
祁连山东段高寒草甸高原鼢鼠和白牦牛营养生态位及其种间关系
2023-09-25 08:48花立民王志鹏唐燕青唐燕花邹小玉杨志财
青海畜牧兽医杂志 2023年2期
周 睿,花立民,王志鹏,唐燕青,唐燕花,邹小玉,杨志财
(1.甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展中心,甘肃 兰州730070;2.青海大学畜牧兽医科学院,省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016;3. 青海省河南蒙古族自治县托叶玛乡农牧业技术服务站,青海 黄南 811500;4.青海省河南蒙古族自治县生态有机畜牧业技术服务中心,青海 黄南 811500;5.甘肃省甘南自治州玛曲县草原工作站,甘肃 玛曲 747300)
动物营养生态位是指动物能够实际和潜在占据、利用或适应的食物资源,不但表现动物自身的营养需求特征,而且也反映其在生态系统中所处的功能和营养层[1]。因此,研究动物营养生态位对掌握生态系统中动物的资源利用状况意义重大[2]。当前,动物营养生态位的研究主要集中在食性分析方面。这是因为食物的组成和分布不仅是动物赖以生存和繁衍的物质基础,也是了解动物与环境的相互关系,掌握其食物来源和营养级位置的前提和基础[3,4]。动物对食物的选择受其生长、发育、竞争以及季节性食物资源变化等诸多因素影响[5-7]。因此,研究动物食性,不仅要关注其在短期内的食物选择,同时也需要探讨动物在整个生长阶段的食性特征及其变化,从而更好地认识和理解该物种在生态系统中的营养位置及其在长期演化过程中的适应策略。
高原鼢鼠是高寒草甸动物群落中的关键种[8],其通过采食、挖掘和排泄等活动参与高寒草地生态系统的能量流动和物质循环,对维持高寒草地生态系统的稳定具有重要作用[9,10]。当前,将高原鼢鼠定义为害鼠的主要原因,除了推土造丘造成鼠丘密布和裸地增加,导致草地生态功能下降外,更重要的原因是高原鼢鼠采食牧草,与放牧家畜争草损害了牧民经济利益,因而多年来处于被灭杀控制状态[11]。但有研究发现,高寒草地有毒植物的蔓延与高原鼢鼠种群密度存在显著负相关关系[13],且高原鼢鼠对杂类草中蔷薇科、毛茛科和龙胆科植物的采食显著高于禾本科植物[14]。才让吉等[15]研究发现,放牧家畜和高原鼢鼠的食性重叠度较低。但这些研究多局限于高原鼢鼠和家畜短期内的食谱组成、食量以及不同食物在食谱中的占比等方面,不能客观解释“与畜争食”的具体内涵。因此,要想对鼠类动物“与畜争食”的危害程度有一个客观准确的认识,必须在不同时间尺度下研究放牧家畜和草原鼠类的食性特征,并对其食物资源生态位的分异或重叠情况作出科学的分析和判断。
纵观国内外现代动物食性研究,主要还是采用直接观察法、粪便分析法和胃内容物分析法等传统方法[16],而传统的动物食性分析方法存在着诸如无法反映动物的长期摄食习惯,食物被咀嚼和消化后难以辨认,小型动物的胃内容物难以鉴别,以及室内喂食存在差异等缺陷[17,18],且种群间营养生态位关系是物种长期进化后的现象,短期食性可能无法反映物种间在时间尺度下的竞争或分化关系。稳定同位素分析(stable isotope analysis, SIA)技术作为一种成熟可行的技术,已成为动物食性研究的重要手段[19]。由于动物对食物中稳定同位素的同化过程需要一定的时间,且不同组织对稳定同位素的同化能力和过程又有差异[20],因此,根据动物不同组织中稳定性碳、氮同位素含量特征,可以推断出动物在不同时间段内的食性信息。利用稳定同位素技术研究生态系统中动植物的互作关系,不仅能够比较准确地反映动物在栖息地中的食物来源,而且能够揭示种间关系,这对于明确草地生态系统中草—畜—鼠之间的关系意义重大。
本研究通过对祁连山东段高寒草甸同域分布的高原鼢鼠和白牦牛的肌肉组织、毛发和骨骼组织样品进行稳定同位素(δ13C和δ15N)分析,研究高原鼢鼠和白牦牛的食性特征,进而明晰白牦牛和高原鼢鼠的营养生态位差异,揭示同域分布下高原鼢鼠和白牦牛的食物竞争关系,为指导草地放牧管理和科学防控鼠害提供理论和技术支撑。
1 材料与方法
1.1 试验地概况
本试验在甘肃省天祝县爪喜秀龙乡的高寒草甸区进行,该地区海拔为2 700~3 100 m,地理坐标为N 37°11′, E 102°29′。昼夜温差较大,空气稀薄,太阳辐射强,气候寒冷潮湿,年均降水量416 mm,主要集中在7~9月,年均蒸发量为1 592 mm,无绝对无霜期,仅分冷、热两季。年均气温0.1℃,7月和1月气温分别为12.7℃~18.3℃,≥0℃的年积温120~140 d。试验区的放牧动物为白牦牛(放牧率:4.04 sheep/hm2),高原鼢鼠是主要的植食性啮齿动物(种群密度:42.48 mounds/ hm2)。
1.2 样本采集
1.2.1高原鼢鼠样本采集 2015年9月从试验区捕捉高原鼢鼠10只,采集鼢鼠右侧后腿胫骨上附着的全部肌肉,封存在离心管中;用剪刀将背部近尾端的一小撮毛发紧贴表皮剪下,封存在离心管中;采集鼢鼠右侧后腿胫骨,封存在离心管中。将以上组织置于-20℃冰箱中冷冻保存。
1.2.2白牦牛样本采集 2015年9月从试验区随机选取6头白牦牛,对其进行编号,宰杀后在每头白牦牛的后臀部取500 g肌肉;在每头白牦牛的尾部紧贴皮层剪取1 cm2牛尾毛;取每头白牦牛部分腿骨。将以上组织装入自封袋置于-20℃冰箱中冷冻保存。
1.3 样本预处理
所有样本均置于65℃烘箱中烘干48 h以保证完全干燥,分别装于信封中并作标记。用玛瑙研钵分别研磨鼢鼠血液、肌肉、骨骼样本,标记编号备用。高原鼢鼠和牦牛毛发的处理参考郭波莉等[21]的处理方法。肌肉样本的处理参考易现峰等[22]有关肌肉样品的干燥方法及Schmidt等[23]对牛肉样品的脱脂方法。骨骼样本的预处理采用常规盐析法提取骨胶原。
1.4 稳定同位素测定
使用Picarro G2201碳同位素分析仪对预处理后的样品进行稳定碳同位素值(δ13C)的测定,测定精度≤±0.1‰;使用 Themomat 253氮同位素分析仪对样品进行稳定氮同位素值(δ15N)的测定,测定精度≤±0.2‰。同位素比值 ( δ值) 计算公式如下:
δ=[(R样品/R标准)-1]×1 000‰
式中,δ为13C值或者15N值;R为13C值和12C值或者15N值和14N值的比值;R样品为样品的R值;R标准为标准样品的R值。为保证试验结果的精度和准确度,每10个样品中放入3个标准品。
1.5 数据分析
采用R语言中的SIBER程序包对所测得的动物肌肉、毛发以及骨骼组织的稳定碳、氮同位素比值(δ值)进行分析,使用R语言中贝叶斯同位素混合模型(Bayesian isotope mixing models),δ13C值为横坐标,δ15N值为纵坐标,通过双变量协方差矩阵绘制出贝叶斯标准椭圆(Bayesian standard ellipse)计算其面积,对应生态位宽度,实现啮齿动物营养生态位宽度与重叠的定量分析[24]。考虑到 TA 可能因个别数据产生较大偏差,使用贝叶斯标准椭圆面积(SEA)可以减少样本误差,若样本数小于 30,则比较标准椭圆校正后的面积(SEAC)[25]。采用成组数据t检验比较高原鼢鼠与牦牛种内和种间以及潜在食源的δ13C和δ15N的差异,显著水平P<0.05。数据以平均值±标准误表示。
2 结果与分析
2.1 高原鼢鼠和白牦牛不同组织的稳定15N 和13C同位素特征
由表1可知,2种草食动物机体组织间的稳定15N和13C同位素差异均显著(P<0.05),高原鼢鼠各组织中,δ15N值从大到小的排序为肌肉(4.83±0.42)、骨胶原(4.33±0.7)、毛发(3.84±0.22),而δ13C值从大到小的排序为毛发(-26.37±0.31)、肌肉(-26.81±0.37)、骨胶原(-29.73±0.89);白牦牛各组织中,δ15N值从大到小的排序为毛发(3.76±0.14)、肌肉(3.23±0.6)、骨胶原(2.77±0.32),而δ13C值从大到小的排序为毛发(-24.07±0.22)、骨胶原(-24.31±0.63)、肌肉(-27.13±0.85)。这说明,高原鼢鼠食物源中的15N和13C稳定同位素在其肌肉中富集的时间最短,在毛发和骨骼中富集的时间最长;而白牦牛食物源中的15N和13C稳定同位素在其毛发中富集的时间最短,在肌肉和骨骼中富集的时间最长。

表1 高原鼢鼠和白牦牛碳、氮同位素比值
2.2 高原鼢鼠和白牦牛各组织营养结构的变化
利用贝叶斯标准椭圆对高原鼢鼠和白牦牛各组织的δ15N、δ13C进行分析,并计算生物群落生态指标参考值。结果显示,白牦牛各组织中,肌肉组织的营养层次(NR)、摄食来源的多样性水平(CR)、δ13C/δ15N围成的总面积(TA)和校正后的总面积(SEAC)最高,其次是骨骼组织,在毛发中这4种生态位指标最低;而在高原鼢鼠各组织中,骨骼组织的上述4种生态位指标最高,其次是肌肉组织,在毛发中最低(见表2)。
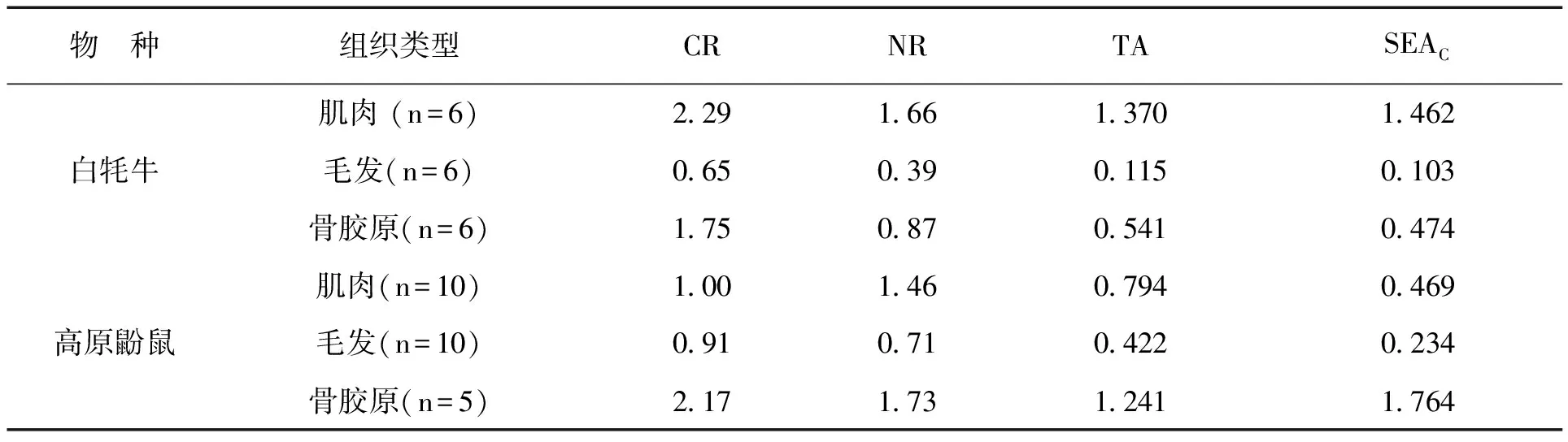
表2 高原鼢鼠和白牦牛各组织营养结构
2.3 高原鼢鼠和白牦牛不同组织营养生态位
以δ13C值为横坐标,δ15N值为纵坐标,拟合出2种食草动物不同组织营养生态位的贝叶斯标准椭圆(见图1)。结果显示,白牦牛各组织稳定同位素生态位宽度面积在肌肉组织中最大(1.462‰2),其次是骨胶原组织(0.469‰2),在毛发组织中最小(0.103‰2);高原鼢鼠各组织稳定同位素生态位宽度面积从大到小依次为骨胶原组织(1.764‰2)、肌肉组织(0.474‰2)和毛发组织(0.234‰2)。
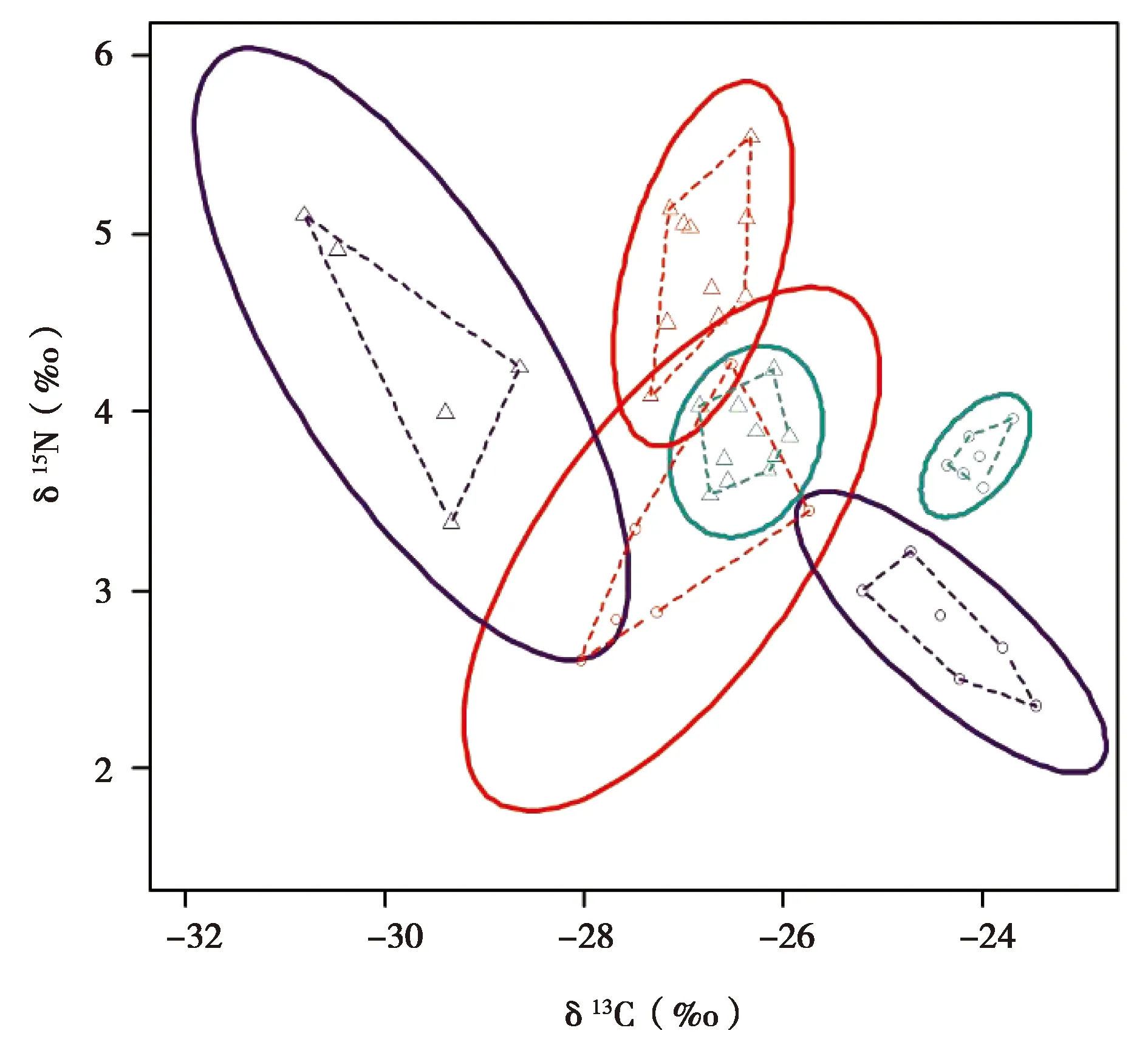
图1 高原鼢鼠和白牦牛不同组织稳定同位素营养生态位宽度
根据动物不同组织的碳、氮稳定同位素富集程度,动物的肌肉、毛发和骨胶原组织分别可以反映动物在2个月左右、半年左右和1年以上的食性信息。因此,本研究比较了高原鼢鼠和白牦牛在同一组织中的营养生态位重叠。结果表明,2种食草动物只有在肌肉组织中有生态位重叠,重叠面积为0.192‰2,分别占白牦牛和高原鼢鼠生态位面积的10.15%和31.32%,毛发和骨胶原组织中无生态位重叠,说明这2种食草动物的营养生态位仅在较短时间尺度上存在重叠,而在长时间尺度上存在明显分化现象。
3 讨论
3.1 高原鼢鼠和白牦牛的碳、氮稳定同位素比值
摄食作为消费者组织中同位素的外源性供应途径,直接影响δ13C和δ15N值的大小[26]。消费者组织中的δ13C含量可以指示其有机质和能量来源。本研究中,2种食草动物各组织δ13C值差异显著,这说明高原鼢鼠和白牦牛的不同组织中有机质来源差异较大,2种食草动物在食物网中可能存在营养分化现象[27]。动物自身的δ15N值可以指示其所在的营养级,本研究中2种食草动物不同组织的δ15N值差异均显著,说明二者并不在同一营养级中,且动物各组织的营养物质来源也差异较大。
根据动物组织内稳定同位素比值可以推断其在不同时间尺度下的食谱信息[28]。本研究发现,高原鼢鼠和白牦牛不同组织中的13C和15N富集程度变化趋势基本一致,δ13C在毛发和骨骼等反映长时间尺度食性信息的组织中大于肌肉组织,δ15N的变化规律则与δ13C恰好相反。可见,2种食草动物稳定同位素的富集与其组织器官完成的生理代谢过程基本一致,这与不同组织中营养物质的含量及其代谢率密切相关。食物中的15N在动物组织中富集的同时,其代谢产物通常以15N贫化的氮分泌物和排泄物排出体外。Robb等[29]对3种啮齿动物的研究表明,动物不同组织的δ15N值积累从爪子、毛发到血液逐渐增大,而δ13C值积累从爪子、毛发到血液逐渐减小。岳闯等[30]对布氏田鼠等4种荒漠啮齿动物不同组织13C和15N稳定同位素的研究也有一致的结果。
3.2 高原鼢鼠和白牦牛各组织营养结构的变化
动物组织中δ13C和δ15N值的变化反映了各组织同化的食源和营养多样性水平,CR和NR值越大,食源的生态位构成越复杂,多样性程度也越高[31]。高寒草甸栖息的食草动物食物来源丰富,摄食的选择性和栖息地植物群落的组成密切相关。因此,动物组织中CR和NR的变化往往反映了动物摄入的食物组成在时间尺度上的变化。根据以往的研究,动物的肌肉、毛发和骨胶原组织分别可以代表动物在2个月左右、半年和1整年的饮食信息[30]。高原鼢鼠骨胶原组织中的CR和NR值远高于肌肉和毛发组织,主要是由于在肌肉组织所代表的植物盛草期,高原鼢鼠的食物资源并不丰富,而在长时间尺度下,高原鼢鼠的食物资源逐渐丰富,造成CR和NR值在骨胶原中最高。白牦牛各组织中的CR和NR值则在肌肉组织所代表的盛草期最高,主要是因为盛草期牧草的多样性和营养最高,使得白牦牛肌肉组织中的CR和NR值最高。
3.3 高原鼢鼠和白牦牛的营养生态位及生态位重叠
生态位宽度和重叠是描述种间生态位关系最重要的指标。通常情况下,物种的生态位宽幅与其对栖息地、食物资源的利用及竞争能力呈正相关[32]。由于各组织样本数较少,因此本研究利用贝叶斯标准椭圆校正面积(SEAC)表征高原鼢鼠和白牦牛的营养生态位宽幅。本研究中,高原鼢鼠各组织稳定同位素生态位宽度面积从大到小依次为骨胶原(1.764‰2)、肌肉(0.474‰2)和毛发(0.234‰2);白牦牛各组织稳定同位素生态位宽度面积在肌肉组织(1.462‰2)中最大,其次是骨胶原(0.469‰2)和毛发(0.103‰2)。说明高原鼢鼠可能在长时间尺度下拥有更广泛的摄食来源,在短时间尺度下的摄食来源相对较窄,而白牦牛可能在短时间尺度下拥有更广泛的摄食来源,在长时间尺度下的摄食来源相对较窄。这可能与动物在不同时间段内食物资源的获取有关。由于动物在不同时间段内的食性信息可以通过其组织的稳定同位素值来获得[28],且根据样本采集时间(9月份)推测,高原鼢鼠和白牦牛肌肉、毛发和骨胶原组织分别可以代表其在2个月左右、半年和1整年的饮食信息。在肌肉组织代表的短期食性信息中,白牦牛可获得的食物资源丰富,因此生态位宽度面积也相对较大,而高原鼢鼠喜食的块根类植物在盛草期并不丰富,因此生态位宽度面积较小。
同域分布动物的营养生态位受种群密度、食物资源获取以及动物对微栖息地选择的影响。许多研究表明,动物会在食物缺乏或种内竞争加剧时倾向性地选择一些不喜食的植物,使种间营养生态位宽度和重叠度变大[33]。本研究发现,2种食草动物的营养生态位在肌肉组织表征的短时间尺度上存在重叠,而在毛发和骨胶原表征的长时间尺度上存在明显分化现象。由于肌肉组织稳定同位素能够反映动物2个月左右的食物来源,因此,在植物的生殖生长旺盛期(7月份),地上部分生长迅速,白牦牛食物资源最为丰富,而高原鼢鼠喜食的块根类植物的地下部分生长缓慢,迫使高原鼢鼠以适口性较差的植物为食。楚彬等[34]利用红外触发相机观察到高原鼢鼠地面活动和采食的现象,进一步说明在盛草期高原鼢鼠会通过调整营养生态位宽幅和自身对食物资源的偏好得以生存,从而与白牦牛产生一定的营养生态位重叠。另外,高原鼢鼠与白牦牛在时间和空间生态位上也存在明显的分化。对高原鼢鼠节律的研究表明,高原鼢鼠的活动高峰期出现在早上6∶00与晚上10∶00,而白牦牛的草地采食活动一般在白天进行[35]。从栖息环境上来看,高原鼢鼠虽然存在地面活动,但其大部分活动场所都是地下洞道系统。因此,2种动物存在明显的时间和空间生态位分化。当营养生态位重叠时,动物可能会通过对其他生态位的分化来削弱营养生态位所带来的竞争从而获得共存。本研究仅分析了特定时间段内同域分布动物间的营养生态位关系,而对不同种群密度下2种食草动物的营养生态位关系还亟待研究。
4 小结
综上所述,同域分布的高原鼢鼠和白牦牛在短时间尺度上存在重叠,而在长时间尺度上存在明显分化现象,这充分证明营养生态位的重叠是动物在特定阶段对微栖息地的生态适应对策。
猜你喜欢
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
力学学报(2021年10期)2021-12-02
能源工程(2021年1期)2021-04-13
兽医导刊(2020年3期)2020-12-30
苏州科技大学学报(自然科学版)(2020年1期)2020-04-13
防护林科技(2019年7期)2019-03-25
水利技术监督(2016年6期)2017-01-15
营销界·化妆品观察(2016年5期)2017-01-10
小资CHIC!ELEGANCE(2015年15期)2015-09-01
