
独脚金内酯抑制因子D53/SMXLs基因的研究进展
2023-12-20 07:01王雪菱王如月李际红王锦楠王冬月牛牧歌孙茂桐
农学学报 2023年10期
王雪菱,王如月,李际红,王锦楠,王冬月,牛牧歌,孙茂桐
(山东农业大学林学院/山东泰山森林生态系统国家定位观测研究站/黄河下游森林培育国家林业和草原局重点实验室,山东泰安 271018)
0 引言
植物激素是植物体自身合成的一类微量有机物,在植物生长的各个阶段起着重要作用。植物激素主要包括生长素、细胞分裂素、脱落酸、赤霉素、乙烯、独脚金内酯等[1-2]。独脚金内酯最初在寄生植物独脚金属的种子中发现并由此得名[3]。独脚金内酯能促进种子萌发,刺激丛枝菌根真菌与寄主植物根的共生,抑制植物分枝,于2008年被鉴定为一种新的植物激素[4-6]。
在水稻、拟南芥等模式植物里,对独角金内酯生物合成和信号转导的机制研究已较为透彻[7-10],在营养条件有限的情况下,植物根中合成的独脚金内酯能促进根毛和侧根生长,进而增加根部的营养物质吸收,同时独脚金内酯被运输到地上的部分可抑制侧芽或分枝的形成,从而降低植物对营养物质的需求[11-12]。随着在不同植物对独脚金内酯突变体的研究,其信号转导途径逐渐变得清晰。其中,负责独脚金内酯信号传导的蛋白大体分为3 类:α/β折叠型水解酶DWARF14(D14)/DECREASED APICAL DOMINANCE2(DAD2)/RAMOSUS3(RMS3),富亮氨酸重复序列F-box 蛋白DWARF3(D3)/MORE AXILLARY GROWTH2(MAX2)以及Clp 蛋白酶家族DWARF53(D53)/SUPPRESSOR OF MORE AXILLARY GROWTH2 LIKE I(SMXLs)[13-15]。在独脚金内酯信号转导过程中,D14和D3可以形成SCF复合体,SCF形成的开放小室可用来容纳独脚金内酯分子,并将其水解成共价连接的中间分子CLIM(Covalently Linked Inter Mediate Molecule),促进D53/SMXLs的Clp蛋白酶家族降解,从而调节植物的分枝[16-18]。Clp 蛋白酶家族的D53/SMXLs 蛋白作为独脚金内酯信号转导复合体的重要成员,在独角金内酯信号转导过程中发挥着重要的作用。
1 D53/SMXLs的发现
水稻独脚金内酯信号转导相关的基因已被广泛研究,而对水稻分枝的研究主要来源于两种类型的突变体,即moc1突变体和Dwarf系列突变体[19-20]。2013 年李家洋和万建民团队同时发现了水稻的短茎显性突变体Dwarf53(d53),该突变体表现出矮化多分蘖。对该突变体进一步研究表明,这是由于D53/SMXLs基因发生显性突变,突变后的蛋白不能被独脚金内酯类似物GR24 降解,因此导致该表型的出现。由此人们推论D53/SMXLs是参与独脚金内酯信号传导的关键基因[7-8]。随着基因组测序技术的不断发展,人们发现大部分植物都含有D53基因家族,拟南芥(Arabidopsis thaliana)有3 个同源基因,水稻(Oryza sativa L.)中有2个,玉米(Zea maysL.)中有1 个,杨树(Populus)中有3个,豌豆(Pisum sativumL.)中有3个[7,21-23]。随着研究的不断深入,D53基因被认为是一种参与独脚金内酯信号转导和调控植物分枝发育的抑制因子[7-8]。
2 D53/SMXLs的结构
D53/SMXLs 及其同源物与Clp 双AAA 结构域域腺苷三磷酸酶(ATPase)家族成员具有相似的二级结构,他们都参与蛋白质分解和蛋白质重塑并具备保守序列[7-8]。AAA 蛋白的功能特征是依赖三磷酸腺苷(ATP)的蛋白质分解和蛋白质复合物组装和分解活动,其中底物通过穿过中心孔从聚集体或复合物中分离出来[24-25]。许多AAA蛋白以DNA/蛋白复合物作为底物,具有转录抑制功能的双AAA 蛋白可直接或间接地与转录因子相互作用[26]。D53蛋白结构上含有EAR-2和EAR-3 保守结构,EAR-2 由结合典型TPD 结合位点1的c端LxLxL基序和结合之前未识别的TPD结合位点2 的n 端DNLIYLDL 基序组成[27]。进一步研究表明,这两个基序都参与抑制独脚金内酯信号,但EAR-2可能对分蘖数更重要,而EAR-3可能对株高更重要。拟南芥SMXL7 保守的c 端EAR 基序的突变也导致了独脚金内酯信号的部分缺失。EAR-2 可与TPD 蛋白沟结合抑制生长素和茉莉酸信号,或与TPD第二位点结合介导TPD 四聚体相互作用[7-8,27]。对水稻的研究表明,2个TPD结合位点的功能是相关的,证明EAR-2结合诱导TPD寡聚,此外,TPD可直接与组蛋白H3和H4以及核小体结合,TPD 的高水平组装与D53 的EAR-2结构结合,使TPD复合核小体之间的相互作用更加稳定[27]。如图1所示。
3 D53/SMXLs功能
3.1 D53/SMXLs在独脚金内酯信号传导中的作用
研究表明,D53/SMXLs基因的主要功能是抑制独角金内酯信号转导[8]。当独脚金内酯浓度足以激活独角金内酯信号通路时,D14 蛋白将独脚金内酯识别并水解为活性分子,D14 蛋白招募SCF(ASK1-CULLINF-BOX)复合体并与D3蛋白结合进一步形成SCFD14复合体,D3蛋白特异性识别并结合独脚金内酯信号抑制因子D53/SMXLs,形成D53-D14-SCFD3蛋白复合体,D53/SMXLs通过泛素结合酶E2的泛素化修饰,并最终被26S 蛋白酶体降解,诱导下游靶基因的表达[7-8,28](如图2)。LIU[21]的最新研究表明,水稻OsD53的同源基因ZmD53在玉米中通过独脚金内酯信号传导与ZmD14A 和ZmD14B 相互作用,从而影响下游基因的表达。
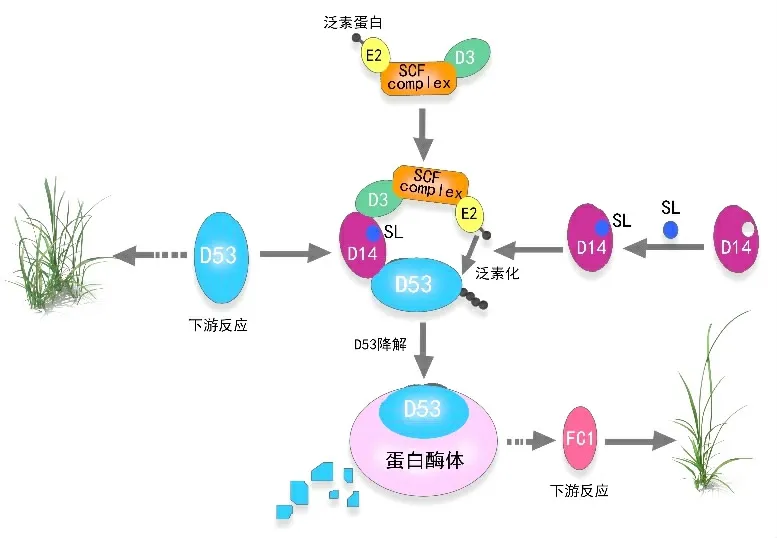
图2 独脚金内酯促进D14-SCFD3介导的D53/SMXLs降解的示意图模型
而在双子叶植物中,D53/SMXLs基因在独脚金内酯信号转导通路中发挥着同样的作用。拟南芥SMXL6、SMXL7和SMXL8作为抑制因子可调控叶片形态和植物分枝[9]。SMXL6/7/8作为MAX2(D3)下游的基因,经过泛素化过程后,通过26S 蛋白复合酶体将SMXL6/7/8降解,使独脚金内酯抑制腋芽萌发,而当独脚金内酯不存在时,则不会形成泛素化蛋白复合体,SMXL6/7/8可与TPL/TPR相结合,从而抑制独脚金内酯的下游基因(如BES1和SPL9/15蛋白)的转录,进而调控腋芽发育[29]。在豌豆中,D53/SMXLs基因家族共包含3个成员,分别为PsSMXL6、PsSMXL7和PsSMXL8,PsSMXL7作用于RMS3(D14)和RMS4(D3)的下游[23]。
杨树作为木本植物的模式植物,关于D53/SMXLs基因也有相关研究报道。杨树的PagD53基因在腋芽和相关节点中表达量较高[22]。进一步研究表明,PagD53含有ClpB以及ClpA蛋白家族结构域,受体蛋白PagD14和PagD53可相互作用,PagD53蛋白通过独脚金内酯的信号传递调节植物分枝发育[30]。然而PagD53在杨树中的下游作用基因,以及其引起的独脚金内酯在内的各种激素如生长素、细胞分裂素、赤霉素等的含量变化尚未明朗[30]。
已有研究表明,D53/SMXLs基因作为独脚金内酯信号转导的抑制因子,还可以此通路来调控其他激素。李家洋团队发现,D53/SMXLs基因通过诱导独脚金内酯进而拮抗细胞分裂素,降解OsCKX9酶基因的转录,促进细胞分裂素的积累[31]。近期的研究发现SMXL6,7,8 的EAR 基序抑制BRC1表达,降低脱落酸的含量[32]。其中,BRC1是抑制腋芽伸长的重要转录因子,它可使靶基因HB40表达,促进脱落酸的合成,进而减少植物分枝;而SMXL6的EAR基序可抑制BRC1基因表达,进而抑制脱落酸的合成,促进植物分枝[33]。当添加GR244DO激素时,可使SMXL6降解,从而释放BRC1,激活HB40的表达,提高脱落酸含量,抑制植物分枝[33]。
豌豆有PSAFB4/5 依赖的根尖反馈信号,该信号上调独脚金内酯生物合成基因的转录丰度[34],并受到独脚金内酯感知的负调控[35-36],这个依赖于RMS 的信号被提议为IAA[37]。Psafb4/5-1是生长素受体家族成员中的一个突变体[38]。Dun等[36,39]发现与野生型相比,Psafb4/5-1主要在植物的上部节点有所降低,Pssmxl7-1突变几乎完全恢复了Psafb4/5-1的茎分枝和高度,这表明Psafb4/5-1表型依赖于SMXL7,由此得出独脚金内酯通过PsSMXL7抑制IAA水平。
综上所述,单子叶和双子叶植物中D53/SMXLs基因在独脚金内酯信号转导通路中发挥着同样的作用,此外D53/SMXLs基因通过独脚金内酯信号传导调控ABA和IAA等激素,进而影响植物发育;在木本植物中D53/SMXLs基因在独脚金内酯信号转导通路的研究较少,其通过独脚金内酯影响其他激素调控尚未报道。
3.2 D53/SMXLs在分枝发育中的功能
D53/SMXLs基因作为独角金内酯信号转导通路的抑制因子,在独角金内酯下游靶基因调控中发挥着重要的作用[7-8]。TPL/TPR 是植物激素信号转导通路中抑制基因转录的共抑制因子(co-repressor)[40]。水稻OsD53 含有3 个典型的EAR 基序,它们与TPL(TOPLESS)/TPR(TPL-RELATED PROTEIN)蛋白相互作用[8]。水稻基因组中有3个TPL/TPR,OsD53可以与TPL2和TPL3相互作用。OsD53可与TPL/TPR蛋白形成转录抑制复合物,协同抑制靶基因在独脚金内酯信号通路下游的表达,使该信号通路下游的基因无反应[7]。进一步研究表明,该下游基因是转录因子IPA1,D53蛋白和TPL蛋白共同形成抑制因子可以结合转录因子IPA1 蛋白,抑制其转录激活,调控植物分枝[7,27,41]。IPA1,又称OsSPL14,是SQUAMOSA启动子结合蛋白样(SPL)家族的成员,属于植物特异性的转录因子[42]。IPA1编码SPL14转录因子,它的表达受miRNA156和miRNA529的调控[43],其中TB1被证明是IPA1的直接转录靶点[44]。IPA1能够直接结合到水稻蘖芽生长的负调控基因OsTB1(TEOSINTE BRANCHED1)的启动子(FC1)上,从而抑制水稻的分蘖[42,45]。SONG 等[40]检测了IPA1对独脚金内酯下游的潜在参与,他们发现D53/SMXLs在体内和体外都能与IPA1发生作用,进而抑制IPA1的转录因子活性。这些结果表明,独脚金内酯信号通路与MIR156/SPL调控模块在TB1/FC1/BRC1启动子处汇合,协同调控植物分枝[46]。FANG 等[47]研究表明,水稻D53与OsBZR1相互作用,这种抑制依赖于OsBZR1的DNA 直接结合,OsBZR1将D53招募到水稻芽中的FC1启动子上,进而调节水稻分枝。油菜素内酯(Brassinosteroids,BRs)通过促进水稻芽的生长而增强分蘖[47]。BRASSINAZOLE RESISTANT 1(BZR1)是BRs 下游转录因子[48]。在面包小麦中,水稻OsD53的同源基因TaD53可与TaSPL3和TaSPL17 蛋白相互作用,从而抑制了TaSPL3/TaSPL17介导的TaTB1表达的转录激活[49]。
HU 等[50]通过转录组学和遗传分析等实验表明BES1在独脚金内酯中调控芽的分枝。拟南芥中当独脚金内酯缺失时,SMXLs与磷酸化或未磷酸化的BES1相互作用,BES1与启动子结合,SMXLs的EAR基序招募TPR2,从而抑制BRC1的表达,增加分枝数量。当独脚金内酯存在时,SMXLs-BES1 复合物在感知独脚金内酯后被AtD14-MAX2降解,从而表达BRC1,抑制芽分枝[50]。TCP 转录因子BRC1 及其同源物被认为是通过协调不同的环境和发育线索来调控芽生长的关键开关[51-52]。最新研究发现,拟南芥的抑制因子SMXL6也是转录因子,通过与转录因子D14、MAX2相互作用来抑制转录,也可以作为转录因子直接与SMXL6/7/8启动子结合,形成负反馈环来维持SMXL6/7/8蛋白的稳态[33],当独脚金内酯存在时,受体D14感知并水解独脚金内酯生成活性激素,D14与SCFMAX2相互作用形成复合物,复合物招募SMXL6,促进其泛素化和降解,消除SMXL6 的表达抑制。新合成的SMXL6 直接与SMXL6、7、8的启动子结合,形成负反馈回路,维持SMXL6/7/8蛋白的稳态[33,53]。
在 玉米 中,IPA1的3 个同 源基 因UB2、UB3和TSH4在限制侧枝原基启动方面起着关键作用,从而影响玉米的分蘖和雄穗分枝数[54]。研究发现,ZmD53可以与UB3和TSH4相互作用,但与UB2不相互作用[21]。水稻OsD53可抑制IPA1的转录激活活性,导致下游基因FC1的表达降低,从而导致更多分蘖[44],而TB1是玉米中的FC1同源物[55],ZmD53通过抑制UB2/UB3/TSH4在TB1上的表达活性,促进分蘖形成,其中UB2、UB3和TSH4可相互作用,并且ZmD53直接调节TSH4的表达以形成正反馈回路[21]。
KERR等[23]发现豌豆PsSMXL7抑制RMS3(D14)下游的PsBRC1,通过独脚金内酯调控PsBRC1的转录,从而调节枝条分枝,BRC1蛋白是一种转录因子,作用于独脚金内酯下游,参与调控豌豆的分枝发育[56]。Pssmxl6和Pssmxl8对下游PsBRC1的调节途径还有待研究[23]。进一步研究表明,PsSMXL7 蛋白通过抑制BRANCHED1(PsBRC1)的表达促进芽的生长,防止PsBRC1通过PsNCED2等下游反应基因抑制芽的生长,独脚金内酯通过靶向PsSMXL7蛋白进行泛素化和降解来抑制芽的生长,PsSMXL7对PsSMXL7/8表达的负反馈调控导致独脚金内酯处理后PsSMXL7/8转录上调[23]。
3.3 D53/SMXLs在其他生长发育中的功能
氮(N)是粮食作物必不可少的主要营养元素,水稻通过改变其根系形态(如根系伸长)来响应NO3ˉ的施用。SUN等[57]研究发现,与NH4+处理相比,NO3ˉ处理后的水稻根系中独脚金内酯信号水平较高,D53 蛋白含量较低,进一步鉴定表明,SPL14的同源蛋白SPL17也与D53相互作用,其中PIN1b是IPA1(SPL14)的靶基因[58]。当NH4+存在时,D53与SPL14/17结合以抑制SPL14/17的转录活性并抑制根伸长,当NO3ˉ存在时,D14对独脚金内酯的感知导致D53通过蛋白酶体系统降解,从而停止抑制SPL14/17介导的PIN1b转录并导致水稻根伸长[57]。
在玉米中,IPA1的3个同源基因UB2、UB3和TSH4在限制侧枝原基启动方面起着关键作用,从而影响玉米的分蘖和流苏数[54]。玉米ZmD53是水稻D53的同源基因,它参与独脚金内酯信号转导通路。Liu等[21]通过酵母双杂交实验分析证明ZmD53可以与UB3和TSH4相互作用,但与UB2不相互作用。他们进一步研究表明,ZmD53抑制UB3/TSH4对UB2/UB3/TSH4启动子的转录激活活性以调节流苏分支数,通过将UB3过表达转基因植物(UB3-OE)与纯合Zmd53/Zmd53转基因植物进行杂交证明ZmD53通过抑制UB3和TSH4的转录活性,从而减少雄穗分枝数。此外,在玉米纯合子Zmd53/Zmd53转基因植株研究中发现,植株穗状花序分枝分生组织数量明显减少,并伴有小穗对分生组织行排列紊乱,而正常的穗部发育过程中没有以上现象,因此Zmd53影响雄性和雌性花序发育[21]。
4 D53/SMXLs在干旱胁迫中起负调控作用
干旱是对作物生长和产量的主要威胁,通过转录组分析,发现SMXL6、7、8参与了干旱响应,其中smxl6/smxl7/smxl8三突变体表现出了比野生型(WT)更强的耐旱性[59-60]。当叶片表面温度升高,SMXL6,7,8基因功能的缺失导致独脚金内酯信号增强,进而促进气孔关闭[61]。与野生型相比,突变体植株叶片表面温度更高,表皮通透性降低,干旱导致的水分流失和细胞膜损伤减少。从而提高抗旱性[61]。此外,smxl6/smxl7/smxl8对abl诱导的气孔关闭和ABA反应敏感[33]。
苹果的研究发现,D53/SMXLs的同源基因是MdSMXL8.2,其启动子具有许多与胁迫相关的顺式作用元件,如热激响应元件HSE、干旱诱导MYB 结合MBS 等,经实验表明MdSMXL8.2能够响应盐胁迫和干旱胁迫[62]。此外,研究表明独脚金内酯可能通过调节细胞骨架和诱导关键基因AFL1调控植物抗旱性[33],由此推测,D53/SMXLs作为独脚金内酯的抑制因子为其提高植物抗旱性提供新的途径。
5 总结
分枝的生长调控是优化植物形态建成的重要因素,其中分枝的发生是高等植物生长发育过程中重要的生命活动。D53/SMXLs 蛋白作为独脚金内酯信号转导途径的关键蛋白,在植物的分枝发育等多个生物过程中发挥着重要而作用。以往研究水稻和拟南芥中D53/SMXLs基因的调控机制居多,少有关于木本植物中D53/SMXLs基因的研究。木本植物的分枝发育可影响栽植密度,解决树木与农作物的肥水、光照及生长等问题,从而满足平原农区林业生产的需要,如在苹果、樱桃、柑橘等果树作物方面,施用独脚金内酯抑制剂可促进植物分枝,提高果业的质量和效率;在杨树等实用树种方面,通过培育窄冠、速生及干形好的理想品种,以实现最佳的观赏实用价值。
笔者基于杨树分枝发育相关的QTL 定位和杨树不同分枝表型的定量结果发现,D53/SMXLs基因可能参与杨树分枝发育的调控,已创制PagD53-OE和PagD53-ANTI等转基因杨树材料,明确PagD53表达量的改变对杨树分枝发育的影响,并已创制PagD53启动子和PagD53-GRF等转基因杨树材料,进而通过GUS染色和ChIP-seq技术分析PagD53组织表达模式和下游调控靶基因及分子机理,为木本植物D53/SMXLs基因的研究提供更多的理论依据。
关于独脚金内酯信号途径的许多步骤还有待完善与研究:(1)近年来研究发现,独脚金内酯、乙烯和细胞分裂素等激素间很可能存在一个动态平衡,相互协同来调控植物的发育,D53/SMXLs基因作为独脚金内酯抑制因子,是否通过细胞分裂素和乙烯调控植物分枝和生根?(2)拟南芥中的SMXL6作为转录因子可直接与SMXL6/7/8的启动子结合,进而抑制SMXL6/7/8的表达,其他同源基因的植物中是否具有这种转录因子的功能有待进一步研究,而且SMXL6与SMXL6/7/8启动子之间是否存在相互选择的机制。(3)虽然D53/SMXLs基因已被证实参与独脚金内酯的信号转导途径,但从类胡萝卜素作为前体到与激素受体结合再到信号转导是一个复杂的过程,D53/SMXLs基因是否与其他基因存在更多的酶促反应?独脚金内酯完整的信号转导途径的发现,让人们对植物分枝的调控有了新的理解,也将引发人们对作物改良的全新思考,为其在实际生产中的广泛应用提供科学依据。
猜你喜欢
语文周报·教研版(2021年28期)2021-08-19
云南医药(2021年3期)2021-07-21
天津中医药(2020年5期)2020-06-01
数学理论与应用(2017年2期)2017-06-27
数学理论与应用(2017年2期)2017-06-27
中成药(2017年4期)2017-05-17
长春中医药大学学报(2017年1期)2017-04-16
中国民族医药杂志(2016年6期)2016-05-09
数学理论与应用(2016年1期)2016-02-28
中国医药导报(2015年24期)2015-02-28
