
小兴安岭十种典型森林群落凋落物生物量及其动态变化
2013-09-07 02:53侯玲玲毛子军
生态学报 2013年6期
侯玲玲,毛子军,孙 涛,宋 元
(东北林业大学森林生态学教育部重点实验室,哈尔滨 150040)
森林生态系统作为陆地上分布最广,面积最大,最重要的自然生态系统,其生物量和净生产力占整个陆地生态系统的86%和70%,每年固定的碳约占整个陆地生态系统的2/3,对于调节全球碳平衡方面具有不可替代的作用[1-3]。凋落物作为森林生态系统的重要结构组成,是物质循环和能量流动的重要途径,在某种程度上反映了森林初级生产力[4-5]。凋落物可以通过养分循环使大量有机质归还林地,改善土壤的理化性质和生物学特性,增加土壤肥力[6]。凋落物层还可以起到保温和水文生态作用[7-8]。目前对于森林凋落物的研究多集中南方地区,而对东北地区不同森林类型的研究较少[9],对凋落物组成的分类也较粗放,多分为枝,叶,杂物3类,不能反映出林分凋落物的组分特征[10]。对年均凋落量等基础数据的研究还缺乏连续性,全面性和完整性。
小兴安岭作为我国重要林区之一,在我国森林生态系统中占据举足轻重的地位,目前国内对小兴安岭森林生态系统凋落物的研究比较零散,大多针对顶级群落阔叶红松林[9,11-12]或个别天然次生林[11,13],对该地区多种典型森林群落凋落物研究较少。本文通过对小兴安岭地区10种主要群落类型凋落物的研究,探讨了原始红松林和典型天然次生林的凋落物产量、组成特征、季节动态变化间的差异,为丰富我国东北地区森林生态系统凋落物的数据和资料,为小兴安岭森林生态系统碳储量和养分循环的研究提供科学参考。
1 实验地概况
本实验样地位于黑龙江丰林国家级自然保护区(48°02'N,128°58'—129°15E),研究区域地处小兴安岭南坡北段,森林覆盖率达95%,森林总面积18165hm2,人为干扰相对较小,植被类型较丰富,属于长白植物区系的小兴安岭亚系。全区为孤岛状的低山丘陵地形,海拔高度在285—688m之间。因所处纬度较高,年均温较低为-0.5℃,年均最低气温在1月份为-24.2℃,年均最高气温在7月为20.3℃,保护区的年均降水量640.55mm,且主要集中在6—9月份,年蒸发量为930mm,无霜期为100—110d。本区海拔变化幅度小没有明显高山,地带性土壤为暗棕色森林土,植被类型是以红松为主的温带阔叶红松混交林,占总面积的78%。天然次生林以白桦次生林、白桦落叶松林、山杨林、枫桦林、杂木林等为主,占保护区总面积的13.4%。该区主要树种组成有兴安杜鹃(Rhododendron dauricum)、胡枝子(Lespedeza bicolor)、枫桦 (Betula costata)、紫椴(Tilia amurensis)、山杨(Populus davidiana)、东北山梅花 (Philadelphus schrenkii)、光萼溲苏(Deutzia glabrata)、青楷槭(Acer tegmentosum)、花楷槭 (Acer ukurunduense)、臭冷杉(Abies nephlolepis)、红皮云杉(Picea koraiensis)、水曲柳(Fraxinus mandshurica)、刺五加(Acanthopanax senticosus)鱼鳞云杉(Picea jezoensis)、红松(Pinus koraiensis)、白桦(Betula platyphylla)、蒙古栎(Quercus mongolica)、落叶松(Larix gmelini)等。
本研究在小兴安岭地区10种典型森林群落内设置试验样地各一块分别为:蒙古栎红松林(QP)、枫桦红松林(BP)、椴树红松林(TP)、云冷杉红松林(PAP)、火烧迹地白桦天然次生林(SB)、采伐迹地白桦落叶松次生林(BL)、云冷杉林(PA)、山杨次生林(SP)、枫桦次生林(SC)、杂木次生林(SCB),试验样地面积为20m×20m,各群落的主要结构特征见表1。
2 研究方法
2.1 凋落物收集方法
凋落物的收集采取直接收集法。在所选的10种林分中各设置一块20m×20m的样地,随机放置8个凋落物收集器,收集器为用40目尼龙网缝制成圆锥形的网兜(这样可以防止强风将收集器中的凋落物刮出影响实验结果),每个收集器面积0.46m2。收集2009和2010两年的凋落物。收集凋落物的时间在每年5—10月,每月收集1次,11月入冬大雪封山后,不便收集,则在第2年春季统一收集1次。每月初将收集器中的凋落物取回后,先将凋落物分为针叶、阔叶、杂物、枝、皮、花、果等7类,然后在65℃恒温下烘至恒重,分别进行称重。对每块样地8个收集器中的凋落物重量取平均值,即为该样地该月凋落物产量。
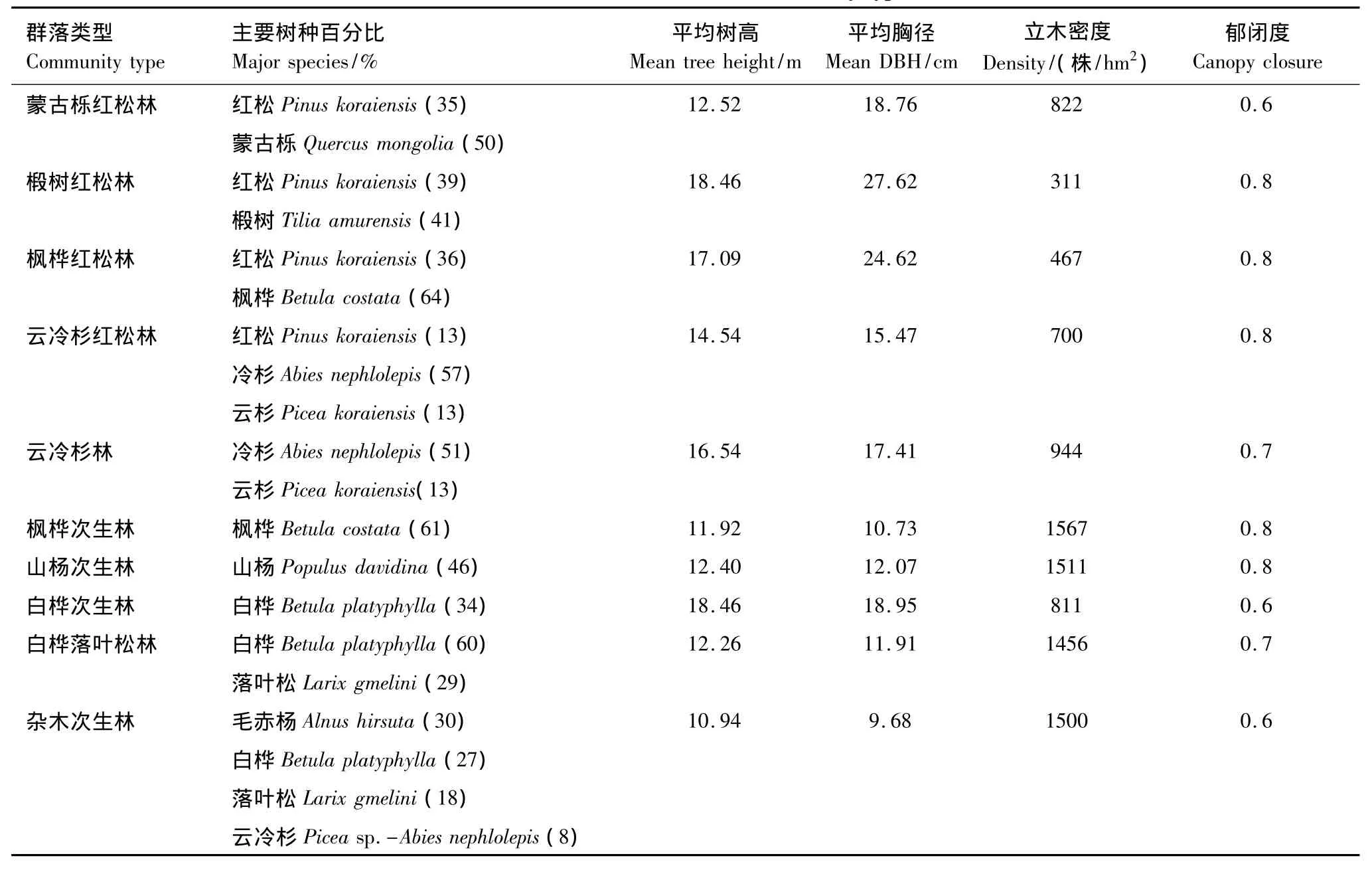
表1 10种群落的主要群落特征Table1 Characteristics of ten community types
2.2 统计分析方法
绘图采用Origin7.5软件,数据分析运用SPSS17.0软件中的One-way ANOVN进行方差分析,最小显著差异法(LSD)检验法分析不同林分凋落量的差异显著性。
3 结果分析
3.1 不同群落类型的年凋落量
凋落物的年均凋落量在10种群落类型间有一定差异(表2),总体上,原始林凋落量大于天然次生林。大小依次为椴树红松林(4.08t·hm-2·a-1)>蒙古栎红松林(3.83 t·hm-2·a-1)>云冷杉红松林(3.55t·hm-2·a-1)>云冷杉林(3.44t·hm-2·a-1)>枫桦红松林(3.43t·hm-2·a-1)>山杨次生林(3.26t·hm-2·a-1)>白桦次生林(3.04 t·hm-2·a-1)>枫桦次生林(2.96 t·hm-2·a-1)>杂木林次生林(2.95t·hm-2·a-1)>白桦落叶松林(2.91t·hm-2·a-1)。4种原始红松林中椴树红松林与枫桦红松林差异显著(P<0.05),与其他3种红松林无明显差异,5种天然次生林之间差异不显著。
3.2 凋落量的月动态变化
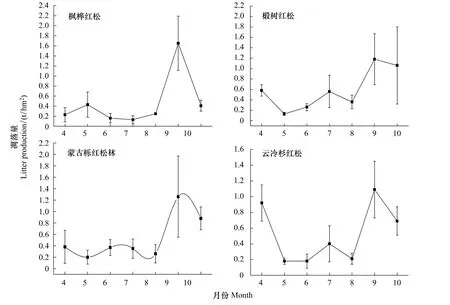
研究结果表明,森林凋落物有明显的季节变化规律,在所研究的10种群落中,枫桦红松林、椴树红松林、蒙古栎红松林、山杨、枫桦、白桦、杂木林为单峰型,云冷杉红松林、云冷杉林、白桦落叶松为双峰型。枫桦红松林从五月开始出现第1个小波峰,而凋落的主要高峰期出现在9月末。椴树红松林4月分凋落量较高,在7月末也有个峰值,而凋落量最高峰值则出现在9—10月份。蒙古栎红松林在4—7月份凋落物量变化趋势较平缓,而它的凋落量高峰期也主要集中在9—10月。云冷杉红松林、云冷杉林、白桦落叶松存在两个明显的凋落高峰,为双峰型凋落模式。从图1可以看出4—5月份常绿针叶林的凋落量急剧上升,形成了凋落第1次高峰,云冷杉红松林第2次高峰出现在9月份,云冷杉林的第2个凋落高峰在9—10月份,它们的两次峰值相差均较小。白桦落叶松两次峰值出现时间为8月和10月份,山杨、白桦、枫桦、杂木4种天然次生林的凋落模式尽管都为单峰型,但凋落量峰值出现时间有差异。山杨林和杂木次生林的凋落峰值在9月末,枫桦林凋落峰值在8—9月,白桦林的凋落高峰主要在8月份(图1)。
3.3 凋落物各组分产量
凋落物的各组分主要由叶(包括针叶和阔叶)、枝、花、果、皮、杂物(动物残体、芽鳞、碎削等统称)部分组成。本研究表明,不同群落类型的凋落物组分存在明显差异,即使是树种组成相同,凋落组分也会由于不同的群落而有差异。3种原始阔叶红松林的凋落物生物量中叶占主要部分,枫桦红松林针叶、阔叶所占比例各为27.6%和51%,其次是小枝和杂物各占总量的9.2%、6.4%,此群落中阔叶所占比重较大。椴树红松林和蒙古栎红松林针叶、阔叶各占总凋落量的49.6%、12.2%和50%、16.2%,花果次之,小枝再次之,这两种群落中针叶比重最大。常绿针叶林中云冷杉红松林和云冷杉林针叶所占比例分别为63.6%和62.9%,其余依次为阔叶(10.3%、15.4%),枝(8.9%、8%),杂物(8%、6.3%),花(6.3%、3.4%)等。落叶阔叶林的凋落物中阔叶的凋落量最高,山杨、枫桦、白桦林的阔叶分别占年凋落量的82%、82.4%和82.3%,小枝分别为总凋落量的9.4%、7.7%、8.3%,而其他组分较少。针阔混交次生林中的白桦落叶松林,其针叶和阔叶比例分别占总量的28%和60.4%,杂木林作为小兴安岭分布较广的群落类型,树种组成复杂,针阔叶所占比例分别为43.9%和37.6%(表3)。
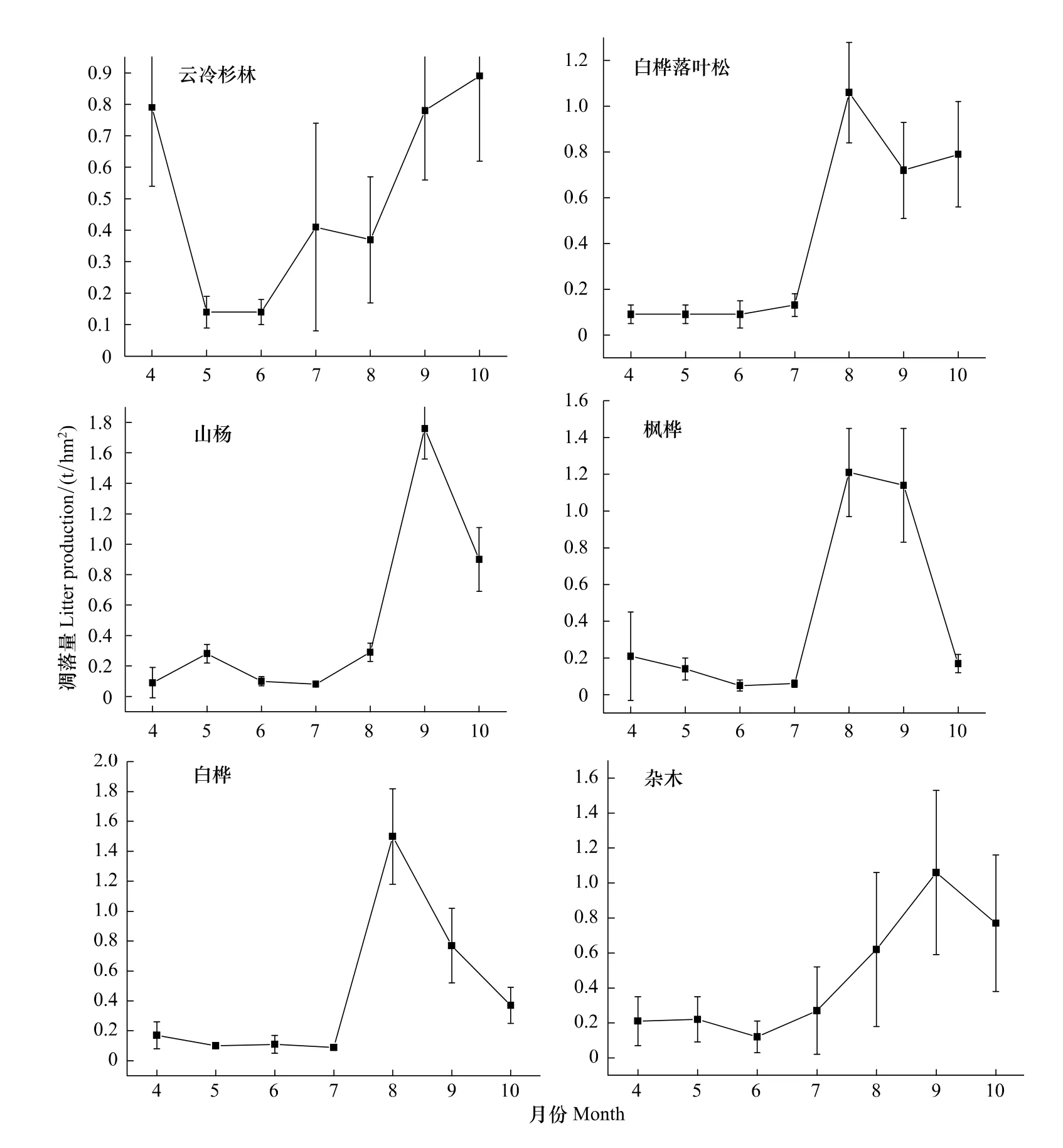
图1 小兴安岭10种群落类型凋落量月动态Fig.1 Monthly variation of litter fall production for ten community types in Xiaoxing'an Mountain
4 讨论与结论
4.1 各群落类型年凋落量差异
森林凋落物是养分循环和碳流动中的重要环节,凋落物产量能够反映森林群落的生态功能,对森林生态恢复和更新有重要作用[14]。本文研究表明四种原始红松林凋落量高于天然次生林,并且2009和2010两年内凋落物产量变化小(表2),与李雪峰在小兴安岭凉水自然保护区测得凋落物年均凋落量阔叶红松林>白桦林>落叶松林相一致[9]。一般来说针阔混交林要比单一林型的凋落量和养分归还量高[15],并且阔叶红松林的净初级生产力高于常绿针叶林和落叶阔叶林[16-17],相应的其凋落物产量也最高。原始红松林作为小兴安岭典型地带植被和森林生态系统的顶级群落[18],其群落结构稳定性较高,受外部因素干扰性小。本文研究的3种阔叶红松林中,椴树红松林凋落量显著大于蒙古栎红松林和枫桦红松林,这与群落主要特征有关,椴树红松林中红松的胸径较大(表1),而凋落物总量又与林分平均胸径成正相关(P<0.05),说明林分平均胸径对凋落物产量有一定影响(表4)。而从生境上看,该林分处于缓坡中下腹,土层较厚,且排水良好,是红松生长的最适宜生境,这也是该林分比分布在其上部的枫桦红松林和陡坡上部的蒙古栎红松林的生产力大的主要原因。因此在气候因素相同的条件下,群落类型也会影响凋落物产量,这与群落结构特征和具体生境有关。以往研究表明凋落物总量主要受年均温和森林类型的影响[9,11-19],本文通过实验得出三种阔叶红松林的凋落物产量也受种类组成和生境差异的影响。张新平等对长白山云冷杉林的研究发现其年均凋落量为2.472t/hm2[11],远小于本研究中的云冷杉林,这主要受地理条件和气候条件影响,使同一林型在不同地点凋落量有明显差异,因为长白山自然保护区云冷杉林海拔在1100—1800m[20],高于阔叶红松林分,环境条件比较严酷,寒冷,因此凋落量较小。本研究中的山杨次生林的凋落量低于帽儿山(4.27t·hm-2·a-1)[13],这主要是由于受纬度影响,两地年均温相差约3℃,进而影响凋落物产量[9]。这结果符合凋落物产量随海拔和纬度的升高而下降的规律。本文中90a林龄的白桦天然次生林年均凋落量高于40a林龄的白桦落叶松林,说明林龄也可能是影响凋落量的一个原因[21]。
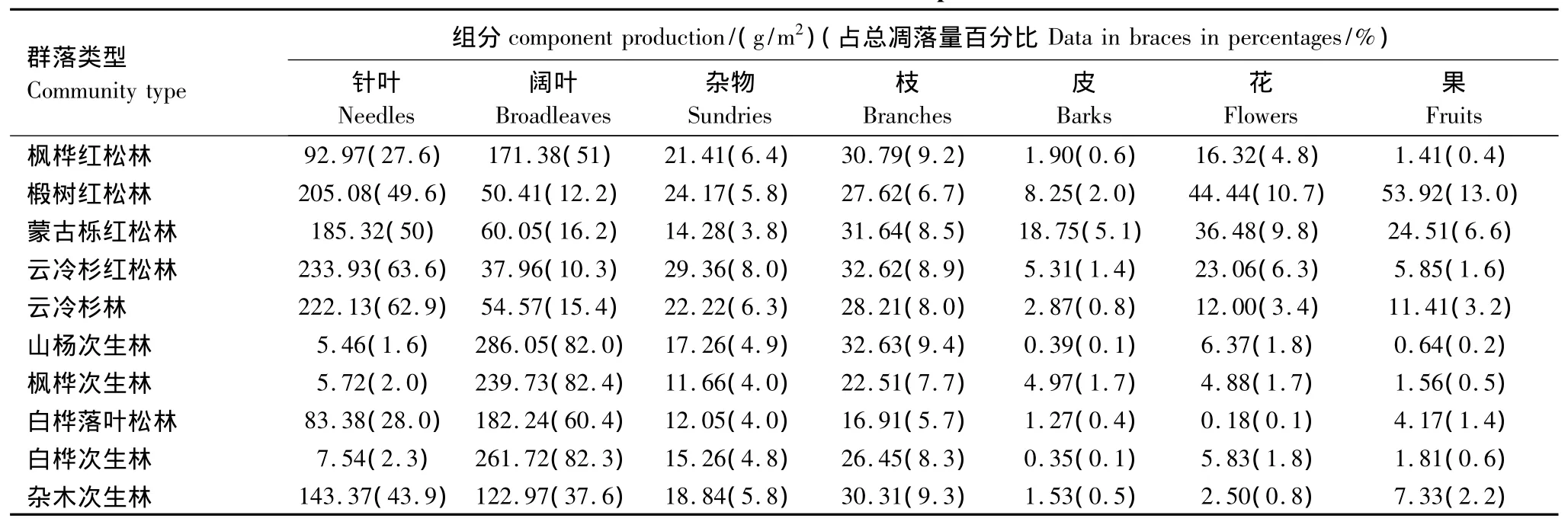
表3 10种群落类型凋落物组分产量Table3 Productions of different litter components in 2010
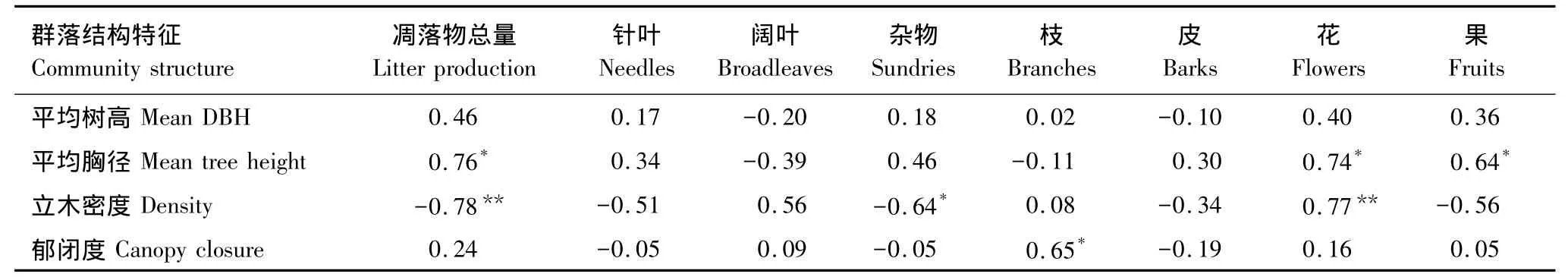
表4 凋落物总量及各组分与群落结构指标的相关系数Table4 Correlation between litter production,litter components and community structure
综上所述本文中10种群落类型的年凋落总量符合Bray统计的全球温带年凋落量3.1—4.9 t·hm-2·a-1的研究结果[22]。小兴安岭地区凋落物总量除了温度、地理条件、森林类型的影响[9,11,23],还受 NPP 值、群落类型、林龄[21]、树种生物学特性,群落结构组成等因素的影响。
4.2 各群落类型凋落量动态差异
本文中10种群落类型凋落物凋落动态分为2个不同类型:单峰型和双峰型。针阔混交林类型和阔叶次生林类型为单峰型,针阔混交林高峰期在9月份,但是在春季(4、5月份)也出现一个小峰值;阔叶次生林高峰期在8月份和10月份,主要是秋季落叶,这是因为在秋季生长停止后,随着气温降低,为减少养分和水分的消耗,形成大量生理性落叶[10],同一凋落模式的群落,由于群落组成不同其凋落高峰期也存在差异,白桦,枫桦8月份凋落量最高,山杨的凋落峰值则在9—10月份最高。因为枫桦和白桦的叶比山杨叶脱落早,这是由树种叶的物候性和耐受性决定的,枫桦和山杨叶的耐低温性不如山杨,这也体现了树种间物候性的不同[24]。针阔混交次生林和常绿针叶林为双峰型,白桦落叶松高峰分别在8月和10月,这是因为白桦叶片在8月就大量凋落,而落叶松针叶凋落峰值在10月份。云冷杉红松林和云冷杉林高峰期分别在9月份和4月份。第2个高峰均在4月原因是由于该地区11月份已进入冬季以后下雪后山路已封,不能每月收集凋落物,所以翌年4月份的凋落物实际上是11—4月份共6个月的,但也可看出,针叶林冬季凋落量远高于针阔混交林和次生林,而这期间主要的凋落物成分为针叶、枝和杂物,即在环境条件基本相同条件下,由于小兴安岭冬季风雪较大,导致常绿针叶林大量针叶枝的大量脱落。而4、5月份由于气温回升,树种发出新叶,老叶缺乏养分而脱落,出现短暂集中的换叶期,形成了凋落的第2次高峰[25]。
本文凋落物研究的季节动态与以往研究的结果相似,常绿森林的凋落量季节动态模式是双峰型,而阔叶落叶林其凋落模式一般是单峰型[26]。因此通过研究可知凋落物的季节动态受气候条件和树种组成生物特性的影响[27],不同生物特性的树种,因为对环境条件的耐受性不同,也会影响凋落模式。
4.3 各群落凋落物组成差异
10种群落中落叶占总凋落量的比例最大,叶片是进行光合作用,为树木生长制造养分的主要器官,而叶的凋落量受温度限制明显[11],因此秋季温度降低后针叶和阔叶大量凋落。原始红松林和云冷杉林的凋落物组分中除了针叶外,枝和花、果占主要部分,主要树种红松、云杉、冷杉生长季产生大量花序,而秋季球果脱落,使该组分所占总凋落量的比例增加。可见常绿针叶林用于繁殖的器官资源分配较大。值得指出的是,产量最大的红松的球果基本没有包括在凋落物中。因为每年秋季红松球果均被人工采摘。阔叶树种的养分主要用于营养器官的生长,所以山杨、枫桦、白桦落叶占80%以上,而花果等繁殖器官比例极小,这是由于白桦、枫桦种子质量轻。枯落枝的比例较大,因为北方冬季寒冷易受风雪侵袭,枝条进行大量的代谢脱落,能够减少树木蒸腾、保存养分[28]。针、阔叶树种存在较大的生理生态学特性差异造成生长分配策略有着较大不同,如用于光合的叶片、用于防御的枝条、用于繁殖的花果之间的比例不同[11,29]。3种阔叶红松林凋落叶中针阔叶所占比例不同,椴树红松林和蒙古栎红松林针叶比例大于阔叶,枫桦红松林中阔叶比例高于针叶,这主要与群落中针叶树和阔叶树的组成比例不同有关,枫桦红松林中枫桦占64%(表1),而其他2个群落中阔叶树种只占21%和30%。因此不同群落类型的凋落物各组分不仅受树种的生物学特性影响还受群落结构组成影响。
总之,小兴安岭原始阔叶红松林比天然次生林的年均凋落量大,群落结构稳定,能更好地进行养分归还,因此保护原始红松林,促进天然次生林向顶级阔叶红松林群落的演替,对提高整个森林生态系统的碳汇能力和生态恢复有深远意义。
[1] Zhou Y R,Yu Z L,Zhao SD.Carbon storage and budget of major Chinese forest types.Acta Phytoecologica Sinica,2000,24(5):518-522.
[2] Wang X K,Feng Z W,Oyang Z Y.Vegetation carbon storage and density of forest ecosystems in China.Chinese Journal of Applied Ecology,2001,12(1):13-16.
[3] Liu G H,Fu B J,Fang J Y.Carbon dynamics of Chinese forests and its contribution to global carbon balance.Acta Ecologica Sinica,2000,20(5):733-740.
[4] Wang SL.Litter ecology,Beijing:Science Press,2010,1(3):14-15.
[5] Peng SL,Liu Q.The dynamic of forest litter and its responses to global warming.Acta Ecologica Sinica,2002,22(9):1534-1544.
[6] Guo W,Zhang J,Huang Y M,Liu X,Wang W,Xue L.Research progress on ecological function of forest litter.Journal of Anhui Agri.Sci.2009,37(5):1984-1987.
[7] Pan K W,He J,Wu N.Effect of forest litter on microenvironment conditions of forestland.Chinese Journal of Applied Ecology,2004,15(1):153-158.
[8] Zhu J Z,Liu JJ,Zhu Q,Wu QX.Hydro-ecological functions of forest litter layers.Journal of Beijing Forestry University,2002,24(5/6):30-34.
[9] Li X F,Han SJ,Li Y W,Hou B Z,Li X L.Litter fall in main forest ecosystems of Northeast China.Chinese Journal of Applied Ecology,2005,16(5):783-788.
[10] Yuan Z Q,Li B H,Bai X J,Lin F,Shi S,Ye J,Wang X G,Hao Z Q.Composition and seasonal dynamics of litterfalls in a broad-leaved Korean pine(Pinus koraiensis)mixed forest in Changbai Mountains,Northeast China.Chinese Journal of Applied Ecology,2010,21(9):2171-2178.
[11] Zhang X P,Wang X P,Zhu B,Zong Z J,Peng C H,Fang J Y.Litter fall production in relation to environmental factors in northeast China's forests.Journal of Plant Ecology.2008,32(5):1031-1040.
[12] Liu C Z,Li JW,Pan G L,Li CR.Litter production and dynamics in broadleaved-Korean pine forests in Xiaoxing'an Mountains.Chinese Journal of Ecology,1993,12(6),29-33
[13] Zhang D L,Mao Z J,Zhu S Y,Zhou B.Litter Falls of 6 major forest stands in Maoershan Mountain of Heilongjiang Province[J].Bulletin of Botanical Research,2008,28(1):104-108.
[14] Zhou G,Guan L,Zhang D,Zhang Q,Yan J,Wen D,Liu J,Huang Z,Kong G,Mo J,Yu Q.Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong,China.Plant Ecology,2007,188:77-89.
[15] Qingkui Wang,Silong Wang,Yu Huang.Comparisons of litterfall,litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China.Forest Ecology and Management,2008,255:1210-1218.
[16] Clark D A,Brown S,Kicklighter D W,Chambers J Q,Thomlinson J R,Ni J,Holland E A.Net primary production in tropical forests:an evaluation and synthesis of existing field data.Ecological Applications,2001,11(2):371-384.
[17] Piao SL,Fang JY,Guo Q H.Terrestrial net primary production and its spatio-temporal patterns in China during 1982-1999.Acta Scientiarum Naturalium Universitatis Pekinensis,2001,37(4):563-568.
[18] Chen JL,Jin G Z,Zhao F X.Litter decomposition and nutrient dynamics at different succession stages of typical mixed broadleaved-Korean pine forest in Xiaoxing'an Mountains.Chinese Journal of Applied Ecology,2010,21(9):2209-2216.
[19] Zhang D Q,Ye W H,Yu Q F,Kong G H,Zhang Y C.The litterfall of representative forests of successional series in Dinghushan,Acta Ecologica Sinica,2000,20(6):938-944.
[20] Zheng J P,Guo Z L,Xu C Y,Fan C N.Seasonal dynamics of litter accumulation in major forest communities on the northern slope of Changbai Mountain Northeast China.Acta Ecologica Sinica,2011,31(15):4299-4307.
[21] Ning X B,Xiang W H,Wang G J,Fang X,Yan W D,Deng X W.Litterfall production and dynamic for twenty years of a successive replanting Cunninghamia lanceolata plantation at Huitong,Hunan.Acta Ecologica Sinica,2009,29(9):5122-5129.
[22] Bray J R,Gorham E.Litter production in forests of the world.Advances in Ecological Research,1964,2:101-157.
[23] Deborah L.Regional-scale variation in litter production and seasonality in tropical dry forests of southern Mexico.Biotropica,2005,37(4):561-570.
[24] Binkley D,Dunkin KA,Bell D,Ryan M G.Production and nutrient cycling in mixed plantations of Eucalyptus and Albizia in Hawaii.Forest Science,1992,38(2),393-408.
[25] Wang J L,Tao Y,Lu ZW.Study on the characteristic of litterfall of Picea likiangensis var.linzhiensis forest in Tibet.Acta Phytoecologica Sinica,1998,22(6):566-570.
[26] Wang F Y.Review on the study of the forest litter fall.Advance in Ecology,1989,6(2):82-89.
[27] Liu Y,Han SJ,Lin L.Dynamic characteristics of litter falls in four forest types of Changbai Mountains,China.Chinese Journal of Ecology,2009,28(1):7-11.
[28] Wei L M,Yu D L,Chen Z R.Study on the dynamic variation of litter falls in Maolan Karst forest.Journal of Nanjing Forestry University(Natural Science Edition),2009,33(3):32-34.
[29] Liu SX,Li W P.Botany.Beijing:Science Press,2007.
参考文献:
[1] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡.植物生态学报,2000,24(5):518-522.
[2] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究.应用生态学报,2001,12(1):13-16.
[3] 刘国华,傅伯杰,方精云.中国森林碳动态及其对全球碳平衡的贡献.生态学报,2000,20(5):733-740.
[4] 汪思龙.森林残落物生态学.北京:科学出版社,2010:14-15.
[5] 彭少麟,刘强.森林凋落物动态及其对全球变暖的响应.生态学报,2002,22(9):1534-1544.
[6] 郭伟,张健,黄玉梅,刘旭,王伟,薛林.森林凋落物生态功能研究进展.安徽农业科学.2009,37(5):1984-1987.
[7] 潘开文,何静,吴宁,吴钦孝.森林凋落物对林地微生境的影响.应用生态学报,2004,15(1):153-158.
[8] 朱金兆,刘建军,朱清科.森林凋落物层水文生态功能研究.北京林业大学学报.2002,24(5/6):30-34.
[9] 李雪峰,韩士杰,李玉文,侯炳柱,李雪莲.东北地区主要森林生态系统凋落量的比较.应用生态学报,2005,16(5):783-788.
[10] 原作强,李步杭,蔺菲,师帅,叶吉,王绪高,郝占庆.长白山阔叶红松林凋落物组成及其季节动态.应用生态学报,2010,21(9):2171-2178.
[11] 张新平,王襄平,朱彪,宗占江,彭长辉,方精云.我国东北主要森林类型的凋落物产量及其影响因素.植物生态学报,2008,32(5):1031-1040.
[12] 刘传照,李景文,潘桂兰,李传荣.小兴安岭阔叶红松林凋落物产量及动态的研究.生态学杂志,1993,12(6):29-33.
[13] 张东来,毛子军,朱胜英,周彪.黑龙江省帽儿山林区6种主要林分类型凋落物研究.植物研究,2008,28(1):104-108.
[17] 朴世龙,方精云,郭庆华.1982—1999年我国植被净第一性生产力及其时空变化.北京大学学报:自然科学版,2001,37(4):564-568.
[18] 陈金玲,金光泽,赵凤霞.小兴安岭典型阔叶红松林不同演替阶段凋落物分解及养分变化.应用生态学报,2010,21(9):2209-2216.
[19] 张德强,叶万辉,余清发,孔国辉,张佑倡.鼎湖山演替系列中代表性森林凋落物研究.生态学报,2000,20(6):938-944.
[20] 郑金萍,郭忠玲,徐程扬,范春楠.长白山北坡主要森林群落凋落物现存量月动态.生态学报,2011,31(15):4299-4307.
[21] 宁晓波,项文化,王光军,方晰,闫文德,邓湘雯.湖南会同连作杉木林凋落物量20年动态特征.生态学报,2009,29(9):5122-5129.
[25] 王建林,陶渊,吕振武.西藏林芝云杉林凋落物的特征研究.植物生态学报,1998,22(6):566-570.
[36] 王凤友.森林凋落量研究综述.生态学进展,1989,6(2):82-89.
[27] 刘颖,韩士杰,林鹿.长白山四种森林类型凋落物动态特征.生态学杂志,2009,28(1):7-11.
[28] 魏鲁明,余登利,陈正仁.茂兰喀斯特森林凋落物量的动态研究.南京林业大报(自然科学版),2009,33(3):32-34.
[29] 刘胜祥,黎维平.植物学.北京:科学出版社,2007.
猜你喜欢
小哥白尼(野生动物)(2021年9期)2022-01-17
矿产勘查(2020年4期)2020-12-28
中国林副特产(2020年5期)2020-10-09
森林工程(2018年3期)2018-06-26
现代农业研究(2017年11期)2018-01-12
学校教育研究(2017年28期)2017-10-21
防护林科技(2017年7期)2017-09-01
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
卫星电视与宽带多媒体(2017年23期)2017-06-20
