
饥饿时间对日本沼虾能源物质利用和能量收支的影响
2010-11-07 07:15刘存歧王军静王军霞张亚娟王晓菲
河北大学学报(自然科学版) 2010年6期
刘存歧,王军静,王军霞,张亚娟,王晓菲
(河北大学生命科学学院,河北保定 071002)
饥饿时间对日本沼虾能源物质利用和能量收支的影响
刘存歧,王军静,王军霞,张亚娟,王晓菲
(河北大学生命科学学院,河北保定 071002)
研究饥饿时间对不同体重的日本沼虾(M acrobrachium nip ponense)能源物质利用和能量收支的影响.结果表明:饥饿初期日本沼虾主要依赖脂肪和碳水化合物供能,随着饥饿时间增加,开始部分消耗体内的蛋白质,最后完全依赖蛋白质作为能量来源,虾体营养组成和含量影响供能物质的利用.能量收支方程表明,在饥饿状态下日本沼虾代谢活动减弱,代谢能(R)逐渐降低,而排泄能(U)逐渐增加,体重对日本沼虾能量收支不产生影响.
饥饿;日本沼虾;能量收支
甲壳动物在饥饿过程中会动用机体储存的能量来维持生命活动,同时也会调整代谢水平适应环境的改变[1].有学者发现甲壳动物对机体储存物质的动用存在先后次序,不同甲壳动物由于生理特征、食性、生长阶段等因素的差异,在饥饿状态下对能量物质的利用特点不同[2].饥饿将影响动物的代谢活动和内源性储存物质的消耗,蛋白质、脂肪和碳水化合物的消耗比率决定着氧氮质量比的大小,Harris[3]提出根据氧氮质量比可以估计甲壳动物能量代谢中能源物质的化学本质.日本沼虾(M acrobrachium nipponense)是广泛分布于我国淡水水域中的一种重要经济虾类,具有较高的经济价值[4].目前饥饿对日本沼虾代谢和补偿生长的影响已有较多研究[5-6],而饥饿条件下日本沼虾能源物质利用方式和能量收支的变化相关报道较少.因此,本实验通过研究日本沼虾饥饿期间的氧氨质量比并建立能量收支方程,探讨日本沼虾的能源利用本质及对饥饿的适应.
1 材料与方法
1.1 实验材料
实验所用日本沼虾购自河北省白洋淀,在玻璃水族箱(规格为30 cm×15 cm×18 cm)中驯养1周,水温为26~28℃,驯养期间每日定时(9:00和16:00)投喂体重5%的人工饵料(表1),残饵及粪便于下次投喂前吸出.养殖用水为曝气自来水,水中放置遮蔽物.

表1 实验饵料配方[7]Tab.1 Ingredien t and proximate com position of the experimen tal diets
1.2 实验方法
驯养结束后,选取体重((1.405 1±0.174 0)g,(0.581 8±0.160 6)g)不同的2组日本沼虾,进行饥饿实验,分别记为Ⅰ组和Ⅱ组.每组6个平行,每个平行4尾虾,养于12个水族箱中,内放遮蔽物,防止互相残食.实验开始前停食1 d.每日定时测定日本沼虾的耗氧率、排氨率、体重损失量,用XRY-1A型氧弹能量计测定日本沼虾单位体重的能值,以计算日本沼虾的生长能(由于体重下降,生长能为负值),饥饿时间为8 d.
1.3 耗氧率、排氨率的测定及氧氮质量比的计算
1.3.1 耗氧率、排氨率的测定
每日每个平行取2尾日本沼虾测定耗氧率和排氨率,耗氧率采用碘量法测定,Na2SO3滴定[8].排氨率采用奈氏试剂法测定,721紫外-可见分光光度计比色[9].耗氧率和排氨率的计算公式如下:

1.4 能量收支方程的建立
本实验中日本沼虾饥饿期间无摄食能,只有能量消耗,生长能(G)为负值,通过日本沼虾机体体重丢失计算而得,生长能计算公式为U+R=G[11].
排泄能测定(U):计算出排氨率后,按照每mg氨氮等于24.83 J换算为排泄能[11].
代谢能测定(R):本实验日本沼虾代谢以呼吸为主,由耗氧率计算得到.
1.5 数据处理
用Sp ss13.0对实验数据进行方差分析和线性相关分析处理.
2 结果与分析
2.1 饥饿对不同体重日本沼虾氧氮质量比的影响
饥饿状态下Ⅰ组日本沼虾的氧氮质量比变化如图1所示.经过8 d的饥饿,日本沼虾的氧氮质量比呈下降趋势,由初期的29.98下降到8 d后的10.39,下降65.34%.饥饿第1,2,3天,氧氮质量比均在24以上.第4,5,6天迅速下降至16,到了第7,8天,氧氮质量比降为10,维持在较低水平.8天饥饿过程中,Ⅱ组日本沼虾氧氮质量比呈下降趋势,饥饿第1天,氧氮质量比较高为37.58,饥饿第2天迅速下降至19(p<0.05),至饥饿第7,8天,氧氮质量比降为最低(8.53),较初值下降了77.30%.
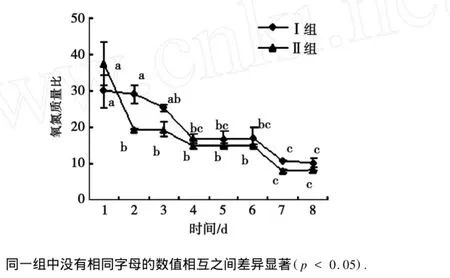
图1 日本沼虾饥饿状态下的氧氮质量比Fig.1 Change of ratio of oxygen to n itrogen in M acrobrachium nip ponense dur ing starvation
2.2 饥饿对不同体重日本沼虾能量收支的影响
不同体重日本沼虾饥饿条件下能量收支方程见表2.由表2可知,代谢能在能量支出中所占比重较大,饥饿第1天,Ⅰ组日本沼虾能量支出中94.5%为代谢能,Ⅱ组日本沼虾能量支出中95.8%为代谢能,随饥饿时间增加,2组日本沼虾代谢能支出逐渐降低,排泄能呈上升趋势.饥饿至第7,8天时,2组日本沼虾的排泄能支出与饥饿初始相比显著增加(p<0.05),分别由开始时的5.5%和4.2%增加到13.6%和16.0%.

表2 饥饿对日本沼虾能量收支的影响Tab.2 Effect of starvation on energy budget of M acrobrachium nip ponense
3 讨论
3.1 饥饿状态下日本沼虾对能源物质的利用
饥饿将影响动物的代谢活动和内源性储存物质的消耗,Harris[6]提出根据氧氮质量比可以估计甲壳动物能量代谢中能源物质的化学本质,氧氮质量比的大小表示蛋白质、脂肪和碳水化合物提供给生物体能量的比率.Mayzaud[12]认为若完全以蛋白质为代谢底物供给能量时,其氧氮质量比为7~10.Ikeda[13]指出以蛋白质和脂肪供能时,氧氮质量比为24,随着脂类和碳水化合物被利用比例的增高,氧氮质量比逐渐增大,如能量完全由脂肪和碳水化合物提供,其氧氮质量比将无穷大.
经研究发现,饥饿过程中甲壳动物对机体储存营养物质的利用次序有所不同,褐虾(Crangon)[14]和日本囊对虾(Marsupenaeusjaponicus)[15-16]先以碳水化合物为主要代谢能源,接着动用脂肪和蛋白质,最后主要由蛋白质供能.凡纳滨对虾(Litopenaeusvannamei)先利用脂肪,其次为碳水化合物和蛋白质[17].而大额糠虾(Gnathophausiaingens)[18]、锯缘青蟹(Scyllaserrata)幼体[19]开始主要由蛋白质和脂肪供能,而后逐渐由脂肪供能.Barclay等[20]研究指出,蛋白质是食用对虾(Penaeusesculentus)饥饿过程中的主要能量来源.而罗氏沼虾(Macrobrachiumrosenbergii)饥饿过程中却利用脂肪和碳水化合物作为主要能源[21].因此饥饿条件下,不同种类的甲壳动物在不同的生长阶段有不同的能源利用方式.
王军霞[22]等已经发现日本沼虾饥饿过程中机体的水分含量和灰分含量呈上升趋势,蛋白质和脂肪含量呈下降趋势,但脂肪的损失率大大超过蛋白质损失率,表明日本沼虾在饥饿过程中主要消耗脂肪作为身体的能量来源.本研究发现,随着饥饿时间的延长日本沼虾的氧氮质量比呈下降趋势,说明日本沼虾在饥饿状态下首先动用脂肪和碳水化合物作为主要能源,而后利用脂肪和蛋白质,最后主要利用蛋白质作为供能物质.Ⅰ组日本沼虾在饥饿第1,2,3天时的氧氮质量比均大于24,说明饥饿初期主要能源为脂肪和碳水化合物;第4,5,6天时,氧氮质量比显著下降为16,说明随着饥饿时间的延长,动用蛋白质的比例逐渐增加.至饥饿的第7,8天,氧氮质量比降至最低(10),表明此时日本沼虾主要利用蛋白质为能源维持生命活动.
与Ⅰ组实验相比,Ⅱ组日本沼虾在饥饿状态下氧氮质量比变化趋势与Ⅰ组相同,但饥饿第1天,Ⅱ组日本沼虾氧氮质量比为37,显著高于Ⅰ组(p<0.05),表明Ⅱ组日本沼虾在此时,动用主要能量来源脂肪和碳水化合物的比例高于Ⅰ组.饥饿2 d时,Ⅱ组日本沼虾氧氮质量比显著下降为19,与Ⅰ组相比差异显著(p<0.05);饥饿至第7,8天,氧氮质量比降至最低(8),并显著低于Ⅰ组(p<0.05),表明体重较小的Ⅱ组日本沼虾与体重较大的Ⅰ组日本沼虾相比,体内存储的脂肪和碳水化合物能源物质较少,在饥饿的第2天就开始部分消耗体内的蛋白质作为能源物质,并迅速以蛋白质作为主要能源物质,说明日本沼虾机体成分组成及营养状况也影响着对供能物质的利用[19].
3.2 饥饿对日本沼虾能量收支的影响
饥饿状态下,水生动物的代谢、能量分配将进行适应性调整,机体内的能量收支方程为U+R-G=0.饥饿期间机体体重下降,表现出负生长,机体动用自身储存物质用于代谢消耗和排泄耗能[23-25].
在饥饿状态下,2组日本沼虾代谢能在能量支出中所占比例较高,排泄耗能很少.随着饥饿时间的延长,代谢能和排泄能呈现相反的变化趋势,代谢能所占比例逐渐下降,而排泄能则逐渐增加,这与饥饿期间日本沼虾能源物质消耗密切相关,随饥饿时间的延长到饥饿后期,2组日本沼虾依赖机体蛋白质供能的比例越来越大,蛋白质代谢增加,氨是蛋白质代谢的主要产物,因此排泄能在饥饿后期较初期高.
本研究表明在相同饥饿时间内,Ⅰ组和Ⅱ组日本沼虾代谢能和排泄能支出比例相近,差异不显著,说明体重对日本沼虾饥饿状态下的能量分配模式影响不显著.朱小明等[26]研究饥饿状态下机体体重对大黄鱼幼鱼能量分配模式没有显著影响,这种现象是否是水产动物饥饿状态下能量学的一个特征,有待进一步研究.
[1]WHYTE J N C,ENGLAR J R,CARSWELL B L,et al.Influence of starvation and subsequent feeding on body composition and energy reserves in the p raw nPandalusplatyceros[J].Can J Fish Aquat Sci,1986,43:1142-1148.
[2]W EA THERLEY A H,GILLS H S.Recovery grow th follow ing periodsof rest restricted rationsand starvation in rainbow trout,SalmogairdneriRichardson[J].Fish Biol,1981,18:195-208.
[3]HSRRIS E.The nitrogen cycle in Long Island Sound[J].Bulletin of the Bingham Océanographie Collection,1959,17:31-65.
[4]刘瑞玉.中国北部的经济虾类[M].北京:科学出版社,1955:3-4.
[5]BOSTWORTH B G,WOL TERSW R.Compensato ry grow th in juvenile red swamp craw fish,Procambarusclarkii[C]//.ROMA IRE R P.Eighth International Symposium on Astacology,Batonrouge,La USA:Louisiana State Univ,Printing Office,1995:648-656.
[6]吴立新,董双林,姜志强.饥饿对甲壳动物生理生态学影响的研究进展[J].应用生态学报,2004,15(4):723-727.
[7]刘媛,王维娜,王安利,等.牛磺酸对日本沼虾生长及酚氧化酶活性的影响[J].淡水渔业,2005,35(2):28-30.
[8]BRETT J R,ZALA C A.Daily pattern of nitrogen of excretion and oxygen consump tion of sockeye salmon(Oncorhynchus nerka)under contro lled conditions[J].Fisheries Research Board of Canada,1975,32:2479-2486.
[9]GALLAGHTER M,MA TTHEWSA M.Oxygen consump tion and ammonia excretion of the American eel Anguilla rostrata fed diets w ith varying p ro tein energy rations and p rotein levels[J].Wo rld Aquaculture Soc,1987,18:107-112.
[10]邢殿楼,张士凤,吴立新,等.饥饿和再投喂对泥鳅能量代谢的影响[J].大连水产学院学报,2005,20(4):290-294
[11]崔奕波.鱼类能量学的理论与方法[J].水生生物学报,1989,13(4):369-383
[12]MAYZAUD P.Respiration and nitrogen excretion of zoop lankton.Ⅳ.The influence of starvation on themetabolism and the biochem ical composition of some species[J].Marine Biology,1976,37:47-58.
[13]IKEDA T.Nutritional ecology of marine zoop lankton[J].Mere Fac Fish Hokkaido Univ,1974,22:1-97.
[14]CUZN G,CAHU C,ALDRIN J R,et al.Starvation effect onmetabolism of Penaeus japonicus[J].Proc World Maricult Soc,1980,11:410-423.
[15]PAPA THANASSION E,KING P E.Effects of starvation on the fine structure of the hepatopancreas on the common p raw nPalaemonserratus(Pennant)[J].Comp Biochem Physiol,1984,77A:243-249.
[16]吴立新,刘瑜,王欣,等.饥饿和再投喂对日本囊对虾代谢率的影响[J].大连水产学院学报,2007,22(2):109-112.[17]于赫男,林小涛,周小壮,等.饥饿胁迫下凡纳滨对虾能源物质的消耗[J].海洋科学,2006,30(12):43-46.
[18]QUETIN L B,ROSSR M,UCH IO K.Metabolic characteristicsof mid-water zoop lankton:Ammonia excretion,O∶N ratios,and the effect of starvation[J].M ar Biol,1980,59:201-209.
[19]翁幼竹,李少菁,王桂忠.饥饿对锯缘青蟹幼体生化组成的影响[J].厦门大学学报:自然科学版,2002,41(1):84-88.
[20]BARCLA Y M C,DALL W,SM ITH D M.Changes in lipid and p rotein during starvation and themolting cycle in the tiger p raw nPenaeusesculentusHaswell[J].Exp Mar Biol Ecol,1983,68:239-244.
[21]DALL W,SM ITH D M.Oxygen consump tion and ammonia-N excretion in fed and starvation tiger p raw nsPenaeusesculentusHaswell[J].Aquaculture,1986,55:23-33.
[22]王军霞,李志华,谢松.饥饿补偿对日本沼虾生长及生化组成的影响[J].河北大学学报:自然科学版,2005,25(6):644-679.
[23]COMOGL IO L,SMOLKO L,AM IN O.Effectsof starvation on oxygen consumption,ammonia excretion and bio-chemical composition of the hepatopancreason adultmalesof the False Sout hern King crabParalomisgranulose(Crustacea,Decapoda)[J].Comparative Biochemistry and Physiology,Part B,2005,140:411-416.
[24]OURAZO B E,V IANA M T,D′ABRAMO L R.Effect sof starvation and dietary lipid on the lipid and fatty acid composition of muscle tissue of juvenile green abalone(Halio tisful gens)[J].Aquaculture,2004,238:329-341.
[25]温小波,陈立侨,艾春香,等.中华绒螯蟹幼蟹饥饿代谢研究[J].应用与环境生物学报,2001,7(5):432-446.
[26]朱小明,王兴春,姜晓东,等.饥饿状态下大黄鱼幼鱼能量收支的研究[J].厦门大学学报,2002,41(4):509-512.
(责任编辑:赵藏赏)
Effects of Starvation Time on Utilization of Energy Materialsand Energy Budget of M acrobrachium nip ponense
LIU Cun-qi,WANGJun-jing,WANGJun-xia,ZHANG Ya-juan,WANGXiao-fei
(Co llege of Life Sciences,Hebei U niversity,Baoding 071002,China)
Under starvation conditions,the effects of starvation time and body mass on utilization of energy substance and energy budget of M acrobrachium nip ponense were studied.The results showed that the sh rimp relied mainly on body fat and carbohyd rates as energy in the beginning of starvation.A long w ith the increase of starvation time,the p rotein began to be consumed partially.A t last p ro tein was the only source of the energy.Nutrition composition affected the utilization of energy substance.The energy budget equation of M acrobrachium nip ponense under starvation conditions suggested that starvation decreased themetabo lic activity of the shrimp,metabolic energy(R)gradually decreased,w hile the excretion(U)increased gradually.The body massof shrimp had little effectson the energy budget.
starvation;M acrobrachium nipponense;energy budget
Q 958
A
1000-1565(2010)06-0706-05
2010-03-17
河北省教育厅博士基金重点资助项目(B2004501)
刘存岐(1967—),男,河北昌黎人,河北大学教授,主要从事水生动物生理生态研究.
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2021年8期)2021-11-04
畜牧兽医科学(2021年21期)2021-06-27
中国饲料(2020年16期)2020-11-06
中国外汇(2019年19期)2019-11-26
海洋与渔业(2017年11期)2018-01-26
中国财政年鉴(2017年0期)2017-07-04
饲料博览(2016年6期)2016-04-05
中国卫生(2015年10期)2015-11-10
