
养殖网箱网纲和网衣水动力载荷研究
2023-10-26 11:18吴皓,刘强,范为
渔业现代化 2023年5期
吴 皓,刘 强,范 为
(1 南方海洋科学与工程广东省实验室(湛江),广东 湛江 524006;2 广船国际有限公司,广东 广州 511400)
根据联合国粮食和农业组织的数据统计,自20世纪80年代以来,水产养殖是过去40年来世界上增长最快的粮食生产方式,预计到2030年水产养殖将为人类提供超60%的鱼类消费[1]。随着水产养殖业不断的投资升级,渔业养殖装备从近海逐步走向深远海,从传统的养殖设施逐步向工业化渔业养殖装备转型升级。作为防止鱼类逃逸的唯一屏障,渔网承担着最为关键的角色[2]。
历年来相关领域的学者和专家对网衣水动力进行了大量的理论和试验研究。Xu等[3]阐述了3种网衣与流体相互作用的数值模拟方法,包括有限元法、计算流体力学(CFD)法以及有限元与CFD相结合的流固耦合法。有限元法涉及网衣的等效模型,包括三角单元模型、杆单元模型、质量弹簧模型以及集中质量模型[4-10]。CFD方法应用多孔介质模型模拟网衣,通过求解Navier-Stokes方程计算网衣水动力[11-14]。Cheng等[15]指出计算网衣水动力载荷的5种Morison模型和6种Screen模型。网衣Morison模型将网线和网结分别视为彼此独立的圆柱形细长体和球体,网衣Screen模型则是将网衣看作是一系列子网片的集合。Magnus等[16]应用Morison模型和Screen模型对两种典型的方形和圆形重力式网箱进行有限元计算,比较网箱在纯流中的变形、养殖体积与拖曳力。曹宇等[17]应用Morison模型研究重力式网箱系统联合浮式养殖平台运动响应的受力分布及变形。苗玉基等[18]采用Screen模型模拟网衣在规则波作用下的波浪载荷,与水槽试验结果吻合较好。
目前对网衣的研究大多关注柔性网箱网衣的受力和变形等,但关于网衣和纲绳耦合的研究还较少。深远海养殖网箱一般由刚性框架和网纲网衣装配而成的养殖网构成一个封闭的养殖空间。网纲一般由竖纤维绳或横竖交错布置的粗纤维绳连接到网箱周边的刚性结构上,承担着自身及网衣的载荷,能够抵制网衣的过大变形从而有效维持养殖体积。
为研究此类网箱养殖网变形与网纲受力,提出一种结合Morison模型和Screen模型计算网纲和网衣水动力载荷以及网纲结构张力的有限元方法。该方法将网纲和网衣模拟成杆单元,通过建立网纲和网衣耦合的有限元模型,求解平衡内外力的控制微分方程组获得耦合模型的张力和变形。通过研究网纲在纯流及波流联合作用下的张力和面法向偏移特性,为深远海网箱工程的网纲网衣的选型与配合、强度校核及布置方案的优化等打下研究基础。
1 网纲和网衣模型
本养殖装备为可移动柱稳式养殖网箱,主要由浮箱、立柱、撑杆及养殖网等组成。养殖网由网衣和网纲构成,网衣目脚长度l为50 mm,网线直径dw为3.5 mm,考虑海洋生物等附着的密实度Sn为0.176[19]。网纲直径为22 mm,最小破断力为3.88×103kN,动刚度为2.17×103kN/m。每面网衣均装配在纵横交错分布的竖纲和横纲上并与网纲缝合连接,初步方案的竖纲间距为1.3 m,横纲间距为1.5 m。网衣和网纲的材质均为超高分子量聚乙烯纤维。图1为网箱整网模型示意图。
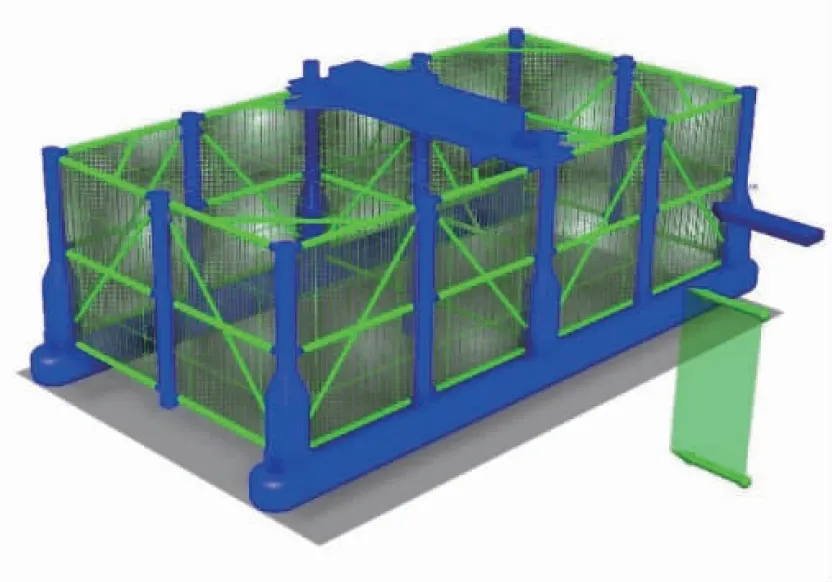
图1 某可移动柱稳式养殖网箱的养殖网模型示意图
2 计算方法
2.1 有限元模型
采用SIMA的RIFLEX非线性有限元求解程序对网纲和网衣相结合的耦合模型进行数值计算。耦合模型采用杆单元模拟网衣并基于Screen模型计算网衣水动力;采用Morison方程计算网纲水动力。从图1可以看到,网箱由多面在结构尺寸等参数上相同的养殖网组成,取相邻立柱间的养殖网为研究对象,构建养殖网网纲和网衣耦合的有限元模型。对象模型如图2所示。

图2 养殖网分析模型
固定边界为左右两边的立柱和上中下的横撑,中间的横撑将养殖网分隔成上下独立的养殖网A和B。模型中垂向和横向的网纲等构件分别以“V”和“H”加编号予以表示。有限元模型应用杆单元对网纲和网衣进行等效模拟,并通过杆端节点共用的方式将网纲和网衣模型耦合,可进行时域耦合计算,见图3。

图3 养殖网网纲和网衣耦合的有限元模型
杆单元为连接两端节点的线单元,可用于计算轴向拉伸变形,但不发生弯曲和扭转变形[20]。其轴向张力为:
(1)
式中:N为单元轴向力;L为拉伸变形后的长度;L0为初始长度;E为弹性模量;A为横截面积。
对网纲和网衣耦合的有限元模型,时域计算的非线性动态平衡微分方程组为:
(2)
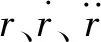
2.2 水动力模型
2.2.1 网衣水动力模型
Kristiansen等[4]将计算网衣受力的水动力模型分成两类,分别是Morison模型和Screen模型。网衣水动力可以表述为下式。

(3)
式中:ρ为水密度;▽为网衣浸水体积;Anet为网衣投影面积;u为流体速度;v为结构速度;经验水动力系数Ca和Cd分别为附加质量力系数和拖曳力系数,其取值取决于试验数据,与库卡数、雷诺数、网线粗糙度以及网衣密实度密切相关[4]。上述等式右边多项式前3项依次为入射力(或称Froud-Krylov力)、绕射力及辐射力,统称为惯性力;最后一项为粘性力。大量试验结果表明,网衣惯性力约为粘性力的dw/H倍(H为波高)[21]。例如,当dw=1 mm,H=1 m时,惯性力约为粘性力的0.1%。故网衣惯性力远小于粘性力,计算中可忽略不计。
网衣水动力既取决于实际的流速,也取决于网衣自身的水动力特性。水动力特性与网衣材料(金属或纤维)、网目形状(方形或菱形)、网目大小、网线直径、网线编织方法(捻线或编织线)以及网衣编织方法(无结或有结网)有关。光滑铜网在稳流中的阻力明显低于粗糙的尼龙网[22]。同环境条件下有结网的阻力比无结网的阻力要高约10%[23]。大量试验结果表明,网衣的水动力特性主要依赖于两个无因次变量,密实度Sn和雷诺数Re[22-25]。其中理想方形网的水动力主要取决于Sn。
Sn是网衣净投影面积与轮廓投影面积的比值。理想的无结方形网(见图4)Sn计算公式为:

图4 养殖网网衣示意图
(4)
式中:dw为网线直径,lD为相邻网线中心距。
Re与网衣的网线直径和流体速度有关。
(5)
式中:U为未受扰动的流体速度;v为流体运动粘性系数。网衣Re的典型范围通常为(100,10 000)。
Screen Model模型将网衣分解为一系列子网片,每一网片的受力可分解为沿网片法向和切向的两个分力或者水平拖曳力和垂向升力的两个分力,见图5。流体与网片相对速度Urel为:

图5 养殖网网衣受力示意图
Urel=γU∞+uw-us
(6)
式中:Urel为相对速度,U∞为流速,uw为水质点速度,us为结构速度。γ=1代表前方的网衣,γ=r代表后方的网衣,r为流速的折减系数。
网衣水动力FH为拖曳力FD和升力FL的两个矢量分力之和。
(7)
(8)
式中:ρw为流体密度,At为网片面积;Urel为相对速度;Cd和Cl分别为拖曳力系数和升力系数;id和il分别表示拖曳力和升力的单位矢量。Cd和Cl由试验得到,主要与Re、Sn及入射角θ有关。其中θ为波流入射方向与网片法向的夹角。本研究的Cd和Cl基于Løland提出的Screen模型计算,其适用密实度范围为(0.13,0.35)[26]。
CD(Sn,θ)=0.04+(-0.04+0.33Sn+
(9)
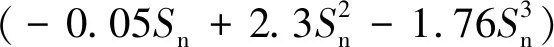
sin2θ
(10)
Løland公式并不包含雷诺数,Cd和Cl不随雷诺数而变化。一般来说,Cd和Cl高度依赖于Sn,并且当Re范围为(100,10 000)时,Re对网线阻力系数的变化几无影响,可忽略[15]。Cd和Cl随Sn的增加而增大,说明在其他条件相同的情况下,网衣Sn越大,Fh越大。Løland提出的Screen模型的Cd随θ的增大而衰减,并遵循余弦函数曲线的规律。Cl的变化遵循正弦函数sin2θ曲线的变化规律。当θ为零时,升力为零;当θ较小时(θ<30°),升力相对于拖曳力是小量,后者起主导作用。
2.2.2 网纲水动力模型
网纲水动力的计算基于Morison模型[27]。网纲由竖纲和横纲交错布置,网目尺寸相对于网纲直径要大得多,Sn对网纲阻力影响很小,所以适合采用Morison方程计算网纲水动力。
(11)
式中:dRope为网纲直径;LRope为网纲长度;ARope为横向截面面积;Cd和CA分别为拖曳力系数和附加质量力系数。在大多数情况下,拖曳力系数主要是Re的函数。式中的拖曳力项可分解为两部分:垂向拖曳力Fn和轴向拖曳力Fτ,见图6。
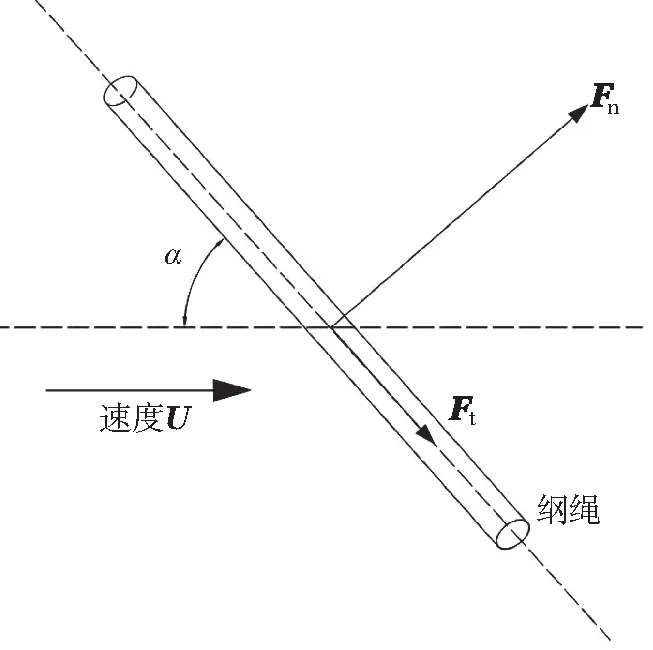
图6 养殖网网纲受力示意图
(12)
(13)
式中:un和uτ分别为垂向和轴向流体相对结构的速度矢量;Cn和Cτ为垂向和轴向拖曳力系数。一般来说,Cτ比Cn要小得多,因此忽略不计轴向拖曳力的模拟结果是可以接受的。
2.3 尾流
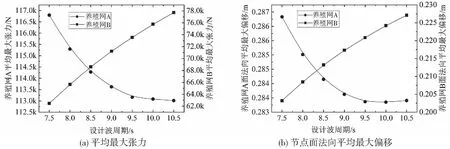
来流经过网衣后将产生尾流效应。由于流体的粘性作用将使尾流发生扰动,流速受到折减[28],下游网衣的受力也将有所减小,一般采用速度折减系数r(0 Udownstream=rUupstream (14) 式中:Uupstream为来流速度,Udownstream为尾流速度。折减系数r一般为Re,Sn及θ的函数,在实践中较常用的计算公式为r=1-0.46Cd,Cd为Screen模型中网衣的拖曳力系数。 网箱坐底吃水d为23.4 m,自存环境条件为:有义波高5 m,流速1.6 m/s。采用确定性的设计波方法,设计波波高H为有义波高的1.9倍,设计波周期范围为7.5~10.5 s,Ursell数最大值约为21.9,适用Stokes 5阶波波浪理论[29]。为研究本养殖网的网纲最大张力与面法向偏移,假定纯流、波流的入射角为0°。 纯流条件下当流速为1.6 m/s时,对养殖网A网纲单元的最大张力值进行统计,作垂线图如图7所示。 图7 养殖网A在纯流1.6 m/s下的最大张力响应统计图 可以看到,竖纲和横纲的最大张力都呈现中间高、两边低的走势,与应用多孔介质模型研究的网衣整体呈现中间受力大的特性相同[30]。由于竖纲的长度要小于横纲,相对伸长率大于横纲,故从图中可以看到竖纲的最大张力整体要大于横纲。统计养殖网A和B网纲单元的最大张力和节点面法向最大偏移并取平均值作图如图8所示。 图8 养殖网A和B随流速的平均最大张力和节点面法向平均最大偏移 图8a表明,随着流速的增加,平均最大张力与面法向最大偏移弱非线性增大。该特性类同于Magnus等[16]指出网箱阻力随流速的增加呈非线性增加的趋势。图8b表明,随着流速增加,网纲承力逐渐增加,拉伸变形也逐渐增大,这与对柔性网衣变形的有关研究得出的随流速增加而增大的特性一致[31]。由于养殖网A部分位于水面以上,图中可以看到A的平均最大张力与面法向最大偏移整体稍低于B。 取T=7.5 s的波流工况,统计养殖网A的网纲单元最大张力并作图9。垂线图表明竖纲和横纲最大张力整体都呈现中间高,两边低的走势。同一竖纲或横纲网纲单元沿轴向的最大张力呈现末端高,中间低的走势。由于养殖网A整体水深比B要浅,设计波波速更大,养殖网A网纲单元的最大张力统计值约为188 kN,比B大约88%,这种显著性差异说明当养殖网B采用与A同规格的网纲时其间距可相对布置的更宽。Qu等[32]对柔性网箱的研究同样表明水深的显著作用,体积剩余率随着水深增加而增加,单位体积的阻力变小。 图9 养殖网A流速1.6 m/s、波幅4.75 m、周期为7.5 s的最大张力响应统计图 通过箱线图10对周期T=7.5~10.5 s下的网纲单元最大张力进行统计性分析发现,养殖网A网纲单元最大张力的统计最大值、中位值和均值随着周期的增大呈现缓慢减小的趋势。箱体宽度变化不大,说明整体网纲单元最大张力的波动程度相当。养殖网B的统计最小值、最大值、中位值和均值随着周期的增大缓慢递增,与A走势相反。 图10 养殖网在流速1.6 m/s、波幅4.75 m下的最大张力统计性分析 这种不同走势的原因在于流速在不同水深处随周期变化的差异性。 从图11可以看到在自由液面附近,流速随设计波周期增大而减小;不同周期的设计波流速均随水深的增加而减小,但程度不同。周期越小,流速随水深的增加衰减越快。养殖网B所处水深的流速随周期增大而增加,故其最大张力随着周期的增加而增加。总体上,在设计波周期范围内的养殖网A网纲单元最大张力的统计最大值显著大于B。 图11 不同周期波速与水深的变化关系 图12表明了养殖网A和B的网纲单元平均最大张力和面法向平均最大偏移随设计波周期的变化。可以看到养殖网A的均值随周期的递增而向下凹;B则呈向上微凸的走势。 图12 养殖网A和B网纲单元平均最大张力和面法向平均最大偏移随周期的变化 提出一种结合计算Morison模型和Screen模型的有限元方法,网衣水动力的计算基于Screen模型,网纲水动力的计算基于Morison模型。通过建立网纲和网衣耦合的有限元模型,研究了有限元模型在纯流、波流联合作用下的张力与面法向偏移。对数值计算结果进行统计分析发现:纯流条件下养殖网A和B的网纲单元最大张力和节点面法向最大偏移的统计最小值、最大值、中位值和均值均随着流速的增加而递增,均值随流速的变化曲线呈弱非线性增加;波流联合作用下养殖网A和B的所有网纲单元最大张力和节点面法向最大偏移的统计特征存在显著差异,同等条件下A的网纲统计最大张力和平均最大张力均比B要大的多,水深的明显作用说明B的网纲间距可相对A布置的更大一些。A的网纲平均最大张力和节点面法向平均最大偏移随设计波周期的增加而非线性递减,B则呈现弱非线性递增的走势。通过网衣网纲的耦合分析方法以及张力与偏移的特性分析,将指导网衣网纲的选型、装配与布置设计得更加合理。例如,由2片独立养殖网分成4块甚至6块等,可有效减小张力与偏移;水深相对更深的养殖网网纲间距可相对布置得更大;单片养殖网网纲可布置成从中间向两边由密向疏等等,这都将对方案设计阶段的设计和优化具有重要的指导意义。3 结果与讨论
3.1 纯流条件下的张力与面法向偏移


3.2 波流联合作用下的张力与面法向偏移



4 结论
猜你喜欢
船舶与海洋工程(2023年3期)2023-07-14
渔业研究(2023年3期)2023-06-20
落叶果树(2021年6期)2021-02-12
中成药(2017年4期)2017-05-17
辽金历史与考古(2017年0期)2017-02-06
电子制作(2017年24期)2017-02-02
渔业致富指南(2016年12期)2016-11-11
湖南农业(2016年3期)2016-06-05
辽宁省博物馆馆刊(2016年0期)2016-05-17
中国酿造(2016年12期)2016-03-01
