
芒果不同部位挥发物及横线尾夜蛾对其触角电位反应
2012-10-12 08:13田厚军陈艺欣邱良妙刘其全占志雄
生物安全学报 2012年2期
田厚军,陈艺欣,魏 辉,邱良妙,刘其全,石 妍,占志雄
福建省农业科学院植物保护研究所,福建福州350013
芒果横线尾夜蛾Chlumetia transverse Walker又名蛀梢夜蛾、钻心虫,属鳞翅目夜蛾科。在我国分布于广东、广西、海南、云南、福建和台湾等省,国外芒果产区如泰国和印度也有广泛分布。由于芒果经济价值较高,近年来大量种植。芒果横线尾夜蛾幼虫蛀食芒果嫩梢和花穗,引起嫩梢枯死、花序顶部丛生或枯死,影响果树的正常生长发育和产量。据初步调查,嫩梢的受害率在35%以上,花穗受害率在25%以上(何林和杨永生,2003)。目前,对该虫的防治基本依赖化学农药(陈业渊等,2001;陈永森等,2009;高松峰等,2000;彭正强等,2008),既污染环境,又容易产生抗药性。
目前,国内外已对多种鳞翅目昆虫的触角感觉器进行了表面细微结构的研究(王桂荣等,2002;杨广等,2001;Hallberg et al.,1994),但对横线尾夜蛾研究较少,且多为发生为害以及触角电镜扫描等的初步描述(冯荣扬,1997;何林,2003;莫圣书和赵冬香,2006)。有关其雌、雄成虫生理生化特征、幼虫为害特性、性信息素、寄主选择机理等均未见报道。周志军和王世贵(2005)证实,横线尾夜蛾触角感觉器是接受外界化学信息的主要结构,具有嗅觉、触觉和听觉等功能(刘玉双和石福明,2005;Zacharuk,1980)。
植物挥发性次生物质是昆虫借以寻找寄主或产卵场所的一类它感信息化合物,调控着昆虫对寄主植物的定向、识别和取食等行为(杜家纬,2001)。不同昆虫对植物挥发性物质的感受能力有着显著差异。许多植物在特定时间和空间都会产生具有各自特性的气味,植食性昆虫以这些气味为线索准确寻找和定位寄主植物(Bruce et al.,2005)。以棉铃虫Heliothis armigera(Hubner)为例,寄主植物信号物质中的六碳醇和醛能引起其雌、雄虫触角电位反应(electroantennogram,EAG)(Chen et al.,1997)。本研究采用电生理方法,测定芒果横线尾夜蛾雌、雄成虫对12种芒果挥发物的触角电位反应,为从化学信号角度阐明其寄主定向和雌、雄交配机理,探讨受害器官与果实、植物天然化学信号物质、害虫三者之间的关系,阐明害虫致害机理提供依据。
1 材料与方法
1.1 材料
横线尾夜蛾幼虫或成虫采集于福建省福清市,于福建省植物保护研究所饲养室用嫩叶、嫩梢饲养。化蛹后用指形管单只分装,羽化后分别取2日龄未交配雌、雄蛾进行电生理行为测定;化蛹后挑取1对蛹分装于同一只指形管中,羽化并交配后用于电生理行为测定。蛹和成虫饲养温度(25±1.0)℃,光照时间16 h·d-1,相对湿度65% ~75%。
根据本试验测得以及已报道的芒果挥发物种类及结构特点,选取具有代表性的信息化合物12种:乙酸乙酯(ethyl acetate)为分析纯,购自天津市科密欧化学试剂开发中心;纯度分别为≥85.0%和≥95.0%的异松油烯(terpinolene)和 α-水芹烯(αphellandrene)购自 Fluka公司;纯度分别为≥95.0% 、≥98.5% 、≥97.0% 、≥70.0% 、≥85.0% 、90.0% 、99.0% 、≥98.0% 、99.0% 、98.0% 的 β-香叶烯(β-myrcene)、β-石竹烯(β-carypohyllene)、香橙烯(aromadendrene)、朱栾倍半萜(valencene)、α-法呢烯(α-farnesene)、(+)-3-坎烯[(+)-3-carene]、α-萜品烯 (α-terpinene)、反-2-己烯-1-醛 (trans-2-hexen-1-al)、(1R)-(+)-α-蒎烯[(1R)-(+)-α-pinene]、顺-3-己烯醇(Z-3-hexen-1-ol)(作为参照)购自Sigma Aldrich公司。石蜡油(paraffin)购自北京化工厂。
SPME 萃取柄(Supleco公司),100 μm 聚二甲基硅氧烷(PDMS)萃取头(Supleco公司),Saturn 3900/2100气相色谱—质谱联用仪(美国Varian公司)。触角电位仪(荷兰Syntech公司)主要包括毛细管电极(内径0.2 mm)、微动操作仪(Syntech MP-15)、直流/交流放大器(Syntech UN-06)和刺激气流控制装置(Syntech CS-05)。
1.2 GC-MS 分析条件
色谱条件:美国Varian Saturn 3900/2100气相联用仪(GC-MS),采用DB-5色谱柱(柱长30 m,内径 0.25 mm,液膜0.25 μm),进样口温度250 ℃,起始柱温 80℃,保持 5 min,以 4℃·min-1升至200℃,保持2 min,以15℃·min-1升至260℃,恒温至色谱分析结束。载气为氦气,纯度>99.999%。流速1 mL·min-1,不分流进样。
质谱条件:电离方式EI,电子能量70 eV,阱温220℃,传输线温度170℃。GC-MS接口温度280℃。采集方式为全扫描,质量扫描范围为40~650 amu。
1.3 挥发性气体收集
使用前将SPME萃取头旋入萃取柄,用无水乙醇浸泡萃取头1 h后,将萃取头在气相色谱仪进样口270℃下老化30 min。将不同生长期的芒果梢、花和果实分别放于小型干燥器内,30 min后驱赶干燥器内空气,然后用无味的透明薄膜封口。将SPME萃取头插入薄膜内吸附挥发物,30 min后用于GC-MS进样。
1.4 触角电位测定方法
触角电位测定方法参照田厚军等(2011)的研究。将20 μL样品溶液均匀滴在长5 cm、宽0.5 cm的“V”字形滤纸条上,放入巴斯德滴管中,滴管末端连接刺激气流控制装置,送气管管口与触角纵向垂直,并相距 1 cm。调节连续气体流量为124 mL·min-1,刺激气体流量为20 mL·min-1,每次刺激时间为0.2 s。2次刺激之间间隔40 s,以保证触角感觉器官功能完全恢复。同一化合物同一剂量在1根触角上平行刺激3次,取平均值,每个样品测试5根触角。对于每种样品化合物,刺激顺序为参照、样品、参照,前后2次参照取平均值。以液体石蜡油为溶剂并作为对照,将12种化合物配制成浓度均为10 μg·μL-1的溶液。每种化合物的EAG 相对反应值以 10 μg·μL-1的顺-3-己烯醇为参照。
EAG相对反应值=(待测样品的反应值-对照反应值)/(参照物的平均反应值-对照反应值)×100%
1.5 数据分析
芒果挥发物成分通过普图库(NIST05和WILEY7)进行计算机检索并辅以人工识别确定化合物种类,按照面积归一化法计算各化合物的相对含量,定量分析在Excel软件上进行。采用SPSS 17.0统计软件进行方差分析,数据采用平均值±标准误(SE)表示,挥发物峰面积以及相对含量差异性、EAG反应显著差异性采用Duncan's新复极差检验法(SSR,P=0.05)进行分析比较。
2 结果与分析
2.1 芒果不同部位挥发性物质的化学成分
芒果3个部位的挥发性物质共检测到29种,且多为萜烯类化合物。芒果花挥发物有22种,含量较高的为 α-水芹烯(43.64%)和异松油烯(29.33%);芒果梢有11 种,α-古芸烯(23.63%)含量最高;成熟芒果有13种,石竹烯含量最高,为29.47%。仅异松油烯和β-荜澄茄油烯在芒果3个部位中均被检测到,但含量存在差异。其中,异松油烯在芒果3个部位挥发物中的含量分别为29.33%、19.99%、0.59%,可见同一物质在不同部位挥发物中的含量差异较大;而β-荜澄茄油烯在芒果花中的含量为3.51% ,梢中为 2.18%,果实中为 6.05%(表 1)。
2.2 横线尾夜蛾成虫对芒果挥发性物质的触角电位反应
芒果横线尾夜蛾未交配雌、雄蛾与已交配雌、雄蛾对同一种化合物的EAG相对值之间均无显著差异(P>0.05)。未交配雌蛾对反-2-己烯醛的EAG反应值与其他化合物相比差异显著(P<0.05)。而异松油烯、香橙烯、乙酸乙酯、β-香叶烯、(+)-3-莰烯、α-水芹烯之间,β-石竹烯、α-法呢烯、(1R)-(+)-α-蒎烯之间,(+)-3-莰烯、朱栾倍半萜、β-香叶烯之间,朱栾倍半萜和α-水芹烯之间均无显著差异(P>0.05)。其余化合物之间则差异显著(P<0.05)。未交配雄蛾对反-2-己烯醛的EAG反应值与其他化合物相比差异显著(P < 0.05)。(1R)-(+)-α-蒎烯和 α-萜品烯之间,(+)-3-莰烯和α-水芹烯之间,香橙烯、朱栾倍半萜、异松油烯之间均有显著差异;α-萜品烯和α-法呢烯之间、α-萜品烯和 β-石竹烯之间、α-萜品烯和香橙烯之间也差异显著(P<0.05)。其余化合物之间则无显著差异(P>0.05)(表2)。
交配雌蛾对反-2-己烯醛的EAG反应值与其他化合物相比有显著差异(P<0.05)。而β-石竹烯、α-法呢烯、(1R)-(+)-α-蒎烯之间,(+)-3-莰烯、朱栾倍半萜、β-香叶烯之间,异松油烯、香橙烯、乙酸乙酯、β-香叶烯、(+)-3-莰烯、α-水芹烯之间,以及朱栾倍半萜和α-水芹烯之间均无显著差异(P>0.05)。其余化合物之间则差异显著(P<0.05)。交配雄蛾对反-2-己烯醛的EAG反应值与其他化合物相比有显著差异(P<0.05)。且 α-萜品烯和α-法呢烯之间、α-萜品烯和 β-石竹烯之间,α-萜品烯和香橙烯之间、(1R)-(+)-α-蒎烯和 α-萜品烯之间,(+)-3-莰烯和 α-水芹烯之间,香橙烯、朱栾倍半萜、异松油烯之间差异也均显著(P<0.05)。其余化合物之间则无显著差异(P>0.05)(表2)。

表1 芒果不同部位挥发物主要化学成分及其相对含量Table 1 Comparison of main constituents and their relative content of main volatiles in different parts of mango plants
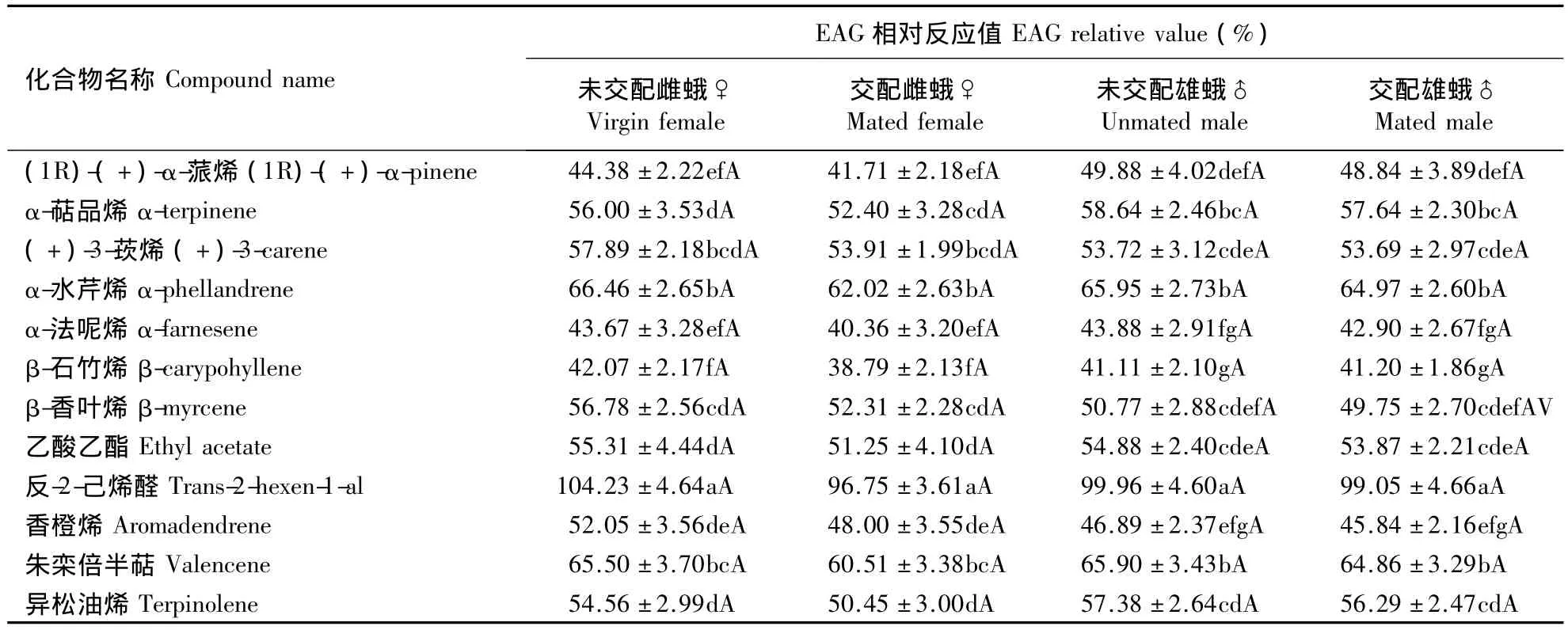
表2 横线尾夜蛾成虫对芒果挥发性物质的EAG相对反应值Table 2 Relative EAG amplitude of C.transverse to mango volatiles
3 结论与讨论
本研究结果表明,芒果果实、花、梢不仅化学组分的数量不同,而且各组分的相对含量差异也较大。仅异松油烯和β-荜澄茄油烯在3个部位均被检测到,但相对含量差别较大。异松油烯在芒果花和果实中的含量分别为29.33%和0.59%。芒果梢中 α-愈创木烯(9.23%)和 α-古芸烯(23.63%)含量远高于芒果花和果实,这是否与横线尾夜蛾幼虫偏爱蛀食芒果嫩梢有关还有待于趋性行为试验的验证。本研究检测到的芒果挥发物成分与文献中报道的化合物(何方奕等,2008;施伟等,2010;郑华等,2008)大多相符,主要为异松油烯、β-香叶烯、β-石竹烯、香橙烯、α-水芹烯等。本研究中芒果不同部位挥发物的种类并不丰富,可能与芒果品种以及不同的地理环境有关。
本试验结果还表明,未交配雌、雄蛾、交配雌、雄蛾对同一种化合物的EAG反应值无显著差异(P>0.05)。这说明雌、雄蛾交配前后对同一种化合物的敏感性并没有变化,且雌、雄蛾对其有相似的嗅觉感受能力(Raguso et al.,1996);同时也说明交配对横线尾夜蛾雌、雄蛾的触角电位反应影响较小。未交配雌、雄蛾与交配雌、雄蛾对反-2-己烯醛的EAG相对反应值(100%左右)远高于其他化合物,说明反-2-己烯醛对横线尾夜蛾可能有较高的趋性行为生物活性。EAG相对反应值较大的还有α-水芹烯(约65%)和朱栾倍半萜(约64%),相对反应值最小的则为β-石竹烯(约40%),但除反-2-己烯醛外,其他各化合物之间均无显著差异(P>0.05);在所测化合物中,EAG相对反应最高值约为最低值的2.5倍,而反应次高值约为最低值的1.6倍。这些化合物是否具有生物活性还有待于室内定向行为和室外田间行为试验的验证。
目前,利用植物源挥发物和性信息素之间的互作关系调控昆虫的行为机制已取得一定进展(Reddy&Guerrero,2004)。王振华等(2008)报道了植物源挥发物对昆虫性信息素的增效作用及其增效机制。苏茂文和张钟宁(2007)利用植物源挥发物对害虫的引诱作用成功防御了害虫。但横线尾夜蛾生活周期较长,室内饲养条件不成熟,仅靠野外采样既费时又费力。因此,建立一套成熟、可行的室内饲养方法尤为重要。同时,植物天然信号物质的化学结构复杂,且组成比例不同结果也不同,而目前的研究仍处于初步探索阶段,要真正提高其对昆虫性信息素的增效作用还需大量的工作。后续研究将关注横线尾夜蛾生理生化行为以及性信息素鉴定和性诱剂开发等方面,以探讨芒果挥发物与横线尾夜蛾性信息素之间的信号联系机理。
陈业渊,李绍鹏,高爱平,冯瑞祥,王家保,魏守兴.2001.无公害食品芒果生产技术规程.北京:中国农业出版社.
陈永森,黄国弟,莫永龙,周俊岸,赵英,梁宏合,蒲金基.2009.四种杀虫剂防治芒果横线尾夜蛾药效试验.广西农业科学,40(10):1312-1314.
杜家纬.2001.植物—昆虫间的化学通讯及其行为控制.植物生理学报,27(3):193-200.
冯荣扬.1997.粤西地区芒果横线尾夜蛾的发生规律及其防治研究.湛江海洋大学学报,17(2):71-74.
高松峰,李晓伟,黄木庭,张毅俊,钟卫国,吴松浩.2000.百虫净防治杧果横线尾夜蛾药效试验.中国南方果树,29(4):37.
何方奕,李铁纯,梁多壮,回瑞华.2008.芒果皮中挥发性成分的GC-MS分析.食品科学,29(10):495-496.
何林.2003.芒果横线尾夜蛾的生活习性及防治.昆虫知识,40(1):83-84.
何林,杨永生.2003.芒果横线尾夜蛾的发生危害及其防治.植物保护,29(2):43-44.
刘玉双,石福明.2005.红缘吉丁(鞘翅目:吉丁虫科)触角感器的扫描电镜观察.昆虫学报,48(3):469-472.
莫圣书,赵冬香.2006.芒果横线尾夜蛾触角感觉器扫描电镜观察.华东昆虫学报,15(2):96-98.
彭正强,韩冬银,符悦冠,刘奎,张方平,黄武仁,张敬宝.2008.芒果病虫害防治技术规范.北京:中国农业出版社.
施伟,刘辉,叶辉.2010.桔小实蝇对五种芒果气味挥发性物质的行为反应.昆虫知识,47(2):318-321.
苏茂文,张钟宁.2007.昆虫信息化学物质的应用进展.昆虫知识,44(4):477-485.
田厚军,陈艺欣,魏辉,占志雄,黄玉清,邱良妙.2011.小菜蛾成虫对9种挥发性物质的电生理反应.福建农业学报,26(4):591-595.
王桂荣,郭予元,吴孔明.2002.棉铃虫触角感器的超微结构观察.中国农业科学,35(12):1479-1482.
王振华,赵晖,李金甫,曾宪东,陈建军,冯汉利,徐家文.2008.植物源挥发物对昆虫信息素的增效作用及其增效机制.应用生态学报,19(11):2533-2537.
杨广,黄贵诚,尤民生.2001.小菜蛾触角的显微结构及其作用.福建农业大学学报,30(1):75-79.
郑华,张弘,张汝国,于连松,陈华君.2008.云南特产“小三年”芒果不同成熟期香气成分的差异.食品科学,29(10):487-490.
周志军,王世贵.2005.二化螟盘绒茧蜂触角感器的超微结构.昆虫知识,42(6):676-680.
Bruce T J,Wadhams L J and Woodcock C M.2005.Insect host location:a volatile situation.Trends in Plant Science,10:269-274.
Chen X,Hou Z Y,Zhang Y,Yan F S and Zhang G X.1997.Olfactory response of cotton bollworm,Helicoverpa armigera and plant volatiles components.Journal of Insect Science,4:159-172.
Hallberg E,Hanssonn B S and Steinbrecht R A.1994.Morphological characteristics of antennal sensilla in the European cornborer,Ostrinia nubilalis(Lep.:Pyralidae).Tissue and Cell,26:489 -502.
Raguso R,Light D and Pichersky E.1996.Electrantennogram responses of Hyles lineata(Lepidoptera:Sphingidae)to volatiles compounds from Clarkia breweri(Onagraceae)and other moth-pollinated flowers.Journal of Chemical Ecology,22:1735-1766.
Reddy V P and Guerrero A.2004.Interactions of insect pheromones and plant semiochemicals.Trends in Plant Science,9:253-261.
Zacharuk R Y.1980.Ultrastructure and function of insect chemosensilla.Annual Review of Entomology,25:27 -47.
猜你喜欢
阅读(快乐英语中年级)(2022年3期)2022-03-30
阅读(快乐英语中年级)(2021年10期)2021-03-08
四川蚕业(2021年3期)2021-02-12
新农业(2020年18期)2021-01-07
昆虫学报(2020年6期)2020-08-06
阅读(快乐英语高年级)(2020年8期)2020-01-08
小小艺术家(2018年9期)2018-11-23
蚕学通讯(2017年3期)2017-03-23
小小说大世界(2014年8期)2014-09-25
长江蔬菜(2014年1期)2014-03-11
