
大长山岛虾夷扇贝养殖海区及其体内细菌数量周年变化
2015-10-09 03:11赵晓美张公亮侯红漫
大连工业大学学报 2015年2期
赵晓美,桑 雪,张公亮,侯红漫
(大连工业大学食品学院,辽宁大连 116034)
大长山岛虾夷扇贝养殖海区及其体内细菌数量周年变化
赵晓美,桑雪,张公亮,侯红漫
(大连工业大学食品学院,辽宁大连116034)
采用平板涂布法(PC法)对大连长海县大长山岛虾夷扇贝(Mizuhopectenyessoensis)养殖海区的表层海水、10m层海水、成年虾夷扇贝鳃中的异养菌和致病性弧菌数量的周年变化进行了调查研究。结果表明,表层海水、10m层海水和虾夷扇贝鳃中,异养菌和致病性弧菌数量最高值均出现于5~7月,异养菌数量最高值分别为7.9×105、5.9×105cfu/mL和1.2×105cfu/g,致病性弧菌数量最高值分别为1.6×102、1.7×102cfu/mL和87cfu/g;最低值均出现于11月至次年1月,异养菌数量最低值分别为1.7×105、8.0×104cfu/mL和2.7×104cfu/g,致病性弧菌数量最低值分别为60、67cfu/mL和28cfu/g。虾夷扇贝的鳃中致病性弧菌数量于春季呈现最高值87cfu/g。
虾夷扇贝;异养菌;致病性弧菌;养殖海域
0 引 言
虾夷扇贝(Mizuhopectenyessoensis)主产于日本北部及俄罗斯远东地区沿海水域,属于冷水性双壳贝类[1]。20世纪80年代初引入中国后,迅速开展增养殖研究和实践。该扇贝生长快、肉味鲜美、营养价值及经济价值高,深受广大养殖户和消费者的欢迎,现已成为中国北方重要的经济养殖贝类之一。虾夷扇贝的养殖目前主要集中在辽东半岛和山东长岛等海区,其中以大连长海县养殖规模最大。
目前对其研究主要集中在增养殖技术[2-3]、家系选育技术[4]、遗传多样性分析[5]、食用深加工[6]、副产品高值化利用[7-8]等方面,关于虾夷扇贝养殖海域中细菌数量及其变化的研究报道较少。水中细菌是养殖生态环境中的重要组成部分,与养殖用的水质和生物疾病的发生有着密切的关系,养殖水体中的营养物质丰富,细菌的数量大大高于正常生态环境,而大部分异养细菌依然是分解者,消耗水体中的富营养物质,维持物质循环;但部分细菌的大量繁殖会威胁到养殖生物的健康,从而引起生物体的疾病。对养殖生物而言,环境细菌生态平衡出现异常往往会引起细菌性疾病的暴发,环境一旦失衡,病原菌大量繁殖,加之养殖生物体免疫力下降,就可引发疾病[9-11],因而养殖环境和养殖生物体内细菌的数量、种群变动对于细菌性疾病预警有着重要的意义。虾夷扇贝为滤食性动物,鳃作为滤食的主要器官,其内部细菌数量与养殖水域中细菌数量密切相关,因此检测虾夷扇贝鳃中的细菌数量对评价其生存海域的细菌环境和其在养殖过程中的患病情况也具有一定意义。本研究中,作者对虾夷扇贝养殖海域的海水以及虾夷扇贝鳃中异养菌及致病性弧菌数量的周年变化进行了研究,旨在为评价该海域水质的营养状况和环境条件以及防治扇贝养殖过程中由细菌引起的病害提供基础的科学依据。
1 材料与方法
1.1样品采集
于2012年8月~2013年7月在长海县大长山岛扇贝养殖区(122°32′55.50″~122°34′13.26″E,39°15′10.74″N~39°15′34.02″)采集水样及虾夷扇贝,使用经过灭菌的洁净纯净水瓶盛装水样,并在采样前用所采水样将采样瓶反复冲洗,分别采集表层、10m层水样;虾夷扇贝样品均为成年扇贝,贝壳直径为75mm左右。每2个月采集一次。样品采集完成后放入4℃恒温箱保存,并迅速运回实验室进行后续实验。
1.2培养基的准备
异养细菌的培养使用2216E海水培养基,主要成分为蛋白胨5.0g,酵母膏1.0g,磷酸高铁0.10g,琼脂20.0g,陈海水1000mL。121℃灭菌20min后倒制平板。
致病性弧菌的培养使用弧菌显色培养基TCBS(硫代硫酸钠枸橼酸胆盐蔗糖琼脂),主要成分为酵母粉5.0g,蛋白胨10.0g,硫代硫酸钠10.0g,枸橼酸钠10.0g,牛胆粉5.0g,胆酸钠3.0g,蔗糖20.0g,氯化钠10.0g,柠檬酸铁1.0g,百里酚兰0.04g,琼脂15.0g,pH(8.6±0.1)。使用时称取89g,煮沸溶解于1000mL蒸馏水中,冷至60℃时,倾入无菌平皿,无须高压灭菌。
1.3细菌的培养及菌落计数
在无菌条件下,使用75%无菌乙醇擦拭扇贝样品体表并用无菌海水冲洗,将虾夷扇贝鳃在无菌超净台上切下转移至无菌离心管中,加入无菌PBS缓冲溶液(0.1mol/L,pH7.4)漩涡振荡5min,然后在4℃下700g离心5min,取上清液作为菌液进行后续实验。
按照《海洋调查规范》[12]中的平板涂布法(PC法)进行细菌培养及菌落计数,取所采水样及上清液各1mL,用灭菌海水对其进行10倍梯度稀释,得到不同浓度梯度的菌液后各取100μL涂布平板,每个浓度梯度的菌液涂布3个平板,然后在恒温培养箱中于(29±1)℃下培养2~5d,最后进行平板菌落计数。
1.4结果统计与分析
2216E培养基生成白色菌落,TCBS培养基生成黄色或绿色菌落,对各个平板生成的菌落进行计数,取菌落数30~300cfu/mL或30~300cfu/g为有效菌落数(海水样品中细菌的计数单位为cfu/mL,虾夷扇贝鳃中细菌的计数单位为cfu/g),并计算3次实验的平均值,得出异养菌和致病性弧菌数量的周年变化。
2 结果与讨论
2.1菌落计数结果
菌落计数结果如表1、2所示。表层海水、10m层海水以及虾夷扇贝鳃中的异养菌总数,1月最少,分别为1.7×105、8.0×104cfu/mL和2.7×104cfu/g;而7月均达到各自最大值7.9× 105、5.9×105cfu/mL和1.2×105cfu/g。表层海水中弧菌数量最低为60cfu/mL,出现在11月,最高达到160cfu/mL,出现于7月;10m层海水中弧菌数量为67~170cfu/mL,1月最低,7月最高;虾夷扇贝鳃中弧菌数量为28~87cfu/g,11月最低,5月最高。

表1 海水中细菌数量Tab.1 Number of bacteria in the seawater

表2 虾夷扇贝体内细菌数量Tab.2 Number of bacteria in scallop
2.2异养菌数量周年变化
异养细菌分解有机物是海洋生态系中能量流动和物质循环中诸多环节中的重要环节之一,异养菌的丰度、活性、水体环境有机物的浓度三者关系密切,因而可以作为海域受有机污染物污染程度指标。此外,海洋中进入溶解状态的有机物质主要靠海洋异养细菌来转化,因此异养细菌在海洋生物生产和生源要素循环中起着十分重要的作用,是微食物环的核心部分。
2.2.1海水中异养菌数量变化
两组海水样品中异养菌数量的周年变化规律如图1所示,从月份看,菌体数量由高到低顺序为7月、5月、3月、9月、11月、1月;从季节上看,由高到低顺序为夏季、春季、秋季、冬季,均为夏季最高,冬季最少,春、秋两季介于二者之间,这与细菌分布的一般规律相符,并且异养菌数处于同一数量级。从空间上来看,一年中各实验月,表层海水中异养菌数量大于10m层海水中异养菌数量,这可能是由于可供异养菌生存的水体有机物以及海洋污染物多存在于海水表层。

图1 海水中异养菌数量的周年变化Fig.1 Annual changes of heterotrophic bacteria numbers in the seawater
2.2.2虾夷扇贝体内异养菌数量
如图2所示,虾夷扇贝鳃中异养菌数量的周年变化趋势,从月份看依次为7月、5月、3月、9月、11月、1月;从季节看依次为夏季、春季、秋季、冬季,夏季最高,冬季最少,春、秋两季介于二者之间。与海水中异养菌数量相比较少,是由于虾夷扇贝自身免疫系统的作用可以杀死部分异养菌。

图2 虾夷扇贝鳃中异养菌数量的周年变化Fig.2 Annual changes of heterotrophic bacteria numbers in the gills of scallop
2.3弧菌数量周年变化
弧菌是自然海水中生存的正常菌群成员之一,是目前海水养殖业中最为常见的条件致病菌,广泛存在于海水以及海产动物如鱼、虾和贝类的消化道和体表上,当环境条件适宜时引起其大量繁殖并表现致病性。一般情况下,自然海水中弧菌的数量较低,但在海水养殖环境中及排污口附近海区其数量明显较高。弧菌喜好生长于有机质丰富的环境中,其数量的多寡可以指示环境的污染程度[10]。
2.3.1海水中致病性弧菌数量
海水中致病性弧菌数量变化如图3所示。从月份看,菌体数量由高到低顺序为7月、5月、9月、3月、1月、11月,10m层海水中菌体数量由高到低顺序为7月、5月、9月、3月、11月、1月;从季节上来看,海水中为夏季最高,冬季最低,春秋两季介于二者之间。

图3 海水中致病性弧菌数量的周年变化Fig.3 Annual changes of Vibrio numbers in the seawater
2.3.2虾夷扇贝体内致病性弧菌数量
在虾夷扇贝鳃中的致病性弧菌数量变化如图4所示。从月份看,菌体数量由高到低顺序为5月、7月、3月、9月、1月、11月;从季节性来看,春夏秋三季均高于冬季,致病性弧菌春季数量的增大,可能是导致虾夷扇贝在春季育苗期间大量死亡的原因之一。因此,加强春季该水域弧菌数量的监测,并在其数量大幅度增加时,采取措施降低致病性弧菌数量,将会有效预防虾夷扇贝细菌性疾病的发生。
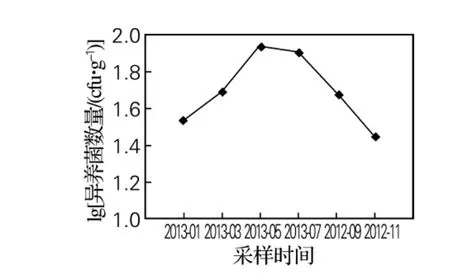
图4 虾夷扇贝鳃中致病性弧菌数量的周年变化Fig.4 Annual changes of Vibrio numbers in the gills of scallop
3 结 论
细菌在海洋中既是生产者,又是分解者,它们是水域中食物链和微生物环的重要组成部分,所以通常通过统计它们的细胞数量和种群的组成来评价水质的营养状况、水域的环境条件,当然水域环境条件的各种物理、化学和生物因素也会直接影响它们的生态分布,因此通过调查某一海域中细菌数量的变化和某些特殊生理类群的细菌的生态分布,可以为了解该海域的环境条件、生境的退化和修复提供基础资料[13]。异养菌数是最常用来指示有机质污染的一个指标,我国国家海洋局在2002年4月颁布的《海水增养殖区监测技术规程》中加入了异养菌指标,并以《异养细菌总数评价等级表》作为参考标准。
本实验通过调查2012年8月—2013年7月大长山岛扇贝养殖海区表层海水、10m层海水与该海区养殖的成年虾夷扇贝鳃中的异养菌、致病弧菌数量,分析了其周年变化趋势,为评价该养殖海域的海水质量、污染情况、水质营养状况等指标提供了基础依据,并为防治扇贝养殖中由于细菌引起的病害起到积极作用。
[1]NAGASHIMAK,SATOM,KAWAMATAK,etal.GeneticstructureofJapanesescalloppopulation inHokka-Ido,analyzedbymitochondrialhaplotype distribution[J].MarineBiotechnology,2005,7(1):1-10.
[2]李文姬,薛真福.持续发展虾夷扇贝的健康增养殖[J].水产科学,2005,24(9):49-51.
[3]张起信,张启胜,李维年,等.虾夷扇贝筏式养殖高产技术研究[J].海洋科学,2000,24(8):14-16.
[4]梁峻,郑怀平,李莉,等.虾夷扇贝养殖群体的遗传力估算[J].海洋科学,2011,35(3):1-7.
[5]常亚青,陈晓霞,丁君,等.虾夷扇贝5个群体的遗传多样性[J].生态学报,2007,27(3):1145-1152.
[6]闫雪,杨静峰,周大勇,等.虾夷扇贝内脏多糖SVP_12的分离纯化及性质研究[J].食品与发酵工业,2009,35(2):172-175.
[7]金文刚,吴海涛,朱蓓薇,等.虾夷扇贝生殖腺多肽的制备及分离[J].大连工业大学学报,2011,30(6):391-395.
(JINWen-gang,WUHai-tao,ZHUBei-wei,etal.PreparationofpeptidesfromdenaturedPatinopecten yessoensisgonadwithneutralproteinase[J].Journal ofDalianPolytechnicUniversity,2011,30(6):391-395.)
[8]曹倩倩,朱蓓薇,杨静峰,等.扇贝糖原酶法提取及其硫酸化修饰条件[J].大连工业大学学报,2012,31(1):15-18.
(CAOQian-qian,ZHUBei-wei,YANGJing-feng,etal.Extractionandsulfationofscallopmuscleglycogen[J].JournalofDalianPolytechnicUniversity,2012,31(1):15-18.)
[9]LIUPing-chung,LINJi-yang,CHUANGWenhsiao,etal.IsolationandcharacterizationofpathogenicVibrioharveyi(V.carchariae)fromthe farmedmarineCobiafishRachycentroncanadumL.withgastroenteritissyndrome[J].WorldMicrobiol Biotechnol,2004,20(1):495-499.
[10]LIUPing-chung,LINJi-yang,HSIAOPei-tze,etal.Isolationandcharacterizationofpathogenic VibrioalginolyticusfromdiseasedcobiaRachycentroncanadum[J].JournalofBasicMicrobiology,2004,44(1):23-28.
[11]王瑞旋,徐力文,冯娟.海水鱼类细菌性疾病病原及其检测、疫苗研究概况[J].南方水产,2005,1(6):72-79.
[12]国家技术监督局.海洋调查规范[S].北京:中国标准出版社,2007:18-30.
[13]徐怀恕,杨学宋,李药,等.对虾苗期细菌病害的诊断与控制[M].北京:海洋出版社,1999:34-40.
TheannualchangeofbacterianumbersinDachangshanIslandaquaculture farmanditscultivatedJapanesescallop(Mizuhopectenyessoensis)
ZHAOXiaomei,SANGXue,ZHANGGongliang,HOUHongman
(SchoolofFoodScienceandTechnology,DalianPolytechnicUniversity,Dalian116034,China)
TheannualchangeofheterotrophicbacteriaandpathogenicVibrionumbersinDachangshan Island,DalianChanghaiaquaculturefarmanditscultivatedJapanesescallop(Mizuhopecten yessoensis)wasstudiedbythedilutedplatemethod.whichwasobtainedfrom.Themaximum numbersofheterotrophicbacteriawere7.9×105,5.9×105cfu/mL,1.2×105cfu/gandVibriowere 1.6×102,1.7×102cfu/mL,87cfu/grespectivelyinthesurfaceseawater,10m-depthseawaterand adultscallops’gillsinMaytoJuly.Theminimumnumbersofheterotrophicbacteriawere1.7×105,8.0×104cfu/mL,2.7×104cfu/gandVibriowere60,67cfu/mL,28cfu/grespectivelyinNovember toJanuary.ThemaximumnumbersofVibrioinscallops’gillswere87cfu/ginspring.
Mizuhopectenyessoensis;heterotrophicbacteria;PathogenicVibrio;aquaculturefarm
TS254.1
A
1674-1404(2015)02-0084-04
2013-12-31.
辽宁省高等学校重大科技平台项目(2011191).
赵晓美(1989-),女,硕士研究生;通信作者:侯红漫(1964-),女,教授.
猜你喜欢
当代水产(2022年8期)2022-09-20
食品安全导刊(2021年20期)2021-08-30
家教世界·创新阅读(2019年11期)2019-12-10
家教世界(2019年31期)2019-12-05
当代水产(2019年5期)2019-07-25
人生与伴侣·共同关注(2018年8期)2018-02-16
中成药(2017年12期)2018-01-19
湖南畜牧兽医(2016年3期)2016-06-05
广东海洋大学学报(2015年3期)2015-12-22
川北医学院学报(2015年5期)2015-12-05
