
凝血因子ⅩⅢ基因多态性与肺血栓栓塞症的相关性研究
2013-10-25 09:36许晓岩杨媛华翟振国庞宝森
首都医科大学学报 2013年1期
许晓岩 杨媛华 翟振国 庞宝森 王 辰
(1.首都医科大学附属北京朝阳医院北京呼吸疾病研究所,北京100020;3.卫生部北京医院北京呼吸疾病研究所,北京100730)
肺血栓栓塞症(pulmonary thromboembolism,PTE)和下肢深静脉血栓形成(deep venous thrombosis,DVT)是静脉血栓症(venous thromboembolism,VTE)的2个不同阶段。目前认为VTE是由于遗传和(或)环境异常造成的一种多基因、多因素疾病。其中凝血系统基因多态性是研究方向之一。凝血因子ⅩⅢ,简称因子ⅩⅢ(FⅩⅢ)基因单核苷酸多态性(single nucleotide polymorphism,SNP)与疾病关系的研究[1-5]显示,A 亚基 Val34Leu(100G/T)多态性中突变等位基因T可能对心脑血管疾病和VTE具有保护作用,但我国对此的研究资料有限。关于B亚基His95Arg(8259A/G)多态性的资料不多,有研究[6]显示其突变等位基因 G可能与 A亚基Val34Leu多态性中的T等位基因有协同作用,显著降低心肌梗死的危险,但据报道[7],此突变会增加DVT的风险,未见其与PTE关系的报道,中国人中更未见该多态性的研究。本研究利用聚合酶链式反应(PCR)-限制性片段长度多态性(restricted fragment length polymorphisms,RFLP)技术,在中国汉族人群中对FⅩⅢ A亚基 Val34Leu和 B亚基His95Arg多态性与PTE的关系进行探讨。
1 资料与方法
1.1 研究对象
采用病例-对照研究。病例组来自2002年6月到2007年2月在首都医科大学附属北京朝阳医院呼吸科确诊的PTE患者114例。所有病例均按照中华医学会呼吸病学分会制定的《肺血栓栓塞症的诊断与治疗指南(草案)》[8]的标准,经核素肺通气/灌注扫描(V/Q)高度可能和/或螺旋CT肺动脉造影(computed tomography pulmonary angiography,CTPA)见血栓直接征象确诊PTE。对照组为与PTE患者1∶1配对的114例健康对照。对照组与PTE患者同为汉族,性别相同,年龄近似(±2岁),既往无DVT、PTE病史,与PTE患者无血缘关系,无心、脑、肺、肾、血液、内分泌系统疾病。主要来自门诊体检者和健康志愿者。对研究对象遵守医学伦理学的一般原则。
1.2 实验方法
1)标本采集:采空腹静脉血4 mL,EDTA抗凝。
2)DNA制备:采用低渗溶血法从新鲜全血中分离白细胞。采用酚-氯仿-异戊醇法提取DNA,用灭菌纯水调整DNA浓度在0.1 μg/μL左右,-20℃保存备用。
3)FⅩⅢ A亚基 Val34Leu基因多态性:采用PCR-RFLP联合聚丙烯胺凝胶电泳(PAGE)检测FⅩⅢ A亚基Val34Leu多态性。
参照文献[9]的引物。引物1:5'-CATGCCTTTTCTGTTGTCTTC-3';引物 2:5'-TACCTTGCAGGTTGACGCCCCGGGGCACTA-3',其中引物2将3'端第二个碱基由胞嘧啶(C)换成了胸腺嘧啶(T),从而引入了一个限制性内切酶DdeⅠ的酶切位点。在基因文库(GenBank)中进行核对并寻找相应序列。在eppendorf梯度核酸扩增仪进行热循环。25 μL PCR反应体系:样品DNA(0.1 μg/ μL)1.0 μL,dNTP 混合物(各 2.5 mmol/L)2.0 μL,引物(50 mmol/L)各 0.2 μL,Ex Taq DNA 聚合酶(大连宝生物工程公司,5U/ μL)0.2 μL,10×反应缓冲液(含Mg2+)2.5 μL,灭菌双蒸水 18.9 μL。PCR 反应条件:预变性95℃ 2 min;变性95℃ 30 s,复性58℃45 s,延伸72℃ 30 s,循环35次;后延伸:72℃ 5 min。2%琼脂糖凝胶电泳分析PCR产物。
取 PCR 扩增产物 8 μL,限制性内切酶 DdeⅠ1 μL(纽英伦生物技术有限公司,10U),缓冲液1 μL,制成混合物,37℃水浴4 h。12%PAGE,分析酶切产物。
4)FⅩⅢ B亚基 His95Arg基因多态性:采用PCR-RFLP联合琼脂糖凝胶电泳检测FⅩⅢ B亚基His95Arg多态性。
参照Reiner等[6]设计的引物。引物1:5'-AAAGACAAGCTTAGTTTCATCATT-3';引物2:5'-TCTTCAGTTTAGGAAATGATTCTTAT-3'。在基因文库(GenBank)中进行核对并寻找相应序列。在eppendorf梯度核酸扩增仪进行热循环。25 μL PCR反应体系:样品DNA(0.1 μg/ μL)1.0 μL,dNTP 混合物(各 2.5 mmol/L)2.0 μL,引物(50 mmol/L)各 0.2 μL,Ex Taq DNA 聚合酶(大连宝生物工程公司,5U/ μL)0.2 μL,10×反应缓冲液(含Mg2+)2.5 μL,灭菌双蒸水 18.9 μL。PCR 反应条件:预变性95℃ 2 min;变性95℃ 30 s,复性58℃30 s,延伸72℃ 30 s,循环35次;后延伸:72℃ 5 min。2%琼脂糖凝胶电泳分析PCR产物。
取 PCR 扩增产物 8 μL,限制性内切酶 NsiⅠ1 μL(纽英伦生物技术有限公司,10U),缓冲液1 μL,制成混合物,37℃水浴4 h。2% 琼脂糖凝胶电泳分析酶切产物。
1.3 统计学方法
采用SPSS 11.5统计软件建立数据库并进行数据整理与分析。计量资料以均数±标准差(±s)表示,计数资料以构成比或率表示。基因频率采用基因计数法计算,基因型分布是否符合Hardy-Weinberg遗传平衡定律及两组间等位基因频率的差异比较采用χ2检验;两组间计算比数比(OR)和95%的可信区间(CI)。以P<0.05为差异有统计学意义。
2 结果
2.1 PTE组和对照组的基本情况
PTE组和对照组各114例,其中每组男性70例,女性44例。2组平均年龄均为55岁(表1)。

表1 PTE组和对照组基本情况比较Tab.1 The age and sex of PTE patients and 114 healthy controls
2.2 FⅩⅢ基因多态性的PCR产物及酶切结果分析
1)FⅩⅢA亚基Val34Leu多态性:PCR扩增出1条含FⅩⅢ A亚基Val34Leu多态性位点的DNA片段,长192 bp(图1)。经限制性内切酶DdeⅠ酶切后,野生型(GG)没有酶切位点,不被切割,仍为192bp 1个片段;杂合型(GT)呈192bp、161bp和31bp 3个片段;未见突变型(TT)。由于31bp片段短,不易观察,故主要区分192bp和161bp 2个片段(图2)。
2)FⅩⅢB亚基His95Arg多态性:PCR扩增出1条含FⅩⅢB亚基His95Arg多态性位点的DNA片段,长264bp(图3)。经限制性内切酶NsiⅠ酶切后,野生型(AA)呈129bp和135bp 2个片段;杂合型(AG)呈264bp、129bp和135bp 3个片段;未见突变型(GG)。由于129 bp和135bp 2个片段仅相差6个bp,2%的琼脂糖凝胶电泳不能区分,仅呈1条条带(图4)。
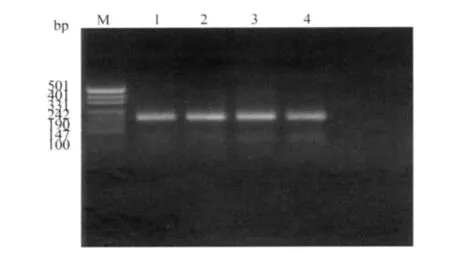
图1 PCR扩增出一条含FⅩⅢA亚基Val34Leu多态性位点的DNA片段,长192 bpFig.1 The polymerase chain reaction(PCR)was used to amplify a 192-bp fragment from the factorⅩⅢA subunit gene


图4 经限制性内切酶NsiⅠ酶切后Fig.4 The PCR product was digested with Nsi I
2.3 PTE组和对照组间FⅩⅢ基因多态性比较
1)FⅩⅢA亚基Val34Leu多态性:PTE组和对照组均未检测到突变纯合子(TT),并且 GG和 GT基因型在2组中分布相同,GG型113(99.1%),GT型1(0.9%),等位基因G和T的频率分别0.996,0.004(表2)。基因型分布均符合Hardy-Weinberg平衡。
2)FⅩⅢ B亚基His95Arg多态性:PTE组,FⅩⅢB亚基His95Arg多态性AA、AG、GG基因型的分布频数分别为111(97.4%)、3(2.6%)、0(0);等位基因A和G的频率分别0.987,0.013。对照组 AA、AG、GG基因型的分布频数分别为104(91.2%)、10(8.8%)、0(0)。等位基因A和G的频率分别0.956,0.044。对2组不同基因型实际频数和期望频数分别做χ2检验,差异无统计学意义,基因型分布均符合 Hardy-Weinberg平衡。2组均未检测到突变纯合子(GG)。2组间基因型的分布差异有统计学意义(χ2=3.997,P=0.046)。2组间等位基因的分布差异有统计学意义(χ2=3.880,P=0.049),PTE 组 G 等位基因分布显著低于对照组(表3)。相对于AA基因型而言,AG基因型有减低 PTE发生危险的趋势(OR=0.281,P=0.046)。

表2 FⅩⅢA亚基Val34Leu多态性基因型频率和等位基因频率Tab.2 FⅩⅢ A Val34Leu(100G/T)polymorphisms n(%)

表3 FⅩⅢB亚基His95Arg多态性基因型频率和等位基因频率Tab.3 FⅩⅢ B His95Arg(8259A/G)polymorphisms n(%)
3 讨论
凝血因子ⅩⅢ,简称因子ⅩⅢ(FⅩⅢ)是机体凝血瀑布反应的最后一个酶,又称纤维蛋白稳定因子。血浆中的FⅩⅢ 以酶原形式存在,呈四聚体结构(A2B2),即2个A亚基和2个B亚基组成,其中A亚基具有转谷氨酸酶活性,B亚基起载体作用。血浆中的FⅩⅢ活化为FⅩⅢa需要2步,第1步为凝血酶裂解A亚基精氨酸(Arg)37-甘氨酸(Gly)38键;第2步为B亚基从A亚基分离,使A亚基的半胱氨酸活性位点暴露。
FⅩⅢ A亚基 Val34Leu(100G/T)多态性与FⅩⅢB亚基His95Arg(8259A/G)多态性位于编码区,造成了氨基酸的置换,可能影响了FⅩⅢ 的活化,进一步影响纤维蛋白凝块的结构及稳定性[10]。
FⅩⅢ A亚基 Val34Leu(100G/T)多态性是指F13A基因的2号外显子发生G→T的点突变,使34号密码子所编码的氨基酸由缬氨酸变为亮氨酸(Val34Leu)。这一多态性位点距凝血酶对FⅩⅢ 的活化部位仅3个氨基酸。现有的研究发现这一突变不影响FⅩⅢ的血浆水平,但使凝血酶对FⅩⅢ 的活化速度,在34Leu明显快于野生型34Val,使纤维蛋白较快发生交联,从而使血小板黏附下降,终止凝血过程,导致纤维蛋白凝块结构改变,稳定性下降[11-15]。关于FⅩⅢA亚基Val34Leu多态性与VTE的关系国外有多篇报道。但对于T等位基因是否对VTE有保护作用,所得出的结论不一。2项Meta分析研究[4-5]中关于该多态性位点与VTE关系的研究,认为T等位基因对VTE起保护作用。另外,文献[16]报道FⅩⅢA亚基Val34Leu多态性存在明显的种族差异。这一多态性在美洲印第安人最高(51.2%),西方白色人种次之(44.3%),黑色人种也较常见(28.9%),而在日本人中最低(2.5%)。
我们的研究仅在PTE组和对照组中各检测到1例FⅩⅢA亚基Val34Leu突变杂合子,即GT基因型,2组中基因型分布及等位基因频率相同。T等位基因在中国汉族人群中少见,基因频率为0.004,与以往的研究[17]相符。说明FⅩⅢ A亚基Val34Leu多态性可能对中国汉族人群PTE的发生影响很小;也进一步体现了基因多态性的地区和种族差异。说明尽管生活环境、习惯及其他因素在疾病的发生过程中起着重要作用,但不同种族在遗传背景上的差异也是导致疾病易感性不同以及临床表现不尽相同的一个重要因素。
FⅩⅢB亚基His95Arg(8259A/G)多态性是指F13B基因8259位点A→G突变,该突变位于3号外显子上,导致FⅩⅢB亚基95位组氨酸(His)被精氨酸(Arg)替代。3号外显子编码FⅩⅢ B亚基的第2个Sushi域,该域参与 A、B亚基的结合。Komanasin等[7]在 Leeds(214 例患者-116 例 DVT、98 例 PTE,247例对照)和LETS(471例DVT,472例对照)两项病例对照研究中探讨FⅩⅢ B亚基His95Arg多态性与VTE的关系,发现突变杂合子AG增加VTE的危险,OR值(95%CI)分别为1.6(1.0~2.6)和1.4(1.0~2.0)。Reiner等[6]一项关于女性心肌梗死(MI)发生危险性的研究,未发现FⅩⅢ B亚基His95Arg多态性与MI相关,但G等位基因与FⅩⅢA亚基Val34Leu多态性中的T等位基因有协同作用,在T等位基因携带者中,G等位基因显著降低MI的危险,并能降低雌激素替代治疗者发生MI的危险。
本研究首次在中国汉族人群中探讨FⅩⅢB亚基His95Arg多态性,PTE组和对照组均未发现突变纯合子(GG)。对照组 AA、AG基因型分别为 104(91.2%)、10(8.8%),PTE 组 AA、AG 基因型分别为111(97.4%)、3(2.6%),AG基因型在PTE组检出较少(P=0.046),相对于AA基因型而言,AG基因型有减低PTE发生危险的趋势(OR=0.281),但95%CI在0.075~1.050,进一步增大样本量,有望得到阳性结果。这与Komanasin等[7]得出的结论不符,考虑可能与疾病类型及人种、地区不同有关。对于FⅩⅢ B亚基His95Arg多态性的研究资料有限,有待更多的研究阐述其与血栓性疾病的关系。
中国汉族人群中,FⅩⅢ A亚基Val34Leu(100C/T)多态性少见,该多态性可能对中国汉族人群PTE的发生影响很小。中国人群中存在FⅩⅢ B亚基His95Arg(8259A/G)多态性。健康对照组中AA、AG、GG 3种基因型的分布为104(91.2%)、10(8.8%)、0(0),G等位基因的频率为0.044。基因型分布均符合Hardy-Weinberg平衡。突变杂合子AG基因型有减低PTE发生危险的趋势。
[1]Voetsch B,Loscalzo J.Genentic determinants of artierial thrombosis[J].Thromb Vasc Biol,2004,24:226-229.
[2]Voko Z,Bereczky Z,Katona E,et al.FactorⅩⅢ Val34Leu variant protects against coronary artery disease:A metaanalysis[J].Thromb Haemost,2007,97(3):458-463.
[3]Shafey M,Anderson J L,Scarvelis D,et al.FactorⅩⅢVal34Leu variant and the risk of myocardial infarction-A metaanalysis[J].Thromb Haemost,2007,97(4):635-641.
[4]Wells P S,Anderson J L,Scarvelis D K,et al.FactorⅩⅢVal34Leu variant is protective against venous thromboembolism:a HuGE review and Meta-Analysis[J].Am J Epidemiol,2006,164(2):101-109.
[5]Gohil R,Peck G,Sharma P.The genetics of venous thromboembolism.A meta-analysis involving approximately 120,000 cases and 180,000 controls[J].Thromb Haemost,2009,102(2):360-370.
[6]Reiner A P,Heckbert S R,Vos H L,et al.Genetic variants of coagulation factorⅩⅢ,postmenopausal estrogen therapy,and risk of nonfatal myocardial infarction[J].Blood,2003,102(1):25-30.
[7]Komanasin N,Catto A J,Futers T S,et al.A novel polymorphism in the factorⅩⅢ B-subunit(His95Arg):relationship to subunit dissociation and venous thrombosis[J].J Thromb haemost,2005,3(11):2487-2496.
[8]中华医学会呼吸病学分会.肺血栓栓塞症的诊断与治疗指南(草案)[J].中华结核和呼吸杂志,2001,24:259-264.
[9]Kangsadalampai S, Board P G. The Val34Leu polymorphism in the a subunit of coagulation factorⅩⅢcontributes to the large normal range in activity and demonstrates that the activation peptide plays a role in catalytic activity[J].Blood,1998,92(8):2766-2770.
[10]ScottE M, Ariёns R, GrantP J. Genetic and Environmental Determinants of Fibrin Structure and Function:Relevance to Clinical Disease[J].Arterioscler Thromb Vasc Biol,2004,24(19):1558-1566.
[11]Balogh I,Szoke G,Karpati L,et al.Val34Leu polymorphism of plasma factor Ⅹ Ⅲ:biochemistry and epidemiology in familial thrombophilia[J].Blood,2000,96(17):2479-2486.
[12]Ariens R A,Philippou H,Nagaswami C,et al.The factorⅩⅢV34L polymorphism accelerates thrombin activation of factorⅩⅢ and affects cross-linked fibrin structure[J].Blood,2000,96(3):988-995.
[13]Ariens R A,Lai T S,Weisel J W,et al.Role of factorⅩⅢin fibrin clot formation and effects of genetic polymorphisms[J].Blood,2002,100(3):743-754.
[14]Shemirani A H,Haramura G,Bagoly Z,et al.The combined effect of fibrin formation and factorⅩⅢ A subunit Val34Leu polymorphism on the activation of factorⅩⅢ in whole plasma[J].Biochim Biophys Acta,2006,1764(8):1420-1423.
[15]Bagoly Z,Koncz Z,Hársfalvi J,et al.FactorⅩⅢ,clot structure,thrombosis[J].Thromb Res,2012,129(3):382-387.
[16]Attié-Castro FA,Zago M A,Lavinha J,et al.Ethnic heterogeneity of the factorⅩⅢ Val34Leu polymorphism[J].Thromb Haemost,2000,84(4):601-603.
[17]郑红,贺颖,连建华,等.深静脉血栓形成患者凝血因子III Val34Leu基因多态性检测[J].郑州大学学报:医学版,2006,41(2):280-282.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
智慧健康(2021年17期)2021-07-30
国际检验医学杂志(2021年7期)2021-04-15
世界最新医学信息文摘(2020年68期)2020-12-25
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
延安大学学报(医学科学版)(2019年1期)2019-03-29
现代检验医学杂志(2016年5期)2016-08-20
西南农业学报(2016年6期)2016-04-16
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
河南医学研究(2014年7期)2014-02-27
