
边缘型人格障碍遗传学研究
2011-05-16 08:04王兰兰肖泽萍
医学研究杂志 2011年7期
王兰兰 肖泽萍
边缘型人格障碍(BPD)长期以来都是精神医学界争论的焦点,其症状表现具有广泛异质性。根据目前DSM-IV对边缘型人格障碍的诊断标准规定,满足9条标准中的5条即可诊断为BPD,这样通过不同的排列组合就有可能得到151种不同的对BPD的诊断标准。这种状况极大阻碍了对BPD病因、病理机制的研究。于是有专家提出进行症状维度及症状内在表型研究,以减少异质性症状的复杂性。美国精神病协会将边缘型人格障碍的症状维度概括为4大类:①情感不稳定性;②冲动攻击,及自伤、自杀行为;③认知-感知功能缺损;④人际关系紊乱。随着行为遗传学技术的发展,越来越多的研究表明人类复杂的行为背后有其内在的生物学原因。对边缘型人格障碍病因的理解,也有许多专家提出了基因-环境交互模型,认为正是由于先天易感性素质与后天不良环境(儿童期创伤、不良的家庭环境及教养方式等)共同作用,导致了边缘型人格障碍的形成[1~3]。相关分子遗传学研究也进一步证实了边缘型人格障碍病理症状、行为背后存在一定基因缺陷。本文就边缘型人格障碍的遗传学研究进行综述。
一、家系研究
基于家系研究及临床观察,Skodol和Torgersen等发现边缘型人格障碍具有家族聚集性,与健康对照相比,BPD患者一级亲属中患有该病的概率显著增高[4]。Nigg等[5]报道 BPD 一级家属中共病有边缘型人格障碍的概率为11.5%,远高于该病在普通人群中的患病率(1%~2%)。
二、双生子研究
Torgersen等最早在1984年对边缘型人格障碍进行了双生子研究,发现7对同卵双生子中同病一致率为0%,而18对异卵双生子同病一致率为11.1%,提示环境因素对边缘型人格障碍的发病起了重要作用。但由于样本量小,研究方法存在很大局限性,故研究结果受到普遍质疑。2000年,Torgersen等[4]再次调查了92对单卵双生子和129对异卵双生子,发现单卵双生子中同时患有边缘型人格障碍的概率为35%,而异卵双生子中同时患有BPD的概率为7%。最近3项双生子研究报道,边缘型人格障碍的加性遗传度分别为69%、35%、42%。这些研究都证明边缘型人格障碍是一种遗传因素和环境因素共同作用导致的复杂的精神类疾病[4,6,7]。但目前尚没有中国各民族人群中双生子研究的报告。
三、连锁研究
Distel等[8]在荷兰收集了711对双生子的表型数据及基因型数据,同时收集了561对其生物双亲的基因型数据,结果发现边缘型人格障碍与第1、4、9、18号染色体存在显著连锁关系,其中连锁最高峰值在9p D9S286位点上(LOD=3.548,P=0.0001)。
四、BPD症状维度相关的候选基因的关联研究
1.冲动攻击及自伤、自杀行为:冲动攻击性和情感不稳定性是BPD的核心症状。研究表明冲动攻击行为主要与5-HT在大脑内的活性有关。结构及功能影像学研究显示冲动行为患者的前扣带回区、海马及杏仁核功能异常,而这些区域主要分布着5-HT神经递质[9]。大量研究显示冲动攻击性及自伤、自杀行为与5-HT功能下降有关。
5-羟色胺转运体(5-HTT),又名可溶性载体家族6-4号(SLC6A4),其功能是将突触间隙的5-HT回吸收至突触前神经元。人类5-HTT基因定位于17q11.1~12,长约31kb,含14个外显子。该基因已知存在3种多态:①启动子区44bp插入/缺失多态性(5-HTTLPR),其常见的等位基因有两个:短等位基因(S)、长等位基因(L);②第二内含子区由17bp核心序列重复9、10或12次而形成的一个可变数目串联重复多态性(VNTR);③第763位碱基C与A置换。其中前二者可影响该基因的转录活性,改变5-HTT的表达,成为了研究的热点。大量研究证明5-HTT基因对自杀行为、冲动行为、以及不稳定性情感起着重要作用,因此5-HTT基因被认为是边缘型人格障碍的候选基因[10~12]。
色氨酸羟化酶(TPH)是5-HT合成限速酶,TPH基因被认为是导致BPD遗传素质的主要候选基因之一。主要有两种亚型TPH-1和TPH-2。TPH-1基因定位于11p14~15.3区(11p14~p15.3)约长29 kb,至少包括11个外显子,一个5'端非转录区。在内含子7中存在两种多态性,即A218C(rs1800532)和A799C(rs1799913)。有研究显示单核苷酸多态性A779C与冲动行为受试者脑脊液中5-HIAA的浓度密切相关,且该基因多态性与有自杀企图的暴力行为者、共病抑郁的酒精依赖者有显著联系。而也有研究发现在有自杀企图的抑郁症患者中携带A799A等位基因者明显较多。目前对TPH-2的研究相对较少。
单胺氧化酶A(MAO-A)是多巴胺(DA)、5羟色胺(5-HT)及去甲肾上腺素(NE)降解的关键酶,其活性变化直接影响单胺类神经递质的灭活率,从而改变突触间单胺类神经递质的水平。人类MAOA基因位于Xp11.23~11.4上,在其启动子区,编码序列上游112kb处有一个30bp重复序列多态性,可出现2、3、3.5、4及5倍的重复。该多态性影响MAOA基因的转录活性,3.5、4、5倍重复长等位基因比3倍重复等位基因转录效率高2~10倍。长等位基因酶活性升高。研究显示转基因小鼠该基因突变会导致MAO活性丧失,并出现较多的攻击和伤害同伴的行为。Caspi等[13]提出假设:MAOA的低活性将不足以调节环境因素(童年期各类应激事件)所造成的NE、5-HT、DA等神经递质的失衡,从而导致各种冲动性及攻击性行为的出现。Caspi等对1037名儿童从其出生开始到26岁进行的前瞻性队列随访研究发现携带低活性MAOA等位基因的男性儿童遭遇儿童期创伤(如躯体虐待、性虐待)或不良家庭环境(如母性抚养者的拒绝及重要抚养者的缺失)会导致反社会行为明显增加。该研究揭示了并非所有经历严重儿童期创伤的个体都会出现冲动攻击行为的个体差异性原因。随后有相关研究进一步证实MAOA基因多态低活性是形成行为障碍、反社会人格障碍、多动障碍及注意缺陷的危险因素。目前虽然尚没有研究探索MAOA与BPD的关系,但基于BPD具有冲动攻击性的临床特点,胆碱能系统高反应性及5-HT系统功能紊乱的生物学特性,以及高度纠结的遗传-环境交互作用的病因模式,对MAOA与BPD的关系进行研究是非常必要的。
2.情感不稳定性:儿茶酚-氧位-甲基转移酶(COMT)也是NE、5-HT、DA的分解代谢酶。COMT基因定位于22p11区域,在第4外显子158号密码上可发生碱基G与A的置换,从而导致COMT上第158个氨基酸残基出现氨基酸Val与Met的置换,从而影响COMT的活性。基因型为Val/Val酶具有高活性,Val/Met基因型的酶具有中度活性,而Met/Met基因型的酶活性低。Val158基因多态性对人类行为和精神功能具有多效作用[14]。Smolka等[15]发现杏仁核、海马、边缘皮质结构中随着Met158等位基因数增加,受试者对负性情绪刺激的反应增强。研究发现Val158等位基因与成人抗负性情绪能力密切相关。而BPD患者存在显著的负性情绪调节障碍。Tadic等[16](2008年)研究也发现BPD患者中存在更多的Met158Met基因型,且Met158Met基因型与5-HTTLPR(S)等位基因密切相关。
去甲肾上腺素转运体(norepinephrine transporter,NET)位于肾上腺素能神经突触前膜上,其功能是将神经元释放的NE再摄取回到突触前膜中,对调控突触间隙中NE浓度、终止神经冲动信号、维持受体对神经递质的敏感性极为重要。人类NET基因(SLC6A2)位于16q12.2染色体上,已有文献报道NET有 14种基因突变,其中启动区 rs2242446(-182T/C)和外显子9的rs5569(1287G/A)是目前认为突变率较高且可能与情感性精神障碍相关的单核苷酸多态性[17]。国内一项汉族人群中的研究显示[18]:携带rs2242446 C等位基因和rs5569 A等位基因可能是情感障碍的易感因素。目前尚没有关于NET与BPD的相关研究,鉴于去甲肾上腺素能系统对应激反应的调节作用提示相关研究具有一定必要性。
3.认知功能缺损:2004年Friedel等[19]提出边缘型人格障碍多巴胺功能紊乱假说。该假说的提出主要基于动物及人类实验和临床药物试验。大量研究显示DA参与情绪信息处理、冲动控制、及认知过程。DA参与调节对显著正性或负性事件的情绪反应,以及对刺激的情绪唤醒;在调节工作记忆及认知的过程中DA也起着重要作用;有研究显示脑脊液中高香草酸(HVA)的浓度与BPD患者的冲动行为相关。另外大量临床研究显示非典型抗精神病药物治疗BPD,对改善其敌意、偏执观念、矛盾情感、冲动控制等临床症状显著有效,如利培酮,奥氮平[20]。而非经典抗精神病药物主要作用于阻断D2受体。而且BPD与物质滥用的高共病率也显示患者可能存在多巴胺能奖赏通路异常。因此与DA功能密切相关的多巴胺转运体基因(DAT1)可能是BPD的重要候选基因。
多巴胺转运体主要功能是再摄取DA,是调节多巴胺能神经递质的重要物质。DAT1基因定位于5p15.3,该基因cDNA3'端末翻译区有一段40bp重复序列,以其为核心序列可出现3~11倍重复序列,常见的是9-、10-重复序列。该多态性影响了DAT1基因的表达,9/10基因型的个体比10/10基因型者转运能力减低约1/4。老鼠DAT1基因失活会导致DA在细胞外持续时间延长。有研究显示9-重复序列 DAT1与BPD有关[21]。
4.人际关系的高敏感性:Gunderson(2008年)提出BPD患者的人际关系高敏感性是由于患者先天素质(5-HT系统和DA系统功能异常)和后天环境(不安全的依附关系)交互作用所致。患者在情感不稳、冲动控制障碍的基础上,同时存在信息处理能力缺陷,于是在后天不良环境因素刺激下发展出分裂、投射等不成熟的防御方式,以及“全或无”的认知特点,患者对客体的认知波动于极端理想化和极端贬低之间,对自我的认识也处于极端不稳定状态,从而导致高度敏感和极端不稳定的人际关系。
五、BPD关联研究
目前直接对边缘型人格障碍群体进行的分子遗传学研究还比较有限。检索美国Pubmed数据库,发现对边缘型人格障碍的关联研究主要集中在对5-HTT、HTR2A、DAT1、COMT 等候选基因(表1)。国内尚没有相关研究报道。
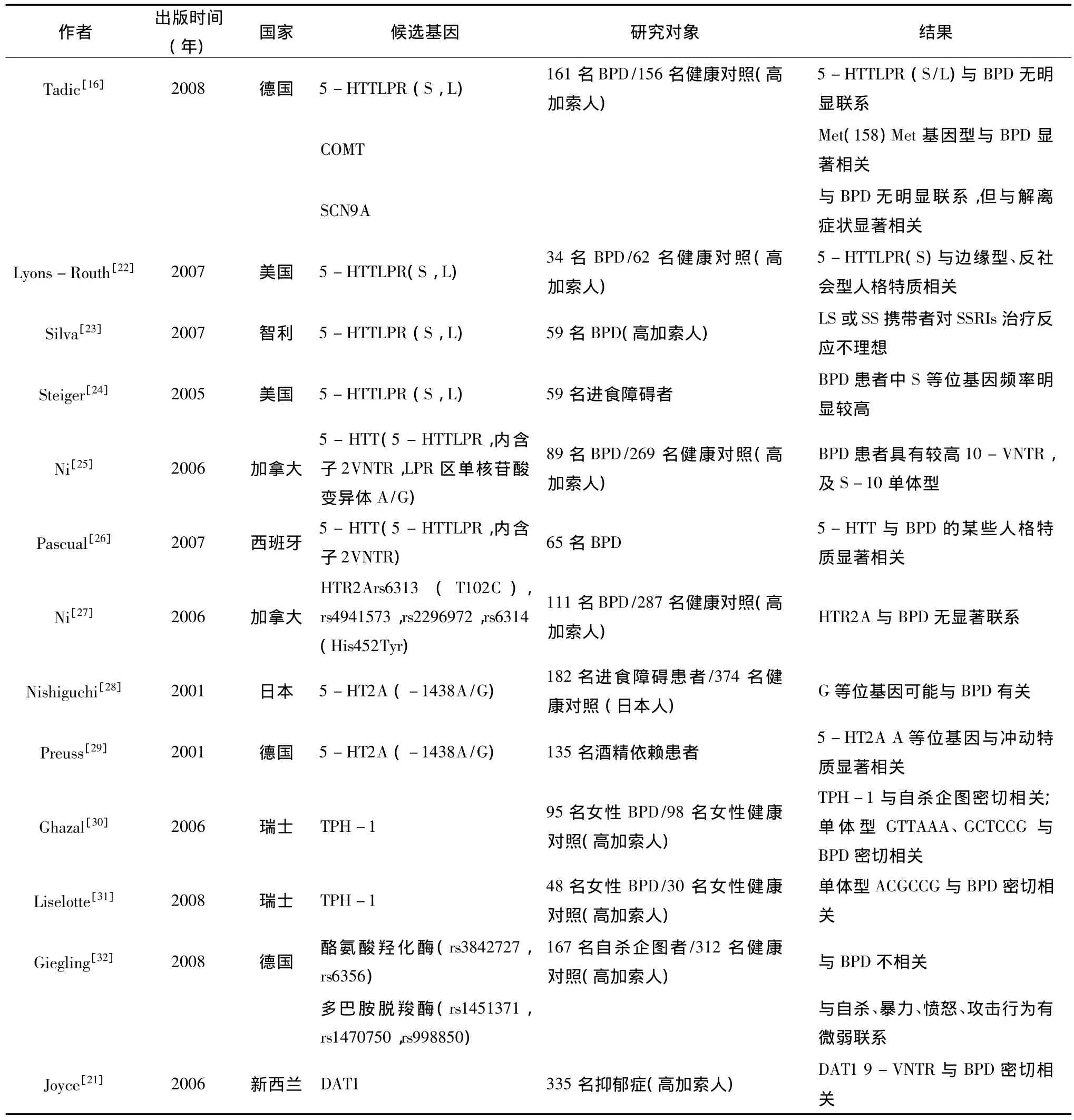
表1 边缘型人格障碍的关联研究
综合这些研究结果,我们发现有6项研究报道了5-HTT与BPD及其症状的关系,其中两篇文献报道5-HTTLPR(S)等位基因与BPD人格特质有关,且其携带者对SSRIs治疗反应欠佳;一项研究显示,与健康对照相比,BPD的5-HTT基因在VNTR标记及单体型上有显著差异。这些结果提示BPD与5-HTT基因有密切联系,5-HTT基因在BPD的发病上可能起着重要作用。3项关于5-HT2A基因的研究中,有两项研究报道高加索人群中5-HT2A基因多态性与BPD无明显联系,而一项对日本人群的研究显示该5-HT2A(-1438A/G)G等位基因可能与BPD有关。5-HT2A基因与BPD的关系有待进一步研究证明。两项对TPH-1的研究均显示该基因多态性与BPD密切相关。研究显示与疾病密切相关的单体型构型位于TPH-1启动子与内含子3之间。但两次研究结果的单体型结论不一。研究报道,9-重复序列等位基因是BPD的危险因素,而且增加了有儿童期创伤者罹患BPD的可能性,进一步论证了关于BPD形成的基因-环境交互模式。一项研究显示COMT Met(158)Met基因型与BPD显著相关,P=0.0085。根据目前文献复习,低活性COMT可能与患者负性情绪处理障碍有关,患者可能会出现更多更强烈的情绪不稳定的临床症状。但研究中并未报道该基因型与BPD相关症状的关系。如果该结论能够得到复制,则COMT与BPD的关系能够得到进一步明确。目前的研究中还涉及对SCN9A基因(疼痛觉相关基因)、酪氨酸羟化酶、多巴胺脱羧酶的研究,但未发现与BPD密切相关。仅SCN9A基因的研究发现其与BPD的解离症状密切相关。
遗传学研究显示边缘型人格障碍是一种累及多基因微效作用的精神疾病,缺陷基因主要集中于5-HT通路,也累及DA通路。环境因素(儿童期创伤经历)可能在缺陷基因是否致病的过程中起着重要作用。
1 Marianne Goodman,Antonia New,Larry Siever.Trauma,Genes,and the Neurobiology of Personality Disorders.Ann N Y Acad Sci.2004;1032:104-116
2 Skodol AE,Siever LJ,Livesley WJ,et al.The borderline diagnosis II:biology,genetics,and clinical course.Biol Psychiatry,2002,15;51(12):951-663
3 Gunderson JG,Lyons-Ruth K.BPD'S interpersonal hypersensitivity phenotype:A gene-environmental model.Journal of Personality Disorders,2008,22(1):22-41
4 Torgersen S.Genetics of patients with borderline personality disorder.Psychiatr Clin North Am.2000,23(1):1-9
5 Nigg JT,Goldsmith HH.Genetics of personality disorders:perspectives from personality and psychopathology research.Psychol Bull,1994,115(3):346-380
6 Torgersen S,Czajkowski N,Jacobson K,et al.Dimensional representations of DSM-IV cluster B personality disorders in a populationbased sample of Norwegian twins:a multivariate study.Psychol Med,2008,38(11):1617-1625
7 Distel MA,Trull TJ,Derom CA,et al.Heritability of borderline personality disorder features is similar across three countries.Psychol Med,2008,38(9):1219-1229
8 Distel MA,Hottenga JJ,Trull TJ,et al.Chromosome 9:linkage for borderline personality disorder features.Psychiatr,Genet.2008,18(6):302-307
9 Soloff PH,Meltzer CC,Becker C,et al.Impulsivity and prefrontal hypometabolism in borderline personality disorder.Psychiatry Res,2003,30;123(3):153-163
10 Bondy B,Erfurth A,Jonge S,et al.Possible association of the short allele of the serotonin transporter promoter gene polymorphism(5-HTTLPR)with violent suicide.Mol Psychiatry,2000,5(2):193-195
11 Frankle WG,Lombardo I,New AS,et al.Brain serotonin transporter distribution in subjects with impulsive aggressivity:a positron emission study with[11c]McN 5652.Am J Psychiatry,2005,162(5):915-923
12 Hoefgen B,Schulze TG,Ohlraun S,et al.The power of sample size and homogenous sampling:association between the 5-HTTLPR serotonin transporter polymorphism and major depressive disorder.Biol Psychiatry,2005,1;57(3):247-251
13 Caspi A,McClay J,Moffitt TE,et al.Poulton R.Role of genotype in the cycle of violence in maltreated children.Science,2002,297(5582):851-854
14 Hosák L.Role of the COMT gene Val158Met polymorphism in mental disorders:A review.Eur Psychiatry,2007,22(5):276-281
15 Skodol AE,Siever LJ,Livesley WJ,et al.The borderline diagnosis II:Biology,genetics,and clinical course.Biol Psychiatry,2002,15;51(12):951-963
16 Tadic A,Victor A,Baskaya O,et al.Interaction between gene variants of the serotonin transporter promoter region(5-HTTLPR)and catechol O-methyltransferase(COMT)in borderline personality disorder.Am J Med Genet B Neuropsychiatr Genet,2009,150B(4):487-495
17 Zill P,Engel R.Identification of naturally occurring polymorphism in the promoter region of the norepinephrine transporter and analysis in major depression.Neuropsychopharmacology,2002;26(4):489-493
18 Sun N,Xu Y,Wang Y,et al.The combined effect of norepinephrine transporter gene and negative life events in major depression of Chinese Han population.J Neural Transm,2008,115(12):1681-1686
19 Friedel RO.Dopamine dysfunction in borderline personality disorder:a hypothesis.Neuropsychopharmacology,2004,29(6):1029-1039
20 Zanarini MC,Frankenburg FR.Olanzapine treatment of female borderline personality disorder patients:a double blind,placebo-controlled pilot study.J Clin Psychiatry,2001,62(11):849-854
21 Joyce PR,McHugh PC,McKenzie JM,et al.A dopamine transporter polymorphism is a risk factor for borderline personality disorder in depressed patients.Psychol Med,2006,36(6):807-813
22 Lyons K,Holmes BM,Sasvari M,et al.Serotonin transporter polymorphism and borderline or antisocial traits among low-income young adults.Psychiatr Genet,2007,17(6):339-343
23 Silva H,Iturra P,Solari A,et al.Serotonin transporter polymorphism and fluoxetine effect on impulsiveness and aggression in borderline personality disorder.Actas Esp Psiquiatr,2007,35(6):387-392
24 Steiger H,Joober R,Israel M,et al.The 5-HTTLPR polymorphism,psychopathologic symptoms,and platelet[3H-]paroxetine binding in bulimic syndromes.Int J Eat Disord,2005,37(1):57-60
25 Ni X,Chan K,Bulgin N,et al.Association between serotonin transporter gene and borderline personality disorder.J Psychiatr Res,2006,40(5):448-453
26 Pascual JC,Soler J,Baiget M,et al.Association between the serotonin transporter gene and personality traits in borderline personality disorder patients evaluated with Zuckerman-Zuhlman Personality Questionnaire(ZKPQ).Actas Esp Psiquiatr,2007,35(6):382-386
27 Ni X,Bismil R,Chan K,et al.Serotonin 2A receptor gene is associated with personality traits,but not to disorder,in patients with borderline personality disorder.Neurosci Lett,2006,20;408(3):214-219
28 Nishiguchi N,Matsushita S,Suzuki K,et al.Association between 5HT2A receptor gene promoter region polymorphism and eating disorders in Japanese patients.Biol Psychiatry,2001,50(2):123-128
29 Preuss UW,Koller G,Bondy B,et al.Impulsive traits and 5-HT2A receptor promoter polymorphism in alcohol dependents:possible association but no influence of personality disorders.Neuropsychobiology,2001,43(3):186-191
30 Ghazal Zaboli,Rinat Gizatullin,Asa Nilsonne,et al.Tryptophan hydroxylase-1 gene variants associate with a group of suicidal borderline women.Neuropsychopharmacology,2006,31(9):1982-1990
31 Liselotte Maurex,Ghazal Zaboli,Stefan Wiens,et al.Emotionally controlled decision-making and a gene variant related to serotonin synthesis in women with borderline personality disorder.Scand J Psychol,2009,50(1):5-10
32 Giegling I,Moreno-De-Luca D,Rujescu D,et al.Dopa decarboxylase and tyrosine hydroxylase gene variants in suicidal behavior.Am J Med Genet B Neuropsychiatr Genet,2008,147(3):308-315
猜你喜欢
兵团教育学院学报(2022年4期)2022-10-15
世界科学技术-中医药现代化(2022年3期)2022-08-22
中国典型病例大全(2022年7期)2022-04-22
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
智慧健康(2021年17期)2021-07-30
心理与健康(2021年2期)2021-07-22
昆明医科大学学报(2021年2期)2021-03-29
新课程·下旬(2018年9期)2018-11-14
青少年科技博览(中学版)(2015年10期)2015-01-11
