
猪繁殖与呼吸综合征病毒GP3蛋白单克隆抗体制备及抗原表位鉴定
2023-08-25 09:03孙杨杨张路捷孙海凤
畜牧兽医学报 2023年8期
袁 丽,孙杨杨,张路捷,张 杰,孙海凤,白 娟,姜 平
(南京农业大学农业农村部动物细菌学重点实验室,南京 210095)
猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)是影响世界养猪业的重要病原,临床上可引起母猪繁殖障碍、仔猪高死亡率和各年龄阶段猪的呼吸系统症状,给我国养猪业带来了巨大经济损失[1-2]。PRRSV 可分为欧洲型(PRRSV-1)和美洲型(PRRSV-2),二者的基因相似性为50%~70%,通过对ORF5序列的系统发育分析,PRRSV-1分为4个亚型,PRRSV-2分为9个谱系[3-4]。我国主要流行PRRSV-2,包括高致病性PRRSV(亚系8.7)、经典PRRSV(亚系5.1)、类NADC30毒株(亚系1.8)及QYYZ毒株(亚系3.5)等基因亚型毒株[5]。
PRRSV 基因组为单股正链 RNA,长度约为15 kb,包含至少 9 个开放阅读框(ORFs)[6-7]。其中ORF3编码的糖基化蛋白3(glycosylated protein 3,GP3)具有7个糖基化位点[8]。GP3蛋白由一个切割的信号肽、一个高度糖基化的结构域、一个短疏水区和一个未糖基化的C末端结构域组成。病毒颗粒中GP3与GP2、GP4蛋白形成异源三聚体,并与宿主细胞CD163相互作用,而CD163是 PRRSV进入细胞的关键受体[9-10]。GP3蛋白也可表现为细胞分泌形式,分泌的GP3可能作为一种诱饵,分散抗体远离病毒颗粒[11]。病毒颗粒中GP3含量较少,但抗原性较高,参与诱导中和抗体产生[12]。目前,PRRSV GP3蛋白结构和抗原特性尚不十分清楚。
本研究根据PRRSV FJ1402毒株序列扩增ORF3基因,通过原核表达系统成功制备出重组GP3蛋白,将其作为免疫原免疫BALB/c小鼠,制备了7株GP3单克隆抗体,鉴定出4个B细胞表位,为 PRRSV诊断和生物学研究奠定了基础。
1 材料与方法
1.1 主要材料
PRRSV FJ1402毒株、PRRSV N单抗、杆状病毒系统表达PRRSV GP3蛋白(片段为28—203 aa,表达系统为pFastBacTMDual )、SP2/0细胞、大肠杆菌E.coliDH5α和Rosetta均由本实验室保存。雌性BALB/c小鼠(6~8 周龄)购自扬州大学实验动物中心。
2×Phanta Max Master Mix和Pres-tained Protein Marker,购自南京诺唯赞生物科技股份有限公司;限制性核酸内切酶和T4 DNA 连接酶,购自Thermo Fisher Scientific公司;山羊抗小鼠IgG(H+L)-HRP和HRP-SPA,购自上海碧云天生物技术有限公司;HisSep Ni-NTA Agarose Resin,购自上海翊圣生物科技有限公司;RPMI-1640培养基、胎牛血清,购自美国 Gibco 公司;FITC-羊抗鼠 IgG、小鼠单克隆抗体亚型鉴定试剂盒,购自Proteintech生物科技有限公司;TanonTM High-sig ECL Western blot底物试剂盒,购自天能科技有限公司;异丙基硫代-β-D-半乳糖(IPTG)、PEG1450、50×HAT和50×HT,购自美国Sigma公司;其他试剂均为国产分析纯。
1.2 重组质粒构建
根据PRRSV FJ1402(GenBank No.KX169 191.1) ORF3基因序列,采用TMHMM Serverv.2.0 网站软件分析GP3蛋白跨膜区为16~38位氨基酸(aa),删除1~38 aa后的目的片段命名为 tGP3 (39~254 aa)。引物为 28a-tGP3-F:5′-CGGGATCCGTTAGGGGCAACTTCTCTTT-3′,28a-tGP3-R:5′-CCCTCGAGTTGCCGCGCGACATTGAGGA-3′,用于扩增 tGP3 片段,大小为 648 bp。PCR 反应条件:95 ℃ 3 min;95 ℃ 30 s,60 ℃ 15 s,72 ℃ 60 s,循环35 次;72 ℃ 5 min。将目的片段和 pET-28a 载体通过BamHⅠ和XholⅠ双酶切后进行连接,连接产物转化至DH5α菌,经菌液PCR 和酶切鉴定正确后送通用生物公司测序,结果显示正确,重组质粒pET-28a-tGP3于-20 ℃ 保存备用。
为了解析GP3蛋白抗原表位,根据 PRRSV FJ1402 ORF3基因序列设计一系列截短体引物并在引物的5′端引入EcoRⅠ酶切位点,3′端引入HindⅢ酶切位点,引物序列如表所示(表1),送南京金斯瑞公司合成。以重组质粒 pET-28a-tGP3为模板,采用PCR方法分别扩增出9个GP3蛋白基因截短体,将截短体和pET-32a载体双酶切后进行连接,构建9 个GP3蛋白截短体重组质粒。

表1 GP3蛋白截短体 PCR 引物序列
1.3 重组蛋白诱导表达及纯化
将重组质粒pET-28a-tGP3转化Rosetta感受态细胞,挑取单菌落,37 ℃培养,测定菌液 OD600 nm为0.6~0.8时加入终浓度为1 mmol·L-1的IPTG,继续培养6 h。收集菌体超声破碎,分离上清和沉淀,SDS-PAGE 鉴定蛋白表达形式,采用镍柱亲和层析法纯化目的蛋白,纯化GP3蛋白用超滤管进行浓缩后测定蛋白浓度,Western blot 试验鉴定GP3蛋白与PRRSV 阳性猪血清的反应性。
1.4 PRRSV GP3 蛋白单克隆抗体的制备
1.4.1 小鼠免疫 按照文献报道[13],将重组GP3蛋白与等体积ISA206佐剂混合,充分乳化后皮下注射小鼠,60 μg·只-1,ELISA抗体效价≥1∶10 000时,通过腹腔注射50 μg重组GP3蛋白进行冲击免疫。3 d后采集脾,进行细胞融合。
1.4.2 单克隆抗体制备 按照文献报道[13-14],取冲击免疫小鼠脾细胞和SP2/0细胞按照7∶1比例混合,加入 PEG1450进行融合。将融合细胞铺于含有2% HAT选择培养基的96孔细胞板中,培养6 d左右全换液,间隔24~36 h 取杂交瘤细胞上清,用间接 ELISA 进行检测。筛选到的阳性杂交瘤细胞需要进行2~3次亚克隆,直至单克隆阳性率为100%。将稳定分泌抗体的杂交瘤细胞扩大培养至T25细胞瓶,冻存后于液氮中保存。
1.4.3 间接 ELISA 筛选阳性杂交瘤细胞 用重组GP3蛋白作为包被抗原,杂交瘤细胞上清作为一抗,同时设立阳性对照(免疫小鼠血清 1∶100 稀释)和阴性对照(SP2/0 细胞上清);羊抗鼠IgG(H+L)-HRP孵育二抗;TMB显色后加入终止液,酶标仪检测OD450 nm时的读数,P/N≥2.1判定为阳性。
1.4.4 腹水制备 按照文献报道[15],取8周龄雌性 BALB/c小鼠,腹腔注射500 μL无菌液体石蜡。7 d后,腹腔注射3×106~4×106个杂交瘤细胞。7 d后观察,小鼠腹部胀大并触之有波动感时用头皮针收集腹水,安乐死。收集的腹水4 000g离心10 min,取上清,分装后,于-80 ℃保存备用。
(1)该水源地历年变化分析得出氨氮、总磷等水质指标有逐年好转的趋势,但是COD存在一定的水质恶化风险。
1.5 单克隆抗体生物学特性鉴定
1.5.1 单克隆抗体细胞株的稳定性试验 将阳性杂交瘤细胞株连续传代,取 5、10、15代细胞上清,间接ELISA检测抗体效价和抗体分泌稳定性。
1.5.2 单克隆抗体亚型鉴定 按照单克隆抗体亚型鉴定说明,将单抗细胞上清按照1∶200 稀释,将待测样品加入板条中,每孔50 μL;加入1×山羊抗小鼠 IgA+IgM+IgG-HRP,每孔50 μL,混匀;室温孵育1 h;弃去孔内液体,PBST 洗板3次;加入显色液,室温避光显色10~20 min,加入终止液,各孔的OD450 nm值最高的孔即为相应的单抗亚型。
1.5.3 单克隆抗体的特异性反应鉴定 以纯化的PRRSV FJ1402、杆状病毒系统和原核系统表达的重组GP3蛋白为抗原,同时设置相应阴性对照,进行SDS-PAGE 和Western blot。将蛋白转印至NC膜上,杂交瘤细胞上清作为一抗,羊抗鼠IgG(H+L)-HRP孵育二抗,加入显色液后判定单抗特异反应性。
1.5.4 间接免疫荧光试验反应性鉴定 PRRSV接种Marc 145细胞,37 ℃培养48 h,加入4%多聚甲醛于37 ℃固定15 min,加入0.1%Triton X-100于室温作用25 min,PBS洗涤后加入单抗37 ℃作用1 h,洗涤后加入FITC-羊抗鼠 IgG,37 ℃作用1 h,洗涤后于荧光倒置显微镜下观察。
1.6 单克隆抗体表位鉴定
1.6.1 Western blot 将GP3蛋白基因截短体重组质粒转化至表达菌种Rosetta,IPTG 诱导表达,SDS-PAGE 后将蛋白转印至NC膜上,以单抗腹水为一抗,采用Western blot鉴定单抗识别的抗原表位。
1.6.2 抗原表位合成多肽间接ELISA 根据上述鉴定的抗原表位,合成多肽,以50 μg·mL-1剂量包被酶标板,加入GP3蛋白单克隆抗体,以猪抗PRRSV血清抗体(G2#)和PBS分别作为阳性和阴性对照,37 ℃作用1 h后,加入HRP-SPA,37 ℃作用1 h。TMB显色后加入终止液,酶标仪检测OD450 nm,P/N≥2.1判定为阳性。
1.7 GP3 单抗抗原表位氨基酸位点对比分析
选取17株不同谱系的PRRSV-2中国流行毒株及参考毒株序列(BB0907、SY0608、HUN4、JXA1、WUH4、GD1404、CH-1a、IngelvacATP、QYYZ、GM2、VR2332、BJ-4、S1、MN184C、NADC31、NADC30、FJ1402),利用Jalview软件对比分析4种抗原表位序列在各毒株中的保守性。
1.8 GP3 单抗与不同毒株反应性
选取针对不同抗原表位的单克隆抗体,采用IFA和Western blot测定其与高致病性PRRSV毒株BB0907和经典PRRSV毒株S1的特异反应性。
2 结 果
2.1 GP3 蛋白基因表达与重组蛋白的制备

A.重组蛋白表达;B.包涵体纯化;C.Western blot鉴定。M.蛋白质相对分子质量标准;1.诱导表达全菌;2.诱导表达上清;3.诱导表达沉淀;4.未诱导表达全菌;5.空载体诱导全菌;6.纯化的重组 GP3 蛋白
2.2 单克隆抗体制备及效价测定
小鼠三免后10 d,间接ELISA测定抗体效价均≥1∶10 000。取效价最高的小鼠冲击免疫,3 d后取脾进行细胞融合,通过间接 ELISA 筛选阳性杂交瘤细胞并进行亚克隆,最终获得7株分泌 PRRSV GP3 蛋白抗体的杂交瘤细胞,分别命名为2E12、4H9、5B2、6B3、6G7、9F3 和 9H5。杂交瘤细胞连续传代至15代,每隔5代取细胞上清检测抗体效价,结果显示效价基本稳定。用7株杂交瘤细胞分别制备小鼠腹水并测定其效价,结果如表2所示,7株腹水单抗(mAbs)ELISA效价为1∶(64 000~2 048 000)。

表2 杂交瘤细胞上清和小鼠腹水抗体效价的测定
2.3 单克隆抗体亚型鉴定
2E12、5B2单抗的重链属于IgG2a亚类,4H9、6B3、6G7、9F3 和 9H5单抗的重链属于 IgG2b 亚类,7株单抗的轻链类型均为 Kappa 型(表3)。

表3 单克隆抗体亚型鉴定(OD450 nm)
2.4 单克隆抗体Western blot特异性鉴定
7 株mAbs均能和PRRSV FJ1402、杆状病毒系统和原核系统表达的重组GP3蛋白发生特异性反应,不与 Marc 145 细胞、sf9 细胞和 pET-28a空载体发生反应(图2)。

A.2E12;B.4H9;C.5B2;D.6B3;E.6G7;F.9F3;G.9H5;M.蛋白质相对分子质量标准;1.纯化的PRRSV;2.Marc-145细胞对照;3.杆状系统表达 GP3 蛋白;4.sf9细胞对照;5.重组GP3蛋白;6.空载诱导全菌蛋白
2.5 单克隆抗体IFA特异性鉴定
图3结果显示,7 株mAbs均能和PRRSV FJ1402株感染的Marc 145细胞发生特异性的荧光,其中5B2和9H5荧光较强;6B3和9F3荧光较弱。

A.2E12;B.4H9;C.5B2;D.6B3;E.6G7;F.9F3;G.9H5;H.PRRSV N单抗;I.SP2/0上清
2.6 单克隆抗体的表位鉴定
本试验构建了一系列GP3蛋白截短体(图4A),Western blot结果显示(图4B),9个GP3蛋白基因截短体重组质粒原核表达蛋白均与His鼠单抗反应,说明重组质粒表达正确。7株单抗中,4H9、5B2、6G7、9H5识别55PLCPTRQAAAEILE68;9F3识别69PGKSFWCRI77;6B3识别78GHDRCSESDH87;2E12识别88DELGFMVPPGLSS100。

A.GP3 蛋白截短体;B.单抗识别区域
抗原表位合成多肽ELISA鉴定结果见表4,除9F3外,6株单克隆抗体识别的抗原表位多肽与Western blot结果一致,同时猪抗PRRSV血清抗体(G2#)和55-68 aa、78-87 aa和88-100 aa多肽反应为阳性。

表4 单克隆抗体表位ELISA鉴定
2.7 GP3蛋白抗原表位氨基酸位点对比分析
采用Jalview软件对比分析PRRSV-2四个主要流行谱系(Lineage)毒株GP3蛋白氨基酸序列,结果见图5,55PLCPTRQAAAEILE68和78GHDRCSESDH87保守性较差;而69PGKSFWCRI77在Lineage 5内具有较好的保守性;88DELGFMVPPGLSS100在Lineage 3和Lineage 5之间具有较好的保守性。
2.8 GP3蛋白与不同基因亚型PRRSV的反应性鉴定
采用针对4个不同抗原表位的单抗4H9、9F3、6B3和2E12,IFA和Western blot结果如图6和7所示,2E12与FJ1402、BB0907、S1毒株反应。4H9、9F3和6B3与FJ1402毒株反应,但不和BB0907、S1毒株反应。
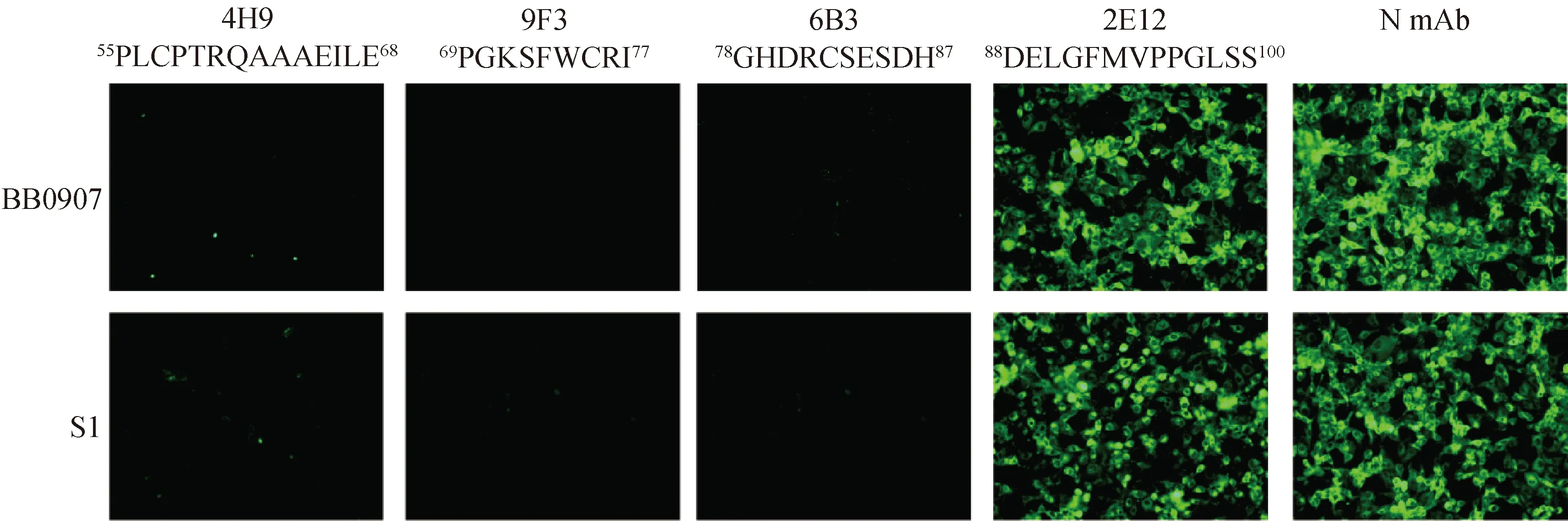
图6 GP3蛋白单克隆抗体与不同亚型PRRSV的IFA反应特性鉴定 (200×)

A.4H9;B.9F3;C.6B3;D.2E12。M.蛋白质相对分子质量标准;1.FJ1402;2.BB0907;3.S1;4.Marc-145细胞对照
3 讨 论
PRRSV存在基因变异和免疫抑制,商品化疫苗免疫保护效力有限[16-18]。研究PRRSV蛋白抗原特性对安全高效疫苗研究具有重要意义。PRRSV-1和PRRSV-2 GP3蛋白氨基酸序列相似性为 54%~60%,PRRSV-2不同基因谱系毒株 GP3蛋白基因保守性较差,可变区域主要位于蛋白N端[19]。本研究采用原核表达系统制备获得GP3重组蛋白,研制出7 株 GP3蛋白单抗,其中2E12单抗与 FJ1402、BB0907和S1毒株存在IFA和Western blot反应特性,4H9、9F3和6B3只与FJ1402毒株反应,而不与BB0907和S1毒株反应,5B2和 6G7与FJ1402毒株感染细胞IFA荧光效果最佳,为PRRSV不同谱系毒株鉴别和GP3蛋白功能研究提供了有用工具。
PRRSV病毒粒子GP3蛋白含量低于 GP5、M 和N 蛋白,但具有较好免疫原性[20]。PRRSV GP3、M和N 蛋白免疫原性水平最高,且 GP3 蛋白与中和抗体产生有关[21]。GP3蛋白有7个糖基化位点,预测蛋白大小27~29 ku,明显小于SDS-PAGE电泳显示的蛋白大小(41~50 ku)[8]。本研究Western blot 结果显示,7 株单抗与PRRSV感染细胞之间除了在40 ku左右有明显反应条带外,在其它位置存在数条反应条带,这可能与GP3 蛋白糖基化有关。此外,由于杆状病毒系统表达的GP3蛋白为第28―203 aa截短体,Western blot结果显示该重组蛋白反应条带分子量明显小于原核系统表达的重组GP3及PRRSV抗原。
病毒蛋白抗原表位解析对明确蛋白结构和抗原特性有重要作用。Wang等[22]基于肽芯片技术鉴定出PRRSV GP3蛋白抗原活性区域之一位于其N端51-106 aa。张阳等[23]通过GP3 蛋白重叠多肽合成,筛选出两个B细胞表位55PLCPTRQAAAEILEPGKS72和82CSENDHDELGFMVPPGLS99。Liang等[24]将GP3蛋白免疫显性肽段55PLCPTRQAAAEILEPGKS72与BSA 偶联制备免疫原,制备出GP3蛋白单抗1E5,证明其可与PRRSV高致病毒株反应,而与经典毒株和类NADC30毒株不能反应。Chen等[25]通过36株PRRSV-2分离株序列比对,发现GP3抗原表位87HDELGFMV94保守性良好,而59TRQAAAEILE68表位区域在其它低毒力毒株之间至少存在1个氨基酸差异。此外,也有报道GP3蛋白不同抗原表位存在部分重叠,如抗原表位59TRQAAAEILE68与两个表位61QAARQRLEPGRN72和67YEPGRSLW74重叠[25]。Wang等[26]利用单克隆抗体鉴定出GP3蛋白的B细胞线性表位69PGKSFWCR76,但该表位不能被阳性血清识别,提示该表位不能在猪体内诱导抗体。本研究通过GP3蛋白基因截短体构建表达,采用研制的单克隆抗体,鉴定出4个B细胞线性表位,分别为55PLCPTRQAAAEILE68、69PGKSFWCRI77、78GHDRCSESDH87和88DELGFMVPPGLSS100,与文献报道类似,但4个表位存在连续性,这可能与蛋白基因截短体构策略有关,这些抗原表位可能还需要更加精准定位。本研究抗原表位合成肽ELISA结果显示,5B2、6G7、6B3和2E12的检测结果均接近于临界值,9F3的检测结果为阴性,这可能与单抗结合能力和稀释度、合成肽抗原性和包被浓度及抗原表位免疫原性弱等因素有关。本研究采用针对4个不同抗原表位的单抗4H9、9F3、6B3和2E12,采用IFA和Western blot检测其与PRRSV 3个谱系毒株反应特性,结果均显示,2E12与FJ1402、BB0907和S1三个谱系毒株均可发生特异反应,4H9、9F3和6B3只与FJ1402毒株反应,而不与BB0907和S1毒株反应,与这三个谱系毒株88-100 aa、55-68 aa、69-77 aa和78-87 aa抗原表位氨基酸比对分析结果一致,但值得注意的是,9F3单抗识别的69-77 aa抗原表位在FJ1402和BB0907毒株之间没有差异,但是9F3单抗IFA和Western blot不能检出PRRSV BB0907和S1毒株感染细胞,提示PRRSV BB0907和S1毒株感染细胞GP3蛋白存在方式与PRRSV FJ1402的存在差异,需要进一步研究证实。
4 结 论
本研究研制出7株PRRSV FJ1402毒株GP3单克隆抗体,鉴定出GP3蛋白4个抗原表位,分别位于55PLCPTRQAAAEILE68、69PGKSFWCRI77、78GHDRCSESDH87和88DELGFMVPPGLSS100。2E12单抗可识别FJ1402、BB0907和S1三个谱系毒株,但4H9、9F3和6B3单抗只与FJ1402毒株反应,可用于PRRSV 检测和GP3蛋白功能研究。
猜你喜欢
遵义医科大学学报(2023年4期)2023-05-05
中国现代医药杂志(2020年10期)2020-12-14
中国免疫学杂志(2017年1期)2017-01-17
现代检验医学杂志(2016年3期)2016-11-15
畜牧兽医学报(2016年5期)2016-07-16
畜牧兽医学报(2015年3期)2015-07-05
医学研究杂志(2015年6期)2015-07-01
医学研究杂志(2015年3期)2015-06-10
特产研究(2015年1期)2015-04-12
生殖医学杂志(2013年5期)2013-03-11
