
光照和地下竞争对林冠下人工更新紫椴苗木形态和生物量分配的影响
2023-08-08 01:14于均屹徐立清张勇王庆成谢旭强袁钰晨王丹妮武慧蓉
森林工程 2023年4期
于均屹 徐立清 张勇 王庆成 谢旭强 袁钰晨 王丹妮 武慧蓉


摘 要:以次生林林冠下人工更新的2 年生紫椴苗木為研究对象,设置2种光照(强光(High light,HL,林隙),弱光(Low light,LL,林冠))和4种排除地下竞争(施肥(F)、切根(T)、切根加施肥(T+F)、对照(CK))处理,测定紫椴苗木形态特征和生物量分配,探究光照和地下竞争对林下人工更新紫椴苗木的影响。结果表明,1)切根和施肥对紫椴苗木苗高和地径生长量有显著的促进作用,其中T和T+F处理对苗木苗高和地径生长量的提高更显著,光照对紫椴苗木苗高和地径生长量无显著影响。2)紫椴苗木生物量和比叶重均受到光照和排除地下竞争措施交互作用的显著影响。8个处理组合中,HL下T和T+F处理对紫椴苗木的生物量积累和比叶重均有显著的促进作用,苗木生物量和比叶重均是HL下T+F处理的最大值,其次是HL下的T处理,LL下的CK处理苗木生物量均高于HL下的CK处理。3)T+F和T处理显著提高了紫椴苗木细根根长、根表面积和根体积,T+F处理较T处理增加更显著,光照对紫椴苗木细根形态无显著影响。综上,林冠下人工更新紫椴苗木时,林冠下适度遮荫的光环境更适宜紫椴苗木生长和更新,在林隙这种较强的光环境下进行紫椴人工更新时,应辅以切根或者切根加施肥的抚育措施,减少上层木根系对土壤资源的竞争以促进苗木生长。
关键词:紫椴(Tilia amurensis);人工更新;光照;地下竞争;生物量;形态特征
中图分类号:S725.7 文献标识码:A 文章编号:1006-8023(2023)04-0038-10
Effects of Light and Underground Competition on Seedling Morphology
and Biomass Allocation of Tilia amurensis Artificially
Regenerated Under Forest Canopy
YU Junyi, XU Liqing, ZHANG Yong, WANG Qingcheng*, XIE Xuqiang, YUAN Yuchen, WANG Danni, WU Huirong
(School of Forestry, Northeast Forestry University, Harbin 150040, China)
Abstract:The artificially regenerated 2 years Tilia amurensis seedlings under the canopy of the secondary forest were taken as the research object, and two kinds of light (high light (HL, gap) and low light (LL, canopy)) were set, and four treatments excluding underground competition (fertilization (F), root cutting (T), root cutting plus fertilization (T+F), and control (CK)) were used to determine the morphological characteristics and biomass allocation of Tilia amurensis, and to explore the influence of light and underground competition on artificial regeneration of Tilia amurensis. The results showed that, 1) root cutting and fertilization significantly promoted the seedling height and ground diameter growth of Tilia amurensis, among which T and T+F treatments significantly increased the seedling height and ground diameter growth, while light had no significant effect on the seedling height and ground diameter growth. 2) Biomass and specific leaf weight of Tilia amurensis were significantly affected by the interaction of light and measures to exclude underground competition. Among the 8 treatment combinations, T and T+F treatments under HL significantly promoted the biomass accumulation and specific leaf weight of Tilia amurensis seedlings. The seedling biomass and specific leaf weight of T+F treatment under HL were at the maximum values, followed by T treatment under HL. The seedling biomass of CK treatment under LL was higher than that of CK treatment under HL. 3) T+F and T treatments significantly increased the root length, root surface area and root volume of Tilia amurensis seedlings, and T+F treatment increased more significantly than T treatment. Light had no significant effect on the fine root morphology of Tilia amurensis seedlings. In conclusion, artificially regenerated Tilia amurensis under forest canopy, a moderately shaded light environment under canopy was more suitable for the growth and regeneration of Tilia amurensis. When artificially regenerated Tilia amurensis was carried out in the strong light environment such as forest gap, root cutting or root cutting plus fertilization should be supplemented to reduce the competition of upper layer wood roots for soil resources and promote seedling growth.
Keywords:Tilia amurensis; artificial regeneration; light; underground competition; biomass; morphology characteristics
收稿日期:2022-11-15
基金项目:中央高校基本科研业务费专项资金项目(2572020DR04)
第一作者简介:于均屹,硕士研究生。研究方向为森林培育。E-mail: 862900978@qq.com
通信作者:王庆成,博士,教授,博士生导师。研究方向为森林培育。E-mail: wqcnefu@163.com
引文格式:于均屹,徐立清,张勇,等.光照和地下竞争对林冠下人工更新紫椴苗木形态和生物量分配的影响[J].森林工程,2023,39(4):38-47.
YU J Y, XU L Q, ZHANG Y, et al. Effects of light and underground competition on seedling morphology and biomass allocation of Tilia amurensis artificially regenerated under forest canopy[J]. Forest Engineering, 2023, 39(4):38-47.
0 引言
随着天然林禁伐的全面实施,我国东北部林区的森林后备资源培育方式由原来皆伐迹地的全光下造林转化为林冠下的补植补造为主,造林地生境的变化对人工更新技术提出了全新的要求[1-2]。在林冠下人工更新苗木时,光和地下资源的可用性是影响苗木建立、存活和生长的重要因素[3],直接影响造林的成活率和保存率,制约我国大面积后备资源培育措施的实施成效,是森林后备资源培育有待解决的问题。
植物生长形态的可塑性是植物适应不同环境和资源水平的重要策略,决定了其生存和生产力的形成,以及植物在森林中分布的丰度和广度[4-5]。光作为树木叶片光合作用的唯一能量来源,是影响树种天然更新最重要的环境因子[6]。在不同的光环境中,苗木的形态和生物量分配会随着光照条件发生改变,进而影响苗木的生长和存活[7-8]。因树种自身的特异性,不同树种对光环境的变化有不同的适应性和可塑性。以往关于苗木对不同光环境响应的研究大部分都是在苗圃或者裸地通过遮荫网模拟光环境[9],并不能完全代表真实的林下光环境,而在自然条件下人工更新苗木对不同光照环境响应的研究涉及较少[10]。
在林冠下人工更新苗木时,上层木及地表植被不仅对地上光资源存在竞争,同时对土壤资源也存在强烈的竞争,导致苗木可利用土壤资源受到严重的限制[11-12]。针对这个问题,在国外切根和施肥被认为是减弱植物对土壤资源竞争的有效方法[13]。针对地下竞争,我国主要通过排除草本根系作为排除地下竞争对苗木的影响进行了一些研究,由于苗木自身的特异性也得到了不同的结果[14-15]。Devine等[16]研究发现上层乔木根系对新植苗木形態和生物量有很大影响,并且这种影响远大于林下植被。因此,对新植苗木排除地下竞争时,不应该仅考虑草本对植物的影响,应该加入上层木对新植苗木地下竞争的影响。而我国通过排除上层木根系的研究大多应用在农林复合经营和农田防护林方面,通过排除防护林树木根系对农作物土壤资源的竞争,可以有效地提高农作物的产量[17]。仅有少数将排除上层木根系作为地下竞争应用在人工更新造林方面[18]。除了排除根系竞争外,适当地施肥也可以增加苗木的生长和生物量的积累,促进根系生长,提高苗木对土壤资源的获取能力,从而增加对生物和非生物胁迫的抵抗力[19-20]。但是以往研究中,施肥目的并不是通过施肥解除来自上层木及地表植被根系对土壤资源竞争而对新植苗木产生的养分耗竭[19],且我国在林冠下人工更新时通过排除上层木及地表植被根系和施肥作为缓解地下竞争方式的研究涉及较少。同时,由于在林冠下人工更新时对苗木存活和生长的限制因素是相对不可控制的,在林冠下会混淆光照和土壤资源对苗木生长的影响[13,21]。因此,不应仅考虑光环境和排除地下竞争对苗木生长的影响,也应该考虑地上和地下竞争二者的交互作用。
紫椴(Tilia amurensis)是我国东北地区珍贵的阔叶树种,亦为红松阔叶林的主要伴生树种[22]。由于长期不合理釆伐,缺乏及时更新和抚育管理,紫椴资源数量和林分质量严重下降, 20世纪80年代起,我国开始营造紫椴人工林,但成林的较少[23-24]。以往研究中发现紫椴具有较强的耐阴性,是林冠下补植补造的首选树种之一[25]。因此,本研究以次生林林冠下人工更新紫椴苗木为研究对象,通过分析林冠下人工更新紫椴苗木形态和生物量分配对光照和地下竞争的响应,筛选出在林冠下人工更新紫椴苗木时,适宜紫椴生长的光环境和抚育措施,探究光照和地下竞争对林冠下人工更新紫椴苗木的影响机制,为紫椴后备资源培育提供实践和理论指导。
1 材料与方法
1.1 研究区概况
研究地点位于东北林业大学帽儿山实验林场尖砬沟森林培育试验站(127°26′~127°39′E,45°23′~45°26′N)。平均海拔约300 m,坡度为6°~15°,属温带大陆性季风气候,年均气温为2.8 ℃,最热月(7月)平均温度为23 ℃,最高气温为34.8 ℃;最冷月(1月)平均温度-23 ℃,最低气温为-44 ℃。年均降水量723 mm,多集中于7、8月份,占全年降水量的50%以上,年平均蒸发量1 093 mm。无霜期120~140 d,≥10 ℃的积温2 526 ℃。地带性植被为红松阔叶林,现存植被为原始植被遭干扰破坏后形成的硬阔叶混交林为主的天然次生林[26]。
样地设在次生林林冠下紫椴造林地,位于山坡的中上部,土壤为典型的暗棕壤。该林分于2021年4月进行了抚育间伐,形成了不同郁闭度的地块。2021年5月进行林冠下造林,苗木为紫椴2 年生实生苗苗木,来自吉林省临江市临江林业局望江楼苗圃。栽植前对新植苗木进行适当的修根,采用穴植法进行栽植,株行距为1 m×2 m。由于进行过抚育间伐,林下基本没有灌木。主要乔木树种有水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、黄菠萝(Phellodendron amurense)、紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、白桦(Betula platphylla)、五角槭(Acer mono)和白牛槭(Acer mandshuricum)等,林分内平均树高18.7 m,平均胸径21.8 cm,林分总密度560 株/hm2。
1.2 试验设计与处理
2021年5月中旬,在次生林紫椴造林地内设置一块20 m×50 m的样地,将样地平均划分为4块,每块内选择5个以“S”形分布的点,在0 ~ 20 cm土层取土测定土壤理化性质,测得土壤pH为4.45、有机质为188.87 g/kg、碱解氮为742.61 μg/g,有效磷为7.73 μg/g,速效钾为440.99 μg/g,土壤容重为0.51 g/cm3,各小样地间土壤容重以及土壤速效性养分之间差异均不显著。在样地内选择林隙和林冠2种林下光环境分别代表强光(High light ,HL)和弱光(Low light ,LL)。于7月份选择晴朗天气采用DRM-FQ光量子双辐射计(Spectrum Technologies,Inc,USA)测定紫椴造林地内林隙和林冠下每株苗木栽植点的光照强度,测定时间为8:00—16:00,每小时测定1次,测定时间控制在10 min以内。最终测得林隙内光照强度为9 700~28 825 lx,林冠下光照强度为1 725~4 803 lx。
于生长季开始(5月初)测定紫椴苗木的苗高和地径(平均苗高为48.9 cm,地径为8.0 mm),测定时在地径位置做好标记,并对每株苗木挂牌标记。强光下苗木总数为88 株,弱光下为80 株。采取完全随机试验设计,对2种光照条件下栽植的苗木采取对照(CK)、切根(T)、施肥(F)、切根+施肥(T+F)进行处理。强光下平均每种处理22 株,弱光下平均每种处理20 株。2021年7月9日对2种光照强度样地内的苗木进行上述4种处理。
切根是以苗木为中心,距离苗木周围各50 cm,挖30 cm深环状的沟,排除上层木及地表植被对土壤资源的竞争,然后用土壤回填,保持水文连续性。因为北方森林主要侧根和细根(占所有根的80%~ 90%)一般集中在土壤顶部30 cm [27],为防止上层木根系由于生长再次进入之前切根的区域与苗木进行土壤养分竞争,因此1 a进行2次切根,切根时间为2021年7月9日和8月7日。
施肥量按土壤本底碱解氮的10%进行施肥,测定土壤碱解氮为742.61 μg/g,得到施氮量为4.46 g/株,换算成奥绿缓释肥315 s(荷兰, 易乐施国际公司,Everris International B.V.生产,N∶P2O5∶K2O=15%∶9%∶12%),为29.73 g/株。该缓释肥由有机树脂包膜,缓释期为5~ 6个月。施肥方式是在距离苗木15 cm处挖一圈3~5 cm的沟,将肥料均匀地撒在沟里,然后用土壤覆盖,防止肥料损耗。施肥时间是2021年7月9日。
1.3 指标测量
在2021年生长季结束(9月末)时,分别用钢卷尺和游标卡尺测量所有苗木的苗高、地径。生长季结束时,每个处理随机选取3株长势基本一致的苗木,将苗木全株取出,分成根、茎、叶3部分装入封口袋中,放入冷藏箱中带回实验室。待叶面积扫描完,将茎、叶分别装入信封中,放入烘箱,65 ℃烘至恒重,用电子天平(±0.01 g)测其生物量。待根系形态测定完成后,烘干测根系生物量。
1.3.1 根系形态指标测定
用去离子水轻轻冲洗除去根系表面的土壤残留颗粒和其他杂质,将直径小于2 mm的根系使用Epson数字化扫描仪( Expression 10 000XL1.0,Epson Electronics Inc.,San Jose,USA) 扫描,根系图像采用WinRHIZORA(pro2004b)软件(Regent Instruments Canada.)进行形态特征分析,测得根长、根表面积和根体积等指标。
1.3.2 比叶重的测定
将带回的叶样用手持式激光叶面积仪测定(CI-203, 美国)测定叶面积,每株苗取5片叶子作为重复。对每株苗木的5片叶子进行标记,测定完叶面积数据后,在105 ℃下杀青10 min,后用65 ℃烘至恒重,电子天平(±0.01 g)称其质量,即可计算出比叶重(Leaf mass per area, LMA),计算公式如下:比叶重=叶片干重/叶面积。
1.4 数据分析
采用SPSS19.0(SPSS for windows, USA)进行数据分析。将光照强度和抚育措施作为2个因素,利用单因素和双因素方差分析及多重比较( LSD) 方法对不同处理数据进行差异显著分析。当存在交互作用时,探究8个处理组合对该指标的影响。采用SigmaPlot12.5(Systat, USA)软件作图。所有数据为平均值±标准误。
2 结果与分析
2.1 切根、施肥和光照对林冠下人工更新紫椴苗木苗高、地径生长量的影响
切根和施肥对紫椴苗木苗高和地径生长量影响显著(P<0.05),其中F、T和T+F处理较CK处理苗木的苗高生长量分别提高了73.1%、110.8%、134.5%(P<0.05),T+F处理较F处理苗木的苗高生长量提高了35.5%(P<0.05);F、T和T+F處理较CK处理均促进了苗木地径的生长,其中T和T+F处理较CK处理苗木的地径生长量分别提高了113.9%和106.2%(P<0.05),其余处理间无显著差异(P>0.05)。光照以及光照和切根、施肥的交互作用对紫椴苗木的苗高和地径生长量均无显著影响(P>0.05)(表1)。
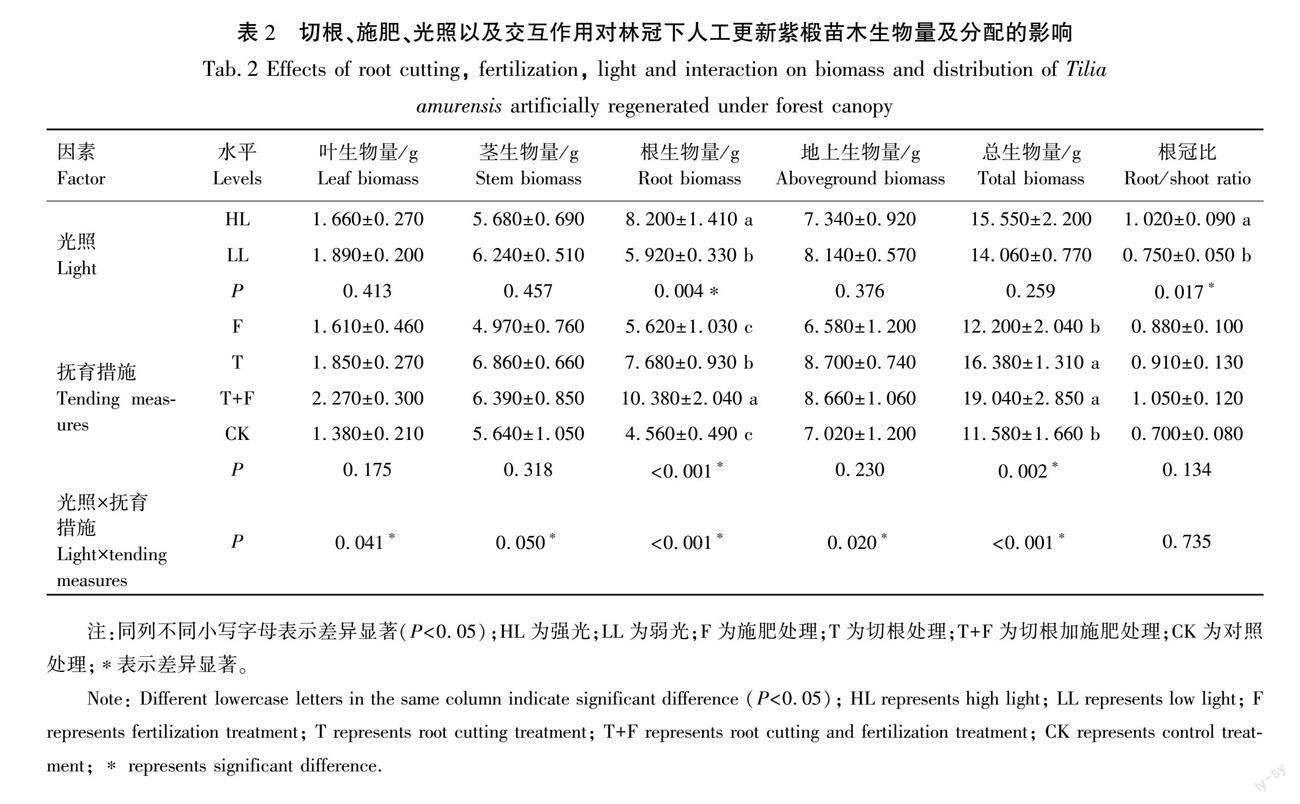
2.2 切根、施肥和光照对林冠下人工更新紫椴苗木生物量及其分配的影响
光照对苗木根生物量和根冠比有显著影响(P<0.05),HL处理较LL处理分别提升了38.5%和36%(P<0.05)(表2)。苗木根生物量和总生物量受到切根和施肥的显著影响(P<0.05),两者从大到小均为T+F、T、F、CK,其中T+F和T处理较其他处理提高了苗木根系生物量和总生物量(P<0.05),T+F处理较T处理苗木的根系生物量提高了35.2%(P<0.05),二者对总生物量没有显著影响(P>0.05),其余处理之间均无显著影响(P>0.05)。
光照和切根、施肥的交互作用对根冠比的影响不显著(P>0.05),对其余所有生物量均有显著影响(P<0.05)(表2)。HL下切根和施肥对苗木生物量有显著影响(P<0.05),而在LL下切根和施肥对苗木生物量无显著影响(P>0.05)(表3)。在HL下,苗木生物量最大值均出现在T+F处理中,T+F处理下根生物量和总生物量较T处理分别提高55%和30.7%(P<0.05), T+F和T处理较F和CK处理提高了苗木的所有生物量(P<0.05),其余处理间均无明显差异(P>0.05)。LL下的CK处理苗木生物量较HL处理下的CK处理均有提高,其中对苗木的茎生物量、地上生物量和总生物量的提高显著,分别提高了77.8%、80.8%、69.6%(P<0.05);HL下的T+F和T处理苗木生物量均高于LL下的T+F和T处理,其中LL下的T+F处理的根生物量和总生物量较HL下分别降低了59.6%、47.3%(P<0.05),LL下的T处理的根生物量较HL下分别降低了39%(P<0.05);不同光照下各生物量F处理之间的比较,仅LL下F处理较HL下的F处理显著提高了苗木的叶生物量(P<0.05),对其余生物量无显著影响(P>0.05)。
2.3 切根、施肥和光照对林冠下人工更新紫椴苗木比叶重的影响
切根、施肥、光照以及交互作用均对苗木比叶重有显著影响(P<0.05)(表4)。苗木比叶重随光照强度的减弱而降低,LL下苗木比叶重较HL降低了12%(P<0.05);T+F处理苗木比叶重较F和CK处理分别提高了18.4%和14.1%(P<0.05),其余处理之间无显著差异(P>0.05)。当考虑交互作用时,LL下切根和施肥对苗木的比叶重无显著影响(P>0.05);HL下切根和施肥对苗木的比叶重存在显著影响(P<0.05),T+F处理较F和CK处理苗木的比叶重分别提高了40.4%、25.9%(P<0.05),T处理较F和CK处理苗木的比叶重分别提高了33.6%和19.7%(P<0.05)(图1)。HL下T+F和T处理较LL下苗木的比叶重分别提高了32.1%和29.2%(P<0.05),其余处理间无显著差异(P>0.05)。
2.4 切根、施肥和光照对林冠下人工更新紫椴苗木细根形态的影响
切根、施肥、光照的交互作用以及光照对苗木细根形态无显著影响(P>0.05),但是也能够看到HL下苗木细根的根长、根表面积、根体积较LL均有所增加(表5)。苗木细根形态受到切根和施肥的显著影响(P<0.05),苗木细根形态最大值均处于T+F处理中,根长和根表面积由大到小依次均为T+F、T、F、CK,且T+F处理下苗木根长和根表面积均显著高于F处理、T处理、CK处理(P<0.05),T处理较CK处理显著提高了苗木的根长和根表面积(P<0.05),其余处理间无显著差异(P>0.05);T+F处理下苗木根体积显著高于F和CK处理(P<0.05),其余处理之间均无显著差异(P>0.05)。
3 讨论
3.1 切根、施肥和光照对林冠下人工更新紫椴苗木生长的影响
苗高和地径是衡量苗木品质的重要指标[28],可以直观反映植物的生长状况。本研究发现,光照对紫椴苗木的苗高和地径的生长无显著影响,而F、T和T+F处理较CK处理均促进了苗高和地径的生长(表1),说明增加土壤资源有利于新植紫椴苗木的苗高和地径生长。这与Promis等[29]的研究结果一致,2个生长季后增加土壤资源同样促进了苗木苗高和地径的生长,即林冠下人工更新苗木时,新栽植的苗木根系较浅使其容易受到地下竞争的影响,通过提高土壤养分有效性能够排除或者减弱地下竞争对苗木产生的影响,从而促进苗木的生长。本研究还发现,T+F和T处理较F和CK处理显著增加了苗木的苗高和地径生长量,说明切根较施肥更有利于苗木苗高和地径的生长。与Platt等[13]的研究结果一致,2个生长季后切根和切根加施肥较对照促进了山毛榉苗高和地径的生长,即只有在保护新植苗木免受来自上层优势树木根系对土壤资源的竞争,才能更好地促进苗木的生长。
3.2 切根、施肥和光照对林冠下人工更新紫椴苗木生物量及其分配的影响
植物生物量的积累主要受到环境中可利用资源的影响,如光照、水分和养分等,生物量的分配是植物对所处环境条件的响应和适应的结果[30]。本研究发现,强光下根冠比显著高于弱光(表2),说明紫椴通过调整根冠比,遵循生物量最优分配策略,将生物量分配给最需要的器官以适应不同的光照环境。这与张玲等[9]的研究结果一致,在1个生长季后遮荫处理降低了黄檗的根冠比,即弱光环境下苗木通过降低自身的根冠比来增加苗木地上部分接触面积,争取更多的光照资源来适应弱光环境;强光环境下苗木光照资源充足,苗木为维持正常的水分吸收和蒸腾,保证较高的光合速率,会减少地上部分生物量的分配[31]。本研究还发现切根和施肥对根系生物量和总生物量有显著的提升,说明增加土壤资源有利于紫椴苗木根系生物量的积累,从而促进苗木总生物量的快速积累。以往的研究认为,土壤资源不是首要的限制因子时,苗木会减少地下生物量的分配,但土壤资源成为限制性因子时,苗木缺少养分,植物会将生物量分配到根系从而吸收更多的养分[32-33],但是本研究发现,增加土壤资源后苗木的根系生物量反而有所增加,可能是当土壤资源增加时,根系快速生長的结果,以最大限度地提高短期养分吸收[34]。这种情况仅在强光条件下有效果,在弱光条件下增加土壤养分对苗木生物量均无显著影响(表3)。可能是弱光条件下掩盖了土壤资源对苗木生长的效益,而在强光下土壤资源对苗木生长的效益得到放大[35]。
从切根、施肥和光照的交互作用中发现,弱光下的对照处理苗木生物量均高于强光的对照处理(表3),说明幼年的紫椴苗木具有一定的耐阴性[25],在适度的遮荫环境下对紫椴苗木生物量积累有促进作用。这与孙欣欣[36]的研究结果一致,即适度遮荫更有利于新植紫椴苗木生物量的积累。本研究还发现,强光下的T+F和T处理苗木生物量均显著高于F和CK处理,说明切根缓解地下竞争的效果较施肥更显著,可能是切根不仅会增加土壤养分,对土壤水分也有所改善[37],而施肥仅改善土壤养分,因为植物在吸收养分的过程中水分也起到了很重要的作用[38]。且本研究还发现,强光下T+F和T处理生物量均高于弱光下各处理的生物量,进一步说明在强光下增加土壤资源对紫椴苗木生物量的积累更有益。即适度遮荫更有利于苗木生物量的积累,在强光下切根和切根加施肥对紫椴苗木的生物量积累促进作用更明显。
3.3 切根、施肥和光照对林冠下人工更新紫椴苗木比叶重的影响
单位叶面积上叶片生物量的分配是一种对生长环境的重要适应策略[5]。本研究发现紫椴苗木比叶重随光照强度的减弱而显著降低(表4),这与徐海东等[39]的研究结果一致,1个生长季后随着光照强度的降低显著降低了麻栎单位面积叶片质量,说明强光下苗木通过增大叶片的厚度来减缓光在内部的传导,进而避免强光对光合机构造成破坏,同时苗木也会减小叶面积来降低蒸腾,避免水分的损耗;弱光下苗木比叶重显著降低,说明弱光下苗木采取扩展叶片增加叶面积的策略,获取更多的光照资源以适应弱光环境。本研究还发现,强光下T和T+F处理显著增加了苗木的比叶重,弱光下增加土壤资源对苗木比叶重无显著影响(图1)。说明增加土壤资源只有在强光下能够发挥作用,促进紫椴苗木更好地进行光合作用。因为植物从土壤吸收的养分依赖于光合作用产生的碳投入,所以苗木需要进行更多的光合作用获取更多的碳以维持苗木的生长[40]。因此,地上生物量的增加,叶片变厚,比叶重变大会避免强光对光合机构造成破壞,从而能够更好地进行光合作用。这与张书娜等[10]研究结果一致,即强光下增加土壤资源后,苗木为维持土壤养分增加后苗木的正常生长,会通过增加比叶重的策略来获取更多光照资源,更好地进行光合作用。
3.4 切根、施肥和光照对林冠下人工更新紫椴苗木细根形态的影响
细根是植物吸收养分和水分的重要器官,植物根系获取养分主要由直径小于2 mm的根系发挥重要作用[41-42]。植物根系形态具有一定的可塑性,可以反映植物对土壤养分的利用策略,对于植物在竞争中获得优势具有重要意义,是衡量植物生产力的重要指标[43-44]。本研究发现,切根和施肥对细根形态影响显著,而光照对细根形态没有显著影响,F、T、T+F处理苗木细根形态指标均高于CK处理(表5),说明细根形态主要受到土壤资源环境的影响,增加土壤资源对苗木细根形态的建成有促进作用。本研究还发现T和T+F处理下苗木细根形态均显著高于CK处理,而施肥只有与切根结合时才能够显著增加细根形态,说明切根较施肥能够更好地建立完善的根系结构。因为排除上层林木根系后从根本上排除了源头的问题,减少上层木根系对土壤养分和水分的竞争,而施肥仅增加了土壤养分,土壤水分得不到改善,水分短缺可能会抑制根系对养分的吸收速率[31]。这与Petri瘙塅an等[37]的研究结果一致,山毛榉和花旗松的细根形态参数都随着切根引起土壤水分和养分供应的改善而增加。即高有效性的养分投入能够降低苗木根系对土壤资源获取能力的限制,促进细根的生长以及增大根系在土壤中吸收养分的范围。
4 结论
综上所述,适度遮荫促进了紫椴苗木生物量的积累,增加紫椴苗木对外界环境胁迫的抵抗力,进而使苗木更好地生长。切根和切根加施肥的抚育措施显著促进了紫椴苗木的生长和生物量积累,且在强光下效果才显著,在弱光下切根和切根加施肥对紫椴苗木的生长无显著影响。因此,林冠下人工更新紫椴苗木时,林冠下适度遮荫的光环境更适宜紫椴苗木生长和更新,在林隙这种较强的光环境下进行紫椴人工更新时,应辅以切根或者切根加施肥的抚育措施减少上层木根系对土壤资源的竞争以促进苗木生长。需要注意的是本研究采用的施肥和切根的方法在实施过程中会投入大量的精力,大面积施用2种方法时,要考虑一定的经济效益。本研究中施肥和切根对紫椴苗木部分生长指标有促进作用,但是影响不显著,光照对紫椴苗木的苗高、地径和根系形态指标影响不够显著,可能是本试验仅对紫椴苗木形态生长指标进行了1个生长季的研究,随着时间的推移,增加土壤养分和不同光照条件可能会对紫椴苗木的生长产生显著的不同的影响,有待开展进一步的研究。
【参 考 文 献】
[1]庄作峰.中国天然林资源保护工程——世纪之交的重大生态工程[J].世界林业研究,2001(3):47-54.
ZHUANG Z F. A great ecological project: China natural forest conservation programme[J]. World Forestry Research, 2001(3): 47-54.
[2]于立忠,刘利芳,王绪高,等.东北次生林生态系统保护与恢复技术探讨[J].生态学杂志,2017,36(11):3243-3248.
YU L Z, LIU L F, WANG X G, et al. Discussion on the protection and restoration technology of secondary forest ecosystems in Northeast China[J]. Chinese Journal of Ecology, 2017, 36(11): 3243-3248.
[3]LIEFFERS V J, MESSIER C, STADT K J, et al. Predicting and managing light in the understory of boreal forests[J]. Canadian Journal of Forest Research, 1999, 29(6): 796-811.
[4]MCCONNAUGHAY K D M, COLEMAN J S. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients[J]. Ecology, 1999, 80(8): 2581-2593.
[5]冯玉龙,曹坤芳,冯志立,等.四种热带雨林树种幼苗比叶重,光合特性和暗呼吸对生长光环境的适应[J].生态学报,2002(6):901-910.
FENG Y L, CAO K F, FENG Z L, et al. Acclimation of lamina mass per unit area, photosynthetic characteristics and dark respiration to growth light regimes in four tropical rainforest species[J]. Acta Ecologica Sinica, 2002(6): 901-910.
[6]朱教君.次生林經营基础研究进展[J].应用生态学报,2002(12):1689-1694.
ZHU J J. A review on fundamental studies of secondary forest management[J]. Chinese Journal of Applied Ecology, 2002(12): 1689-1694.
[7]夏婵,李何,王佩兰,等.不同光照强度对赤皮青冈幼苗光合特性的影响[J].中南林业科技大学学报,2021,41(7):72-79.
XIA C, LI H, WANG P L, et al. Effects of shading on the photosynthetic characteristics of Cyclobalanopsis gilva seedlings [J]. Journal of Central South University of Forestry & Technology, 2021, 41(7): 72-79.
[8]徐飞,郭卫华,徐伟红,等.不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J].生态学报,2010,30(12):3098-3107.
XU F, GUO W H, XU W H, et al. Effects of light intensity on growth and photosynthesis of seedlings of Quercus acutissima and Robinia pseudoacacia[J]. Acta Ecologica Sinica, 2010, 30(12): 3098-3107.
[9]张玲,张东来.遮荫条件下黄檗生长和生理响应的性别差异研究[J].植物研究,2020,40(5):735-742.
ZHANG L, ZHANG D L. Gender differences in growth and physiological response of Phellodendron amurense Rupr. in condition of overshadow[J]. Plant Research, 2020, 40(5): 735-742.
[10]张书娜,王庆成,郝龙飞,等.光照和施肥对白桦林冠下水曲柳、胡桃楸苗木生长的影响[J].森林工程,2015,31(2):51-56.
ZHANG S N, WANG Q C, HAO L F, et al. Effects of light and fertilization on the growth of Fraxinus mandshurica and Juglans mandshurica seedlings under the canopy of Betula platyphylla secondary forest[J]. Forest Engineering, 2015, 31(2): 51-56.
[11]DEVINE W D, HARRINGTON T B. Belowground competition influences growth of natural regeneration in thinned Douglas-fir stands[J]. Canadian Journal of Forest Research, 2008, 38(12): 3085-3097.
[12]AMMER C, STIMM B, MOSANDL R. Ontogenetic variation in the relative influence of light and belowground resources on European beech seedling growth[J]. Tree physiology, 2008, 28(5): 721-728.
[13]PLATT K H, ALLEN R B, COOMES D A, et al. Mountain beech seedling responses to removal of below-ground competition and fertiliser addition[J]. New Zealand Journal of Ecology, 2004, 28(2): 289-293.
[14]唐毅,蒋德明,陈卓,等.地上竞争与地下竞争对科尔沁沙地榆树幼苗生长的影响[J].应用生态学报,2011,22(8):1955-1960.
TANG Y, JIANG D M, CHEN Z, et al. Effects of aboveground and belowground competition between grass and tree on elm seedlings growth in Horqin sandy land[J]. Chinese Journal of Applied Ecology, 2011, 22(8): 1955-1960.
[15]张静静,刘宁,南宏伟,等.草本地下竞争对3种幼苗存活和生长的影响[J].中南林业科技大學学报,2019,39(1):71-76.
ZHANG J J, LIU N, NAN H W, et al. Effects of herb underground competition on seedlings survival rate and seedlings height growth of three one-year-old seedlings[J]. Journal of Central South University of Forestry & Technology, 2019, 39(1): 71-76.
[16]DEVINE W D, HARRINGTON T B. Belowground competition from overstory trees influences Douglas-fir sapling morphology in thinned stands[J]. New forests, 2009, 37(2): 137-153.
[17]田福军.杏-农复合系统大扁杏林带根系的分布规律及对农作物生长的影响[J].辽宁林业科技,2014(3):23-25.
TIAN F J. Distribution of root system and its effect on crop growth in apricot almond plantation system[J]. Liaoning Forestry Science and Technology, 2014(3): 23-25.
[18]张命军.帽儿山实验林场杂木林主要树种天然更新影响因子研究[D].哈尔滨:东北林业大学,2012.
ZHANG M J. Factors influencing natural regeneration of major tree species in mixed broad-leaned forests in Maoershan forest farm, Heilongjiang Province[D]. Harbin: Northeast Forestry University, 2012.
[19] OLIET J A, PLANELLES R, ARTERO F, et al. Nursery fertilization and tree shelters affect long-term field response of Acacia salicina Lindl. planted in Mediterranean semiarid conditions[J]. Forest Ecology and Management, 2005, 215(1-3): 339-351.
[20]杨阳,张德鹏,及利,等.配比施肥对紫椴播种苗生长、养分积累及根系形态的影响[J].中南林业科技大学学报,2021,41(9):63-70.
YANG Y, ZHANG D P, JI L, et al. Effects of formula fertilization on growth, nutrient accumulation and root morphology of Tilia amurensis seedlings[J]. Journal of Central South University of Forestry & Technology, 2021, 41(9): 63-70.
[21]CARSWELL F E, DOHERTY J E, ALLEN R B, et al. Quantification of the effects of aboveground and belowground competition on growth of seedlings in a conifer–angiosperm forest[J]. Forest Ecology and Management, 2012, 269: 188-196.
[22]史凤友,陈喜全,陈乃全,等.椴树落叶松人工混交林的研究[J].东北林业大学学报,1991(S1):45-53.
SHI F Y, CHEN X Q, CHEN N Q, et al. Research on the artificial mixed forest of Tilia amurensis and Larix olgensis[J]. Journal of Northeast Forestry University, 1991(S1): 45-53.
[23]臧思航,张永鑫,范春楠,等.紫椴种子休眠的影响因素[J].北华大学学报(自然科学版),2021,22(3):308-312.
ZANG S H, ZHANG Y X, FAN C N, et al. Influence factors of Tilia amurensis seeds dormancy[J]. Journal of Beihua University (Natural Science Edition), 2021, 22(3): 308-312.
[24]徐程扬.紫椴幼苗、幼树对光的响应与适应研究[D].北京:北京林业大学,1999.
XU C Y. Responses and adaptations of Tilia amurensis seedlings and saplings to light regimes[D]. Beijing: Beijing Forestry University, 1999.
[25]王书凯.辽东山区主要树种耐阴性的研究[D].哈尔滨:东北林业大学,2004.
WANG S K. Study on tree toleration in the forests in the east of Liaoning Province[D]. Harbin: Northeast Forestry University, 2004.
[26]潘建平,赵克尊.东北林业大学帽儿山实验林场资源增长与生态环境建设的研究[M].哈尔滨.东北林业大学出版社,2007.
PAN J P, ZHAO K Z. Research on resource growth and ecological environment construction of Maoershan Experimental Forest Farm in Northeast Forestry University[M]. Harbin: Northeast Forestry University Press, 2007.
[27]JACKSON R B, CANADEL J, EHLERINGER J R, et al. A global analysis of root distributions for terrestrial biomes[J]. Oecologia, 1996, 108(3): 389-411.
[28]刘欢,王超琦,吴家森,等.氮素指数施肥对1年生杉木苗生长及氮素积累的影响[J].浙江农林大学学报,2017,34(3):459-464.
LIU H, WANG C Q, WU J S, et al. Growth and N accumulation in seedlings of Cunninghamia lanceolata clones with N exponential fertilization[J]. Journal of Zhejiang A&F University, 2017, 34(3): 459-464.
[29]PROMIS A, ALLEN R B. Tree seedlings respond to both light and soil nutrients in a Patagonian evergreen-deciduous forest[J]. PLoS One, 2017, 12(11): e0188686.
[30]劉青青, 马祥庆, 李艳娟, 等. 杉木种子萌发及幼苗生长对光强的响应[J]. 应用生态学报, 2016, 27(12): 3845-3852.
LIU Q Q, MA X Q, LI Y J, et al. Response of seed germination and seedling growth of Chinese fir to different light intensities[J]. Chinese Journal of Applied Ecology, 2016, 27(12): 3845-3852.
[31]CLARKA B, BULLOCK S. Shedding light on plant competition: modelling the influence of plant morphology on light capture (and vice versa)[J]. Journal of Theoretical Biology, 2007, 244(2):208-217.
[32]LEWIS S L, TANNER E V J. Effects of above-and belowground competition on growth and survival of rain forest tree seedlings[J]. Ecology, 2000, 81(9): 2525-2538.
[33]RIEGEL G M, MILLER R F, KRUEGER W C. Competition for resources between understory vegetation and overstory Pinus ponderosa in northeastern Oregon[J]. Ecological applications, 1992, 2(1): 71-85.
[34]HUTCHINGS M J, JOHN E A, WIJESINGHE D K. Toward understanding the consequences of soil heterogeneity for plant populations and communities[J]. Ecology, 2003, 84: 2322-2334.
[35]COOMES D A, GRUBB P J. Impacts of root competition in forests and woodlands: a theoretical framework and review of experiments[J]. Ecological Monographs, 2000, 70(2): 171-207.
[36]孙欣欣.遮荫对胡桃楸和紫椴苗木形态和生理的影响[D].哈尔滨:东北林业大学,2013.
SUN X X. Effect of shading on morphology and physiology in Juglans mandshurica and Tilia amurensis seedlings[D]. Harbin: Northeast Forestry University, 2013.
[37]PERTRITAN I C, VON LUPKE B, PETRITAN A M. Effects of root trenching of overstorey Norway spruce (Picea abies) on growth and biomass of underplanted beech (Fagus sylvatica) and Douglas fir (Pseudotsuga menziesii) saplings[J]. European Journal of Forest Research, 2011, 130(5): 813-828.
[38]徐明岗,张一平,张君常.根—土界面养分运移研究进展[J].土壤通报,1997(1):47-50.
XU M G, ZHANG Y P, ZHANG J C. Research progress on nutrient transport at the root-soil interface [J]. Chinese Journal of Soil Science, 1997(1): 47-50.
[39]徐海东,熊静,成向荣,等.麻栎和闽楠幼苗叶功能性状及生物量对光照和施肥的响应[J].生态学报,2021,41(6):2129-2139.
XU H D, XIONG J, CHENG X R, et al. Responses of leaf functional traits and biomass of Quercus acutissima and Phoebe bournei seedings to light and fertilization[J]. Acta Ecologica Sinica, 2021, 41(6): 2129-2139.
[40]霍常富,孙海龙,王政权,等.光照和氮营养对水曲柳苗木生长及碳-氮代谢的影响[J].林业科学,2009,45(7):38- 44.
HUO C F, SUN H L, WANG Z Q, et al. Effects of light and nitrogen on growth, carbon and nitrogen metabolism of Fraxinus mandshurica seedlings[J]. Scientia Silvae Sinicae, 2009, 45(7): 38-44.
[41]CHEN W, ZENG H, EISSENSTAT D M, et al. Variation of first-order root traits across climatic gradients and evolutionary trends in geological time[J]. Global Ecology and Biogeography, 2013, 22(7): 846- 856.
[42] MCCORMACK M L, DICKIE I A, EISSENSTAT D M, et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes[J]. New Phytologist, 2015, 207(3): 505- 518.
[43]姚甲宝,楚秀丽,周志春,等.不同養分环境下邻株竞争对木荷和杉木生长、细根形态及分布的影响[J].应用生态学报,2017,28(5):1441-1447.
YAO J B, CHU X L, ZHOU Z C, et al. Effects of neighbor competition on growth, fine root morphology and distribution of Schima superba and Cunninghamia lanceolata in different nutrient environments[J]. Chinese Journal of Applied Ecology, 2017, 28(5): 1441-1447.
[44]LEUSCHNER C, HERTEL D, SCHMID I, et al. Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility[J]. Plant and Soil, 2004, 258(1): 43-56.
猜你喜欢
中国机械工程(2022年8期)2022-05-09
Chinese Physics B(2021年9期)2021-09-28
中国机械工程(2021年8期)2021-05-07
今日农业(2020年19期)2020-12-14
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
音乐教育与创作(2019年8期)2019-05-16
中国科技信息(2015年2期)2015-11-16
小小说月刊·下半月(2015年11期)2015-05-14
植物营养与肥料学报(2014年1期)2014-03-11
