
伪威氏按蚊mtDNA-Cytb基因群体遗传学研究*
2011-08-21 06:51汤林华周水森潘嘉云王学忠杨曼尼卓玛央金
中国人兽共患病学报 2011年12期
刘 茜,武 松,汤林华,周水森,黄 芳,潘嘉云,王学忠,杨曼尼,卓玛央金
2.中国疾病控制中心寄生虫病预防控制所,世界卫生组织疟疾、血吸虫病和丝虫病合作中心,上海 200025;
3.云南省寄生虫病研究所,普洱 665000;
4.西藏自治区林芝地区疾病预防控制中心,林芝 860100
课题组于2007与2010年在西藏疟疾流行区墨脱县进行媒介调查发现,该县的按蚊主要包括多斑按蚊复合体和带足按蚊,对多斑按蚊复合体进行实验室种型鉴定发现包括伪威氏按蚊和威氏按蚊[1],其中伪威氏按蚊为绝对优势蚊种,在伪威氏按蚊中采用巢氏PCR方法检测出间日疟原虫SSU r DNA基因特异序列[2],从而从分子角度判定伪威氏按蚊为该疟疾流行区的传疟媒介。云南省与西藏疟疾流行区相邻,伪威氏按蚊在该省分布广泛。课题组拟采用线粒体细胞色素B基因对西藏、云南和缅甸拉咱市的伪威氏按蚊进行群体遗传分析,以期为云南省伪威氏按蚊在疟疾传播作用方面提供研究思路。
1 材料与方法
1.1 样本采集 所有按蚊成蚊标本课题组采集于2007年西藏与云南及缅甸现场。采集到的按蚊经过形态学鉴定至多斑按蚊复合体,带回实验室采用分子方法进行具体种型鉴定。5个群体共获伪威氏按蚊共44份样本用于mt DNA-Cytb基因群体遗传分析,见表1。

表1 伪威氏按蚊采集地点与编号Tab.1 Anopheles pseudowillimori collection spot and ID
1.2 mt DNA-Cytb基因扩增
1.2.1 基因提取 采用简易快速DNA提取法[3],即将1~2个蚊腿置于1.5 m L离心管,加入35μL裂解液(10 mmol/L p H8.0 Tirs HCL,1 mmol/L EDTA,1%Nonidet P-40 100μg/m L蛋白酶 K),匀浆;再加入35μL消毒双蒸水,煮沸5 min;10 000 r/min离心2 min,取上清液置-30℃备用。
1.2.2 扩增体系与条件 采用文献报道的扩增方案[4],正 向 引 物:5′-GGA CAA ATA TCA TTT TGA GGA GCA ACA G-3′;反向引物为:5′-ATT ACT CCT CCT AGC TTA TTA GGA ATT G-3′。扩增体系为50μL,体系包括10×PCR缓冲液5.0 μL,4.5μL MgCl2(25 mmol/L),1.0μL d NTPs(10 mmol/L),正、反向引物0.8μL(20 mmol/L),2.5 μL redTaq聚合酶(1U/μL),1.5μL DNA 模板,33.9μL dd H2O。反映条件94℃预变性2 min,94℃变性30 s,45℃退火30 s,72℃延伸30 s,共35个循环,72℃孵育8 min。扩增产物送基因公司进行测序。
1.3 数据分析 测序结果以Chromas(Version 2.13)进行核对,必要时手工调整,序列比对采用Clustal X进行,MEGA软件包统计序列特征,包括碱基含量、转换和颠换的数目等,并计算序列的差异性(p距离),依据差异性构建邻接树(Neighbor Joining Tree,NJ)。将序列数据转换为ARLEQUIN(Version 3.0)可识别的文件后,统计和计算各群体的基因序列碱基分化参数,置换位点数目、单倍型的类型和在群体中的分布等。单倍型之间的系谱关系由TCS 1.21[5])软件包构建。群体遗传结构由ARLEQUIN的AMOVA模块计算,包括群体间的分化参数Fst(F-statistics)、基因流水平Nm(Nm=1-Fst/4Fst)和群体内和群体间的变异成分。
2 结 果
2.1 扩增情况 受检伪威氏按蚊DNA标本均扩增出mt DNA-Cytb基因目标片段,扩增片段约为445 bp,所有的扩增产物在琼脂糖凝胶中观察,均为单一、清晰的条带,长度与预期一致。
2.2 伪威氏按蚊mt DNA-Cytb序列特征 测序获得伪威氏按蚊Cytb基因共44条,比对序列,发现差异极小,经GenBank的BLAST比对,确认该序列片段为特异扩增的Cytb基因。在序列比对后,取其中的420 bp部分片段进行后续分析,结果显示,碱基置换的位点为26个,其中23处转换(50.0%),2处颠换(7.69%),1个超变位点(3.85%),共27个单倍型(61.36%),其中有6个单倍型在群体中共享,见表2。该片段的平均AT含量为75.6%,GC含量为24.4%。
2.3 单倍型之间的关系 将本研究得到的伪威氏按蚊Cytb基因的33个单倍型应用TCS1.21软件包建立95%简约家系网络图(图1),网络图中每个圈代表其中一个单倍型,空白部分是未检测到的单倍型,在家系网络图中各单倍型呈高水平的平行演化,COI_DX02(18)、COI_DX01(12)和 COI_MN16(5)在群体中的分布丰度较高,分别占27.69%、18.46%和7.69%,COI_DX02作为外群的最大可能值0.2125,Cytb_cy15作为外群的最大可能性为0.1925。应用Mega软件包计算本研究伪威氏按蚊mt DNA-Cytb基因单倍型之间的遗传距离,并依据遗传距离构建聚类关系树(图2),树的拓扑关系显示各单倍型之间的聚类关系没有明显的与地理分布呈相关性。
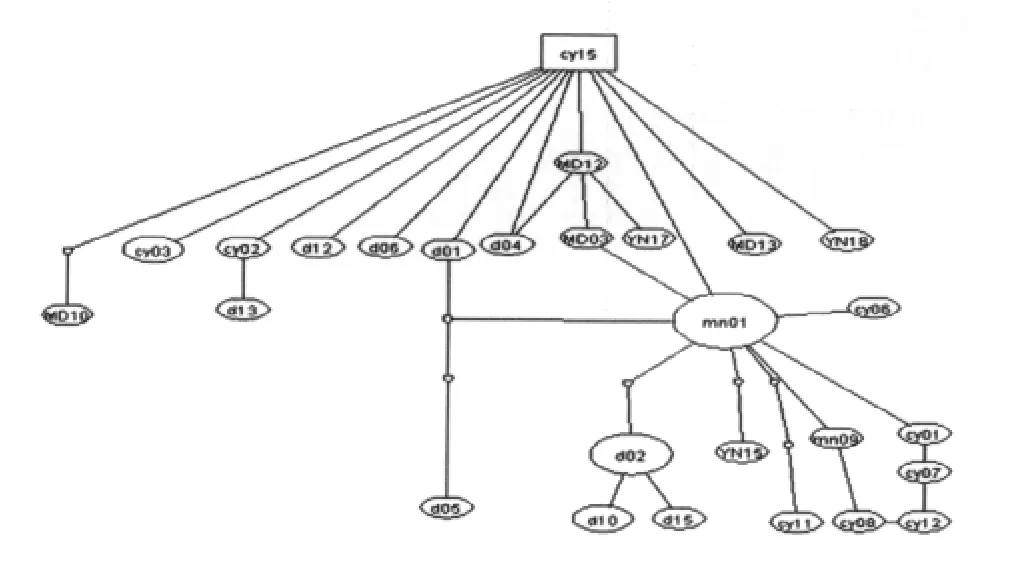
图1 伪威氏按蚊mtDNA-Cytb基因单倍型的家系网络图Fig.1 Family constellation network map of mtDNA-Cytb haplotypes of Anopheles pseudowillimori
2.4 群体遗传结构 分步AMOVA计算结果显示,群体内变异绝对大于群体间变异,群体内差异占总差异的98.30%,群体间占1.70%,见表2,所有群体的Fst值为0.016 96,见表3,Nm为4.168,大于1,提示基因流水平可以完全抵御遗传漂变造成的群体分化,群体间几乎没有发生遗传分化。本研究结果显示西藏墨脱县与云南景洪群体间Fst值最大(0.086 50),云南勐腊与缅甸拉咱市、察隅和墨脱群体间的Fst值依次为-0.081 12、-0.003 81和-0.103 03;墨脱与缅甸拉咱市群体间的Fst值为-0.005 12,提示上述群体相互间基因流水平非常高,群体遗传分化程度很小。
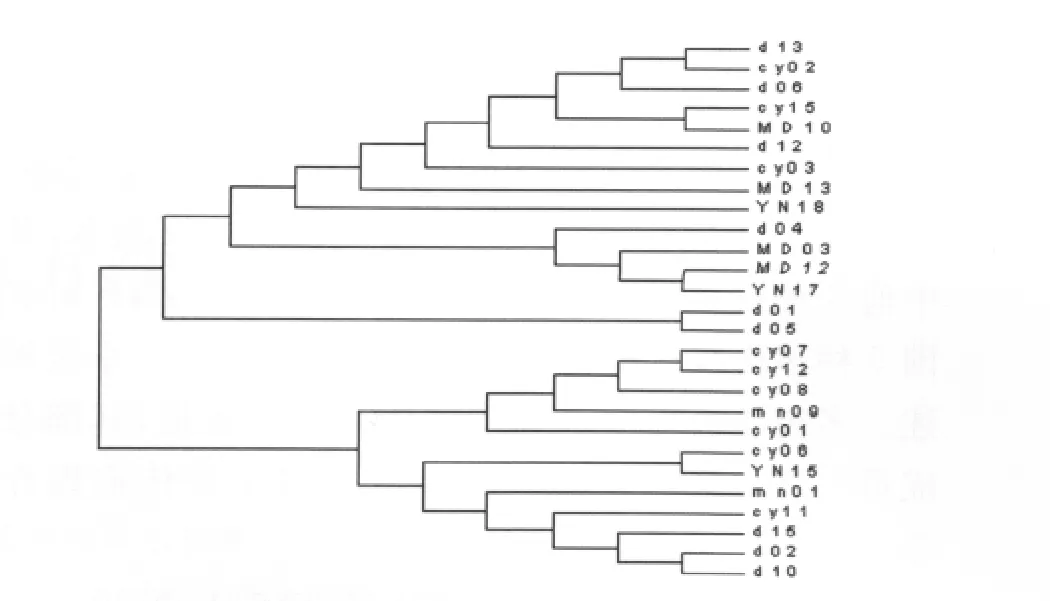
图2 伪威氏按蚊mtDNA-Cytb基因单倍型之间的聚类关系Fig.2 Cluster relation of mtDNA-Cytb haplotypes of Anopheles pseudowillimori

表2 伪威氏按蚊群体mtDNA-Cytb序列基本数据摘要Tab.2 mtDNA-Cytb gene data of Anopheles pseudowillmori population
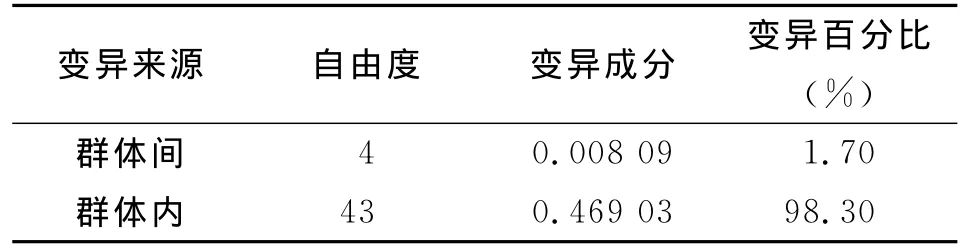
表3 伪威氏按蚊群体mtDNA-Cytb基因AMOVA分析Tab.3 mtDNA-Cytb AMOVA analysis of Anopheles pseudowillmori population
3 讨 论
多斑按蚊(Anophelesmaculatus)由Theobald于1901年定名,模式产地为香港,隶属按蚊属,塞蚊亚属,新塞蚊系,主要分布于东洋区(Oriental region),从印度次大陆,经东南亚至中国台湾。多斑按蚊过去一直被认为是单一蚊种,后经科学家多年研究证实,发现多斑按蚊是由多个形态相似的蚊种组成的复合体,多斑按蚊复合体在我国主要分布于北纬34°以南地区,是山区、半山区及河谷地区的主要蚊种。目前该复合体已经包括9种,Harbach[6-7]最终确立多斑按蚊复合体有9成员种,即多斑按蚊(An.maculatus)、威氏按蚊(An.willmori)、伪威氏按 蚊 (An.pseudowillmori)、塞 沃 按 蚊 (An.sawadwongporni)、达罗毗按蚊(An.dravidicus)、异形按蚊(An.dispar)、诺他按蚊(An.notanandai)、格林按蚊(An.greeni)和多斑按蚊核型 K,其中前5种在我国有分布。姬淑红[8]和陆宝麟[9]对我国5种多斑按蚊复合体的形态鉴别特征进行系统阐述。多斑按蚊复合体成员种形态非常相似,其部分成员种是马来西亚、泰国、老挝等国主要传疟媒介之一[10-12]。
课题组前期研究在西藏疟疾流行区墨脱县发现,伪威氏按蚊为该地的主要传疟媒介,云南省与西藏自治区相邻,董学书[13]发现多斑按蚊复合体中的伪威氏按蚊、威氏按蚊和多斑按蚊在云南省为全省分布,占云南山区、半山区和河谷区按蚊总数的42.68%,其中伪威 氏占 22.44%,多斑 按蚊占9.35%,甚至在云南省某些地区多斑按蚊复合体占捕获按蚊的比例高达94.6%,为绝对优势蚊种[14]。并经流行病学调查发现,多斑按蚊与伪威氏按蚊和疟疾发病关系密切,推测可能为传疟媒介;但都因未能发现子孢子感染的直接证据,未能确定其为传疟媒介。本研究从群体遗传学的角度,探讨了西藏疟疾流行区墨脱县、察隅县与云南省勐腊、景洪和缅甸拉咱市伪威氏按蚊的群体遗传分化关系,发现各群体间基因交流频繁,尚未发生群体间的分化。另群体内变异(98.30%)远大于群体间变异(1.70%),提示伪威氏按蚊可能曾经历过近期群体扩张事件,且目前仍处于群体扩张阶段,故mt DNA-cytb基因序列的核苷酸变异主要发生在于群体内个体间研究,结果也暗示了云南省的伪威氏按蚊理论上也应具备疟疾的传播能力,应该加强云南省伪威氏按蚊传疟能力的研究。
[1]武松,潘嘉云,王学忠,等.西藏墨脱县疟疾流行区多斑按蚊复合体种型鉴定[J].中国寄生虫学与寄生虫病杂志,2008,26(4):286-289.
[2]武松,黄芳,张国庆,等.巢氏PCR判定西藏疟疾流行区传疟媒介[J].中国人兽共患病学报,2010,26(7):648-650,653.
[3]Kambhampati S,Black WC,Rai KS.Random amplified poly morphic DNA of mosquito species and population(Diptera:Culicidae):techniques,statistical analysis,and applications[J].Med Entomology,1992,29(6):939-945.
[4]Beard CB,Hamm DM,Collins FH.The mitochondrial genome of the mosquito Anopheles gambiae:DNA sequence,genome organization,and comparisons with mitochondrial sequences of other insects[J].Insect Mol Biol,1993,2(2):103-124.
[5]Das NG,Bhuyan M,Das SC.Entomological and epoidemiological studies on malaria Rajmahal range,Bihar[J].Lndian J Malariol,2000,37(3/4):88-96.
[6]Harbach RE.The classification of genus Anopheles(Diptera:Culici-dae):a working hypothesis of phylogenetic relationships[J].Bull Entomol Res,2004,94(6):537-553.
[7]Walton C,Somboon P,o’Loughlin SM,et al.Genetic diversity and molecular identification of mosquito species in the Anopheles maculatus group using the ITS2 region of r DNA[J].Infection Genetics and Evolution,2007,7:93-102.
[8]姬淑红,陆宝麟.中国多斑按蚊类群小记[J].动物分类学报,1991,16(2):224-227.
[9]陆宝麟.中国动物志 昆虫纲 第九卷 双翅目 蚊科(下卷)[M].北京:科学出版社,1997:88,92,99,102,106.
[10]Rahman WA,Adanan CR,Abu Hassan A.Species composition of adult Anopheles populations and their breeding habitats in Hulu Perak district,Peninsular Malaysia[J].Southeast Asian J Trop Med Public Health,2002,33(3):547-550.
[11]Kengluecha A,Singhasivanon P,Tiensuwan M,et al.Water quality and breeding habitats of anopheline mosquito in northwestern Thailand[J].Southeast Asian J Trop Med Public Health,2005,36(1):46-53.
[12]Vythilingam I,Phetsouvanh R,Keokenchanh K,et al.The prevalence of Anopheles(Diptera:Culicidae)mosquitoes in Sekong Province,Lao PDR in relation to malaria transmission[J].Trop Med Int Health,2003,8(6):525-535.
[13]董学书.云南多斑按蚊种团的地理分布、生态习性与疟疾的关系[J].寄生虫与医学昆虫学报,1996,3(2):100-105.
[14]和春桐.云南贡山县独龙河谷地带传疟媒介的调查研究[J].现代预防医学,2008,35(23):4670-4671.
猜你喜欢
自我保健(2021年8期)2021-12-01
红河学院学报(2021年4期)2021-11-19
矿产勘查(2020年5期)2020-12-19
中华养生保健(2020年5期)2020-11-16
建材发展导向(2020年16期)2020-09-25
世界农药(2019年3期)2019-09-10
——“零疟疾从我开始”
中华灾害救援医学(2019年5期)2019-01-21
中国病理生理杂志(2018年6期)2018-01-22
今日民族(2017年1期)2017-02-15
中华老年口腔医学杂志(2016年3期)2017-01-15
