daltonii Müll.Arg., Calophyllum inophyllum L., Lindera bifaria Hosseus, Premna bengalensis C.B.Clarke etc.)grow in other parts of India,but do not occur in the present-day near the fossil localities.However,extant but regionallyextinct NLRs(Millettia extensa (Benth.) Benth.ex Baker, Dysoxylum costulatum Blume, Rourea caudata Planch., Sloanea dasycarpa Hemsl., Shorea leprosula Miq.,S.obtusa Wall.ex Blume etc.)have disappeared from India and now grow in other parts of the world.Given the apparent constancy of the climate in this region,we presume that significant changes in biotic interactions might be a reason for the disappearance of taxa from the present-day vegetation, including human activity.However,these taxa are presently growing in other regions of India,including the northeast region and south India,due to the availability of suitable conditions.If climate changes are involved,then theyare subtler than can be detected by the CLAMP proxygiven the existing statistical uncertainties.
TheupperPaleocene record of CalophylluminIndia(Bhattacharyya,1967; Ambwani,1992) suggests India is the centre of origin of this genus.Subsequently, this genus gradually became a ubiquitous component of the Neogene Siwalik forests of India and moved into other adjoining Southeast Asian regions,Polynesia,and the east coast ofAfricaasevidencedbylaterrecordsoutsideIndia(Khanetal.,2017a).Khan et al.(2017a) suggested that distinct, but modest, elevation changes in northeastern India,possibly related to Himalayan orogeny since the Miocene, might have caused the disappearance of Calophyllum inophyllum from the entire eastern Himalayas.
Based on fossil records Khan et al.(2016)suggested that Shorea was a common forest element in the Neogene (Miocene time)Siwalik forests of the eastern Himalayas.They also reviewed the historical phytogeography and highlighted the phytogeographic implications of this genus.
Several other low-elevation eastern Himalaya taxa have undergone range shifts since the mid-Miocene.According to Khan and Bera (2007), Dysoxylum costulatum probably migrated to the Malaya region after the Miocene.Khan and Bera (2017) suggested that Pinus was an important component of tropical-subtropical evergreen forest in the West Kameng district of Southeastern Himalaya during the Miocene but this conifer taxon subsequently declined from the local vegetation.At present, Rourea caudata(Connaraceae)does not grow in India and is confined to the tropical evergreen forest of southeast Asian regions (China and Myanmar)where conditions are more suitable.Khan and Bera (2016) suggested that this species of Connaraceae probably migrated to these Southeast Asian regions after lower Siwalik sedimentation(middleupper Miocene).
Khan et al.(2017b)reported the occurrence of the extant species Mastixia arborea (family Cornaceae) from the middle Miocene to early Pleistocene Siwalik sediments of Southeastern Himalaya.This report provided the first-ever fossil record of Mastixioids from India, as well as Asia.At present, M.arborea does not grow in the eastern Himalayas but is endemic to the tropical evergreen forests of the Western Ghats.They suggested that its extinction from the entire eastern Himalaya and probable movement to the Western Ghats is likely due to climate change(Siwalik forests experienced a weaker monsoon,i.e.,less rainfall seasonality than now)in the area,related to the Himalayan Orogeny during Miocene-Pleistocene times.They also suggested that its disappearance from presentday vegetation proximal to the fossil locality may be related to the gradual intensification of rainfall seasonality since the late Miocene.Such intensification is not evident in the CLAMP data.Similarly, More et al.(2018) reported Sloanea dasycarpa of Elaeocarpaceae from the Geabdat Sandstone Formation, Pliocene of Darjeeling foothills.They suggested that after the Pliocene the species might have migrated from the Darjeeling Himalayan region to adjoining southeast Asia (China, Myanmar, and Vietnam), the area of the present-day distribution of modern Sloanea, due to possible increasing aridity and rainfall seasonality,but again this is not evident in the data presented here.
12.Conclusion
This is the first thorough review of the eastern Himalayan Siwalik flora.Based on our overview of published palaeobotanical data from the Siwalik sediments,the following key points emerge:
(1) The eastern Himalaya has a rich Siwalik plant fossil record spanning the mid-Miocene to the Pleistocene, which, over the past two decades,has been investigated extensively with an increasing number of fossil taxa being reported.These fossil taxa are important for answering outstanding questions on plant diversity and floral evolution in this region.
(2) Plant species from the Cenozoic of the eastern Himalayas are diverse.To date, approximately 219 fossil species belonging to 162 genera of 42 families have been documented.They cover ferns, gymnosperm, and angiosperms, of which angiosperms are by far the most diverse, including 216 fossil species grouped into 159 genera of 39 families.In the fossil angiosperms, Fabaceae, Dipterocarpaceae, Lauraceae, Annonaceae, Euphorbiaceae, Calophyllaceae, Anacardiaceae,Apocynaceae,Rubiaceae,and Lythraceae are among the most diverse families; and Shorea, Dipterocarpus, Calophyllum,Millettia, Glochidion, Actinodaphne, Combretum, Bauhinia,Lagerstroemia, Uvaria, Rinorea Aubl., Sterculia, Ficus L., Terminalia,and Persea are among the most species-rich genera.
(3) Most of the families and genera represented in the fossil record are still part of the modern natural vegetation in the eastern Himalayas,but some taxa have disappeared.Thus far,fourteen taxa are known to have become extinct in the eastern Himalayas, namely Mastixia arborea, Shorea tumbuggaia,S.leprosula, S.obtusa, P.parviflora, C.cinerea, P.daltonii,C.inophyllum, L.bifaria, P.bengalensis, M.extensa,D.costulatum, R.caudata, and S.dasycarpa.In addition,phytogeographic exchanges of Siwalik elements of the eastern Himalayas with southeast Asia also occur.
(4) A gradual change in floral composition through the Siwalik succession is apparent.Floras changed significantly from the middle Miocene to the early Pleistocene through to today,but invoking climate change as the explanation is problematic as no distinctive(statistically significant)shift in climate metrics can be detected in foliar adaptations.If the climate contributed to these changes, it was through very subtle changes that affected overall taxon fitness, most likely intensification of rainfall seasonality.
(5) Here,we introduce new quantitative proxy palaeo-humidity measurements and explore new insights into the hydrological regime.Eastern Himalayan Siwalik forests experienced a monsoonal tropical warm humid climate.Tropical Siwalik forests of the eastern Himalaya prior to the Quaternary have a weaker monsoon (less rainfall seasonality)than now.
Declaration of competing interest
The authors declare that they have no competitive interest.
Author contributions
Data collection and analysis were performed by MAK,SM, RAS,TEVS,AA,TH and SB.The first draft of the manuscript was written by MAK,SM and all authors commented on previous versions of the manuscript.All authors read and approved the final manuscript.
Acknowledgments
MK and SM gratefully acknowledge the Department of Botany,Sidho-Kanho-Birsha University for providing infrastructural facilities to accomplish this work.SB acknowledges the Centre of Advanced Study (Phase-VII), the Department of Botany, the University of Calcutta for providing necessary facilities.We also thank Prof.Paul Valdes,Bristol University,U.K.,for providing the modern WorldClim2 climate data for the fossil localities.RAS and TEVS were supported by NERC/NSFC BETR Project NE/P013805/1.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2022.12.003.

- 植物多样性的其它文章
- Honoring a legend: Celebrating the life and legacy of Professor Heng Li
- The life of Xuan Zhou, founding father of the Gastrodia elata cultivation and industry in China
- Global patterns of taxonomic and phylogenetic diversity of flowering plants: Biodiversity hotspots and coldspots
- Distribution and conservation of near threatened plants in China
- Insights into cryptic speciation of quillworts in China
- Patterns of floristic inventory and plant collections in Myanmar