
桃小食心虫滞育期间糖类含量的变化
2011-04-30 05:45丁惠梅武三安
山西农业科学 2011年6期
丁惠梅,武三安
(北京林业大学林学院,北京100083)
桃小食心虫(Carposina sasakii Matsmura)是我国北方重要的果树害虫,多为一、二化性,老熟幼虫在八九月份脱果后,入土结成扁圆形的冬茧滞育,滞育时间长达半年之久。
目前,关于桃小食心虫的研究,国内绝大多数集中于生态学方面,如预测预报、发生规律、滞育特性[1-5]、防治技术及饲养技术[6-7];少数是关于基础生物学[8-9]以及酶和分子水平[10-12]。国外关于桃小食心虫的研究主要在日本和韩国,其也多是关于生态学[13-14]和形态生理[15]方面的研究,对于越冬期间生化指标含量变化的研究尚未见报道。
同大多数冬滞育昆虫相同,桃小食心虫也要度过长达半年的滞育期,且呆在土中不食不动,死亡率较高,这主要受昆虫自身滞育深度、营养状况以及滞育期间和滞育后发育期间的温度等环境条件的影响,而从表面上又很难鉴定越冬虫体的健康状况。对其他滞育昆虫的研究表明,低温会使细胞膜结构和蛋白功能受损,离子转运通道不畅甚至消失,因而导致昆虫受损或死亡[16-18]。而醇类(如甘油、山梨醇/甘露醇、肌醇等)、糖类(如糖原、海藻糖、葡萄糖/果糖等)、蛋白(如滞育蛋白、脯氨酸、丝氨酸、丙氨酸等)以及脂肪等生物分子(称为冷冻保护剂)已经被证实在0℃以上低温和结冰时,具有保护细胞膜、稳定蛋白结构[19-23],降低渗透压,降低结冰点和减少结冰概率[24-25],保护昆虫免受低温伤害的功能。因此,研究越冬期间桃小食心虫生化指标含量的动态变化,有助于了解该种昆虫的滞育深度和健康状况,结合温、湿度等环境条件对滞育的影响,可以较为准确地预测昆虫滞育后发育进度、春季出蛰期和冬后存活的有效种群数量,从而为越冬后防治提供有价值的虫情信息。
本试验从生化指标变化的角度出发,对桃小食心虫滞育期间总糖含量变化进行研究,以期为该种害虫的测报提供一种新的思路。
1 材料和方法
1.1 预试验中对幼虫出土有效积温的估算
将25℃(L∶D=12∶12)恒温养虫室饲养的桃小食心虫脱果幼虫,待结茧后,取冬茧600头。短暂中间温度过渡后(这段时间不计入),放入恒温15℃、全暗、湿度70%的人工气候箱内,处理60 d;接着再经过短暂中间温度过渡,放入5℃冰箱中处理30 d;最后经过短暂中间温度(过渡时间约5 d,不计入),放入恒温25℃、全暗、湿度70%的人工气候箱内,观察幼虫出土情况。单天出土数据未详细记录,幼虫大量出土(单天20头以上)集中在第15~22天之间。若以文献[13]中发育起点温度7.6℃计算,有效积温大概在700~850℃之间。
1.2 试验设计
1.2.1 降温阶段 将25℃(L∶D=12∶12)恒温养虫室饲养的桃小食心虫脱果幼虫,待结茧后,选取冬茧300头,在人工气候箱或者冰箱内进行不同温度处理:(1)15 ℃,0 d;(2)15 ℃,15 d;(3)15 ℃,30 d;(4)15 ℃,45 d;(5)15 ℃,60 d;(6)15 ℃ 15 d,5 ℃ 15 d;(7)15 ℃ 30 d,5 ℃ 15 d;(8)15℃45 d,5℃15 d;(9)15℃60 d,5℃15 d。
1.2.2 升温阶段 将25℃(L∶D=12∶12)恒温养虫室饲养的桃小食心虫脱果幼虫,待结茧后(1~2 d),取冬茧500头,经过短暂中间温度过渡后,放入恒温15℃、全暗、湿度70%的人工气候箱内,处理30 d;接着在5℃冰箱中处理90 d,期间不进行生物分子含量的测定,然后进行升温阶段试验(只在升温阶段测定生物分子含量)。在人工气候箱内进行不同温度处理:(10)15℃,0 d;(11)15 ℃,15 d;(12)15 ℃,30 d;(13)15 ℃15 d,25 ℃ 15 d;(14)15 ℃ 30 d,25 ℃ 15 d。
1.3 总糖含量的测定
总糖含量的测定参照仵均祥(2002)的方法进行,采用分光光度计法。单头测定,重复5次。
2 结果与分析
2.1 降温阶段
从表1可以看出,在处理(1)~(5)的阶段中,总糖含量整体呈现出持续降低的变化趋势,但下降的幅度越来越小。对单个处理进行分析得到出,处理(1)也就是刚结茧幼虫,含量最高,表明糖类是桃小食心虫的主要能量物质;与0 d时相比较,15 d时降低14.13%,约为0 d时含量的85.87%,表明在滞育开始阶段糖类消耗较为明显;15~30 d时继续降低10.85%,约为0 d时含量的75.02%;30~45 d时又降低9.8%,约为0 d时含量的65.22%;45~60 d时降低了4.33%,约为0 d时含量的60.89%,表明随着滞育深度的推进,能量物质的消耗变得缓慢,这是滞育昆虫的共性。

表1 桃小食心虫降温阶段的总糖含量
在处理(6)~(9)的阶段中,总糖含量整体呈现先降低后升高的趋势,拐点位置出现在处理(8)。单个处理分析得出,处理(6)的总糖含量较处理(2)低,与处理(3)较为接近,但略低于处理(3)。有研究表明,有些昆虫5~10℃能量物质如糖原等转化较明显,低温有利于滞育深度的推进。处理(7)总糖含量介于处理(3)和(4)之间,表现出与处理(4)更为接近,推测桃小食心虫此时的滞育深度可能也更加接近于处理(4)。处理(8)总糖含量降到最低,其后出现拐点,含量开始上升,此时的总糖含量与处理(5)更加接近,滞育深度也可能更加接近处理(5)。处理(9)总糖含量上升,表明其他小分子物质如海藻糖、甘油等又开始向糖原等糖类物质转化,滞育出现解除的迹象。
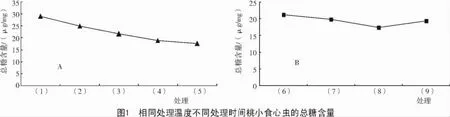
2.1.1 降温时间对总糖含量的影响 在相同的处理温度15℃下,经历不同的处理时间0,15,30,45,60 d,桃小食心虫的总糖含量呈现出持续降低的趋势,但总体看来,下降的速率越来越缓,可能是由于桃小食心虫在开始处理的15 d,尚未完全进入滞育状态,准备滞育但能量物质的储备还未全部完成,此时滞育前期如桃小食心虫是在未脱果前,取食苹果时积累的能量物质如糖类等消耗较为明显,糖原转化成海藻糖、甘油等小分子防冻剂。在15~30,30~45 d,总糖的含量降低速率较之前要慢,这可能有2个方面的原因:一是滞育还处于调整阶段,滞育深度还未达到最深;二是15℃仍然属于比较高的温度,新陈代谢较快,虽然已开始滞育,但能量物质的消耗仍然比较快。45~60 d总糖含量的下降速率明显降低,表明此时桃小食心虫的滞育深度已较深,虽然在比较高的温度下,能量物质的消耗仍然比较缓慢(图1-A)。
在先经历 15℃的 15,30,45,60 d不同处理时间后,再经历5℃相同的时间15 d处理后,桃小食心虫各处理的总糖含量如图1-B所示。在图1-B中处理(8)处出现拐点,其后曲线的变化趋势由下降变为升高。与图1-A的不同之处,在处理(6)~(7)阶段,总糖含量约降低 1.4μg/mg;处理(7)~(8)阶段,总糖含量降低了 2.4μg/mg。有研究表明,较低温度有利于滞育的推进,推测桃小食心虫在5℃时经历相同的处理时间15 d,对滞育深度的加强要比15℃时更明显。处理(8)~(9)的时间段内总糖含量上升,推测在处理(8)时滞育深度已达到最深,再往后滞育开始解除,表现出储能物质又开始积累。
2.1.2 降温温度对总糖含量的影响 在经历相同处理时间(30,45,60 d)不同处理温度(15,5℃)后,桃小食心虫的总糖含量如图2所示。经历相同处理时间不同处理温度后,总糖含量总体上比较接近,有2种类型:类型一,经历5℃低温处理15 d后,总糖含量更高,如处理(7)比处理(4)的总糖含量高约1μg/mg;类型二,经历5℃低温处理15 d后,总糖含量变低,如处理(6)比处理(3)总糖含量低 2μg/mg,处理(8)比处理(5)总糖含量低约0.3μg/mg。在5℃低温下,新陈代谢速率减慢,糖类的消耗较15℃处理中要少,但是低温(0℃以上)有利于滞育深度的加强,2个方面的作用相调节,使总糖含量的变化呈现图2所示的规律。

2.2 升温阶段
本试验升温阶段共有5个处理,如表2所示。由表2可知,处理(10)桃小食心虫总糖含量最高,甚至比降温阶段处理(1)即幼虫刚结茧时的总糖含量还要高,在随后的温度处理中,总糖含量逐渐下降且幅度较大,呈现出下降速率先快后慢的趋势。相同处理温度不同处理时间表现为:处理(10)~(11),(11)~(12)阶段,总糖含量的下降值逐渐增加,分别约为3.0,7.4μg/mg;处理(13)~(14)阶段,总糖含量下降约 3.8μg/mg。相同处理时间不同处理温度表现为:处理(13)桃小食心虫总糖含量较处理(12)低约2.4μg/mg,也就是处理(11)~(13)比处理(11)~(12)总糖含量的下降值更大。

表2 桃小食心虫升温阶段的总糖含量
升温阶段处理(10)总糖含量高,可能是由于在较低温5℃下长期放置的冬茧,滞育深度较深,新陈代谢速率较低,为了适应长期的低温处理的缘故;在升温开始阶段,桃小食心虫体内的总糖含量下降较慢,因为虫体由低温到高温要有一个适应过程,当冬茧感知到升温是持续的、稳定的,才会大量地消耗能量物质,这也是昆虫在进化过程中对自然环境长期适应的结果。
3 结论与讨论
本试验中桃小食心虫体内的总糖含量随着滞育深度的不同而变化。其中,在滞育初期,总糖含量较高,下降速率也较快;随着滞育进程的推进,在未到达滞育维持阶段之前,仍在下降,但速率减缓;滞育开始解除时,含量又略有回升。升温阶段,总糖含量持续降低,可能是用于解除滞育的能量消耗。挑小食心虫的这种变化规律,与已测定的很多种昆虫相似,说明糖类是滞育期间的主要能量物质。根据体内总糖含量,可以推测出桃小食心虫的滞育深度。
桃小食心虫在越冬期间的自然死亡率较高,原因可能是多方面的,其中温度和处理时间是最重要的影响因子,在0℃低温下处理的桃小食心虫冬茧,死亡率随着处理时间的延长而增大,长时间(3个月以上)处理死亡率会高达95%,但是仍然会有部分存活。对0℃低温下处理3个月的冬茧进行生化指标测定,测得的含量较高,这可对本试验升温阶段出现的处理(10)总糖含量偏高的现象作出解释,因为升温阶段中,5℃处理3个月,也存在一定的死亡率,所以生化指标的测定值会偏高。由于桃小食心虫滞育态冬茧的自然死亡率较高,且现在的果园多进行了套袋、喷施农药等管理,再者试验昆虫入土结茧滞育,北方地区冬天温度较低,地面上冻,冬天挖土采集虫源也很困难等原因,对桃小食心虫进行室内生化测定时,选取温度和处理时间相结合来获取不同的滞育深度是一种比较合适的方法。当然,室内试验设计的再合理也无法与大田试验完全相符,所以,如果能结合大田试验的数据,得到的结论将更加可信。
[1] 李秉钧,吴维均,黄可训.光照及温度对桃小食心虫滞育影响的初步研究[J].昆虫学报,1963,12(4):423-431.
[2] 黄可训,王宜智,叶正襄,等.光周期和温度对桃小食心虫滞育的影响[J].昆虫学报,1976,19(2):149-155.
[3] 花保祯,曾晓慧,张皓.不同寄主上桃蛀果蛾的滞育研究[J].西北农业大学学报,1998,26(5):25-29.
[4] 赵飞,王慧.桃小食心虫生物生态学研究进展[J].山西农业科学,2010,38(5):36-38.
[5] 尹河龙,刘贤谦,马瑞艳,等.桃小食心虫种群空间分布型及抽样技术研究[J].山西农业科学,2010,38(6):43-46.
[6] 张月亮,慕卫.光照时间和冷藏对桃小食心虫生长发育和繁殖的影响[J].应用生态学报,2006,17(7):1348-1350.
[7] 范仁俊,李捷,马春森.北方果树食心虫检测和防控新技术研究与示范[J].山西农业科学,2010,38(5):32-35.
[8] 李定绪,王红伟,王佳阳,等.桃小食心虫在不同温度下的实验种群生命表[J].昆虫学报,2010,53(7):773-779.
[9] 李定旭,康照奎,王佳阳,等.桃小食心虫的发育起点温度和有效积温[J].昆虫知识,2010,47(5):923-926.
[10] 谭六谦,王光香.不同寄主桃小食心虫酯酶同功酶的比较研究[J].落叶果树,1989,21(3):21-22.
[11] 韩青梅.6种寄主桃蛀果蛾遗传变异的RAPD分析[J].西北农林科技大学学报:自然科学版,2001,29(3):91-94.
[12] 徐庆刚,花保祯.桃蛀果蛾寄主生物型分化的RAPD分析[J].昆虫学报,2004,47(3):379-383.
[13] Dong-soon Kim,Joon-ho Lee.Egg and larval survivorship of Carposina sasakii (Lepidoptera:Carposinidae)in apple and peach and their effects on adult population dynamics in orchards[J].Entomol,2002,31(4):685-690.
[14] 津川力,山田雅辉.Forecasting the Outbreak of Destructive Insects in AppleOrchards II.On theGrowth of Larval Stage in the Peach FruitMoth,CarposinaniponensisWALSINGHAM[J].Japanese society of applied entomology and zoology,1960,4(2):96-101.
[15] 津川力,山田雅辉,白崎将瑛,等.Forecasting theOutbreak of Destructive Insects in AppleOrchards VI.On the Development of the Gonads in Hibernating Larvae of the Peach Fruit Moth.Carposina niponensisWALSHINGHAM[J].Japanese society of applied entomologyand zoology,1964,8(2):111-117.
[16] Morris G J,Coulson G,Meyer M A.Cold shock,a widespread cellular reaction[J].Cryo-Letters,1983,4:179-192.
[17] Kelty JD,Killian K A,Lee R E.Cold shock and rapid coldhardening of pharate adult flesh flies(Sarcophaga crassipalpis)-effects on behaviour and neuromuscular function following eclosion[J].PhysiologicalEntomology,1996,1:283-288.
[18] Kostal V,Vambera J,Bastl J.On the nature of pre-freezemortality in insects:water balance,ion homeostasis and energy charge in theadultsof Pyrrhocorisapterus[J].Journalof ExperimentalBiology,2004,207:1509-1521.
[19] Bale JS.Insects and low temperatures:from molecular biology todistributionsand abundance[J].Philosophical Transactionsof the RoyalSociety,2002,357b:849-862.
[20] Crowe JH,Crowe LM.Stabilization ofbiologicalmembranesat low wateractivities[J].Cryobiology,1983,20:346-356.
[21] Crowe JH,Carpenter JF,Crowe L M,et al.Are freezing and dehydration similar stressvectors?A comparison of modes of interaction ofstabilizing soluteswith biomolecules[J].Cryobiology,1990,27:219-231.
[22] Carpenter JF,Crowe JH.Themechanism of cryoprotection of proteinsby solutes[J].Cryobiology,1988,25:244-255.
[23] WilliamsW P.Cold-induced lipid phase transitions[J].Philosophical Transaction of the Royal Society of London,1990,326b:555-570.
[24] Storey K B,Storey JM.Biochemistry of cryoprotectants[M]//Lee Jr.,R.E.,Denlinger,D.L.(Eds.).Insects at Low Temperature.New York:Chapman&Hall,1991:64-93.
[25] Lee RE.Principlesof insect low temperature tolerance[M]//Lee R.E.,Denlinger DL.(Eds.).Insects at Low Temperature.New York:Chapman&Hall,1991:131-148.
猜你喜欢
农业技术与装备(2021年9期)2021-12-01
落叶果树(2021年6期)2021-02-12
河北果树(2020年2期)2020-05-25
科技与创新(2016年22期)2017-03-30
农民致富之友(2017年8期)2017-03-18
特产研究(2016年3期)2016-04-12
浙江柑橘(2016年2期)2016-03-11
新疆农垦科技(2014年6期)2014-02-28
新疆农垦科技(2014年2期)2014-02-28
食品科学(2013年14期)2013-03-11
