
玉米杂交种及其亲本氮同位素组成差异研究
2015-12-05 02:57王周锋刘卫国郝瑞娟张岁岐
作物研究 2015年4期
王周锋,刘卫国 ,郝瑞娟,张岁岐
(1长安大学环境科学与工程学院/旱区地下水文与生态效应教育部重点实验室,陕西西安710054;2中国科学院地球环境研究所/黄土与第四纪地质国家重点实验室,陕西西安710075;3西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌712100)
杂交育种作为一种有效的育种手段,由于包含了杂种优势群的划分、自交系的培育、对自交系一般配合力和特殊配合力的检测等内容,执行起来不仅费时费力而且育种成本也很高[1]。而杂种优势作为普遍存在的一种复杂生物学现象,在农业生产中获得了广泛应用,但对其形成机理迄今尚未阐述清楚[2]。因此,检测和识别杂种优势效应,快速筛选出强优势组合,将大大提高杂交育种的效率。利用植物体氮同位素技术可以拓展科学家在植物吸收和利用氮素方面的机理机制认识[3]。植物体的生理特征例如氮素吸收机制、氮素同化路径的差异性以及氮素在植物体内的重新分配均会导致植物体氮同位素值的差异性。植物种间的氮同位素值差异可能是由于不同植物品种对不同形式氮源的依赖程度不同[4]。Robinson 等[5]对30 种不同野生大麦的研究发现,不同大麦品种氮同位素值差异达到1.5‰。生长在硝态氮介质中的植物,不同基因型间硝酸还原酶分布的差异导致吸收过程中的分馏以及植物根叶间氮同位素值的差异[3]。Warner等[6]对 Houdeum bulgare的研究显示,基因型间的差异是由于硝酸还原酶在体内分布不同导致,说明无论在哪种形式的氮素介质中,氮同位素的分馏是由于同化酶和外界氮源浓度共同作用的结果。可见,利用同位素技术能够获取植物种间氮素吸收和分配等信息。
玉米是世界上最主要的粮饲兼用的作物之一,在我国约有2/3的玉米种植在干旱半干旱地区,常年受旱面积为40%,减产幅度为30%[7]。杂种优势的利用是作物育种上提高作物产量的主要手段之一,玉米是世界上杂种优势利用面积最大的作物[8]。利用杂种优势理论育种也是提高玉米产量的主要手段[9]。氮素是植物生长的主要矿质元素,氮素的吸收以及在植物体内的分配不同决定了植物生长策略的不同[10],对于植物体内不同部位氮同位素值的研究可以指示植物体内氮素的分布状况。因此,本研究选定具有杂种优势的子代及其父母本,通过研究氮同位素值在根、茎、叶中的差异性,分析杂种优势和氮素分配之间的关系,为进一步深入研究氮素在植物体内的分配以及杂种优势提供科学依据。
1 材料与方法
1.1 试验材料
供试材料为黄土高原主栽品种户单4号(抗旱)及其父本803(不抗旱)和母本天四(抗旱)。材料由西北农林科技大学玉米研究所育种室提供。
玉米种子经0.1%的HgCl2溶液消毒10 min,取出用自来水反复冲洗,再用蒸馏水冲洗,在蒸馏水中吸胀6 h,然后放入蛭石与石英沙(V/V=2/3)混合培养介质中在25℃培养箱萌发,出苗约3 d后当种子根长至5~6 cm时,将苗移入高20 cm、直径18 cm的塑料桶中培养(苗基部用脱脂棉裹住,桶上部用塑料泡膜板作支架,桶外部用双层黑塑料布遮光。每桶留苗3株,每个品种设6个重复。起初在桶中装入蒸馏水,使植株适应生长24 h后换成营养液。营养液成分:1/2 Hoagland全营养液。
将塑料桶放入日产KG-206SHL-D型人工气候室中培养,白天光照为250~300 μmol photons/(m2·s),光暗周期为14/10 h,昼夜温度为27/20℃,空气相对湿度(RH)为60% ~70%。每天用加氧泵向溶液中通气3~4次,每次60 min,保证根系良好生长。每48 h换一次营养液。待幼苗长至20 d后测定(苗龄23 d,处理18 d),采集植物根、茎、叶,在40℃恒温鼓风烘箱中烘干至恒重,用玛瑙研钵研磨后进行样品测试。
1.2 研究方法
植物氮同位素值的测定:在中国科学院地球环境研究所同位素实验室进行。用Finngan公司生产的EA-Delta plus质谱仪测试样品的δ15N和氮含量,其中氮同位素值的计算公式为:
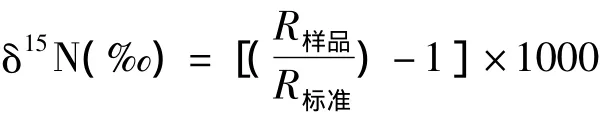
式中:R为15N/14N,氮的标准为大气氮,偏差为±0.3‰,含量的偏差为±0.1%。EA氧化炉温度设置为980℃,还原炉温度设置在650℃,样品测试时间设置为1 800 s。
2 结果与分析
2.1 杂交种及其亲本的氮同位素组成及氮含量差异
研究表明,杂交种及其亲本在根、茎、叶内的氮含量分布为叶片和茎中氮含量明显较根系高,叶片和茎氮含量分别为6.2%和6.3%,根系中氮含量为4.9%(图1)。对比杂交种及其亲本根、茎、叶中氮含量可以看出,叶片中氮含量大小顺序为子代户单4号>母本天四>父本803,在茎中氮含量大小依次为父本803>母本天四>户单4号,根系中没有明显的规律性,这说明父母本和子代玉米氮素在体内的分配不同。
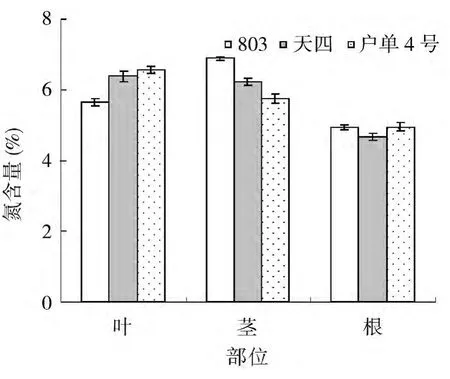
图1 杂交种及其亲本不同部位氮含量
杂交种及其亲本氮同位素组成研究表明:根、茎、叶中氮同位素组成的规律一致,茎中氮同位素组成最高,根系中氮同位素组成最低。在根、茎、叶中的氮同位素组成分别为6.4‰、11.8‰、10.0‰。对比杂交种及其亲本根、茎、叶中氮含量可以看出,在根、茎、叶中,均为母本天四的氮同位素组成最低。在茎和叶中,父本803氮同位素组成最高,而在根系中,子代户单4号氮同位素组成最高(图2)。
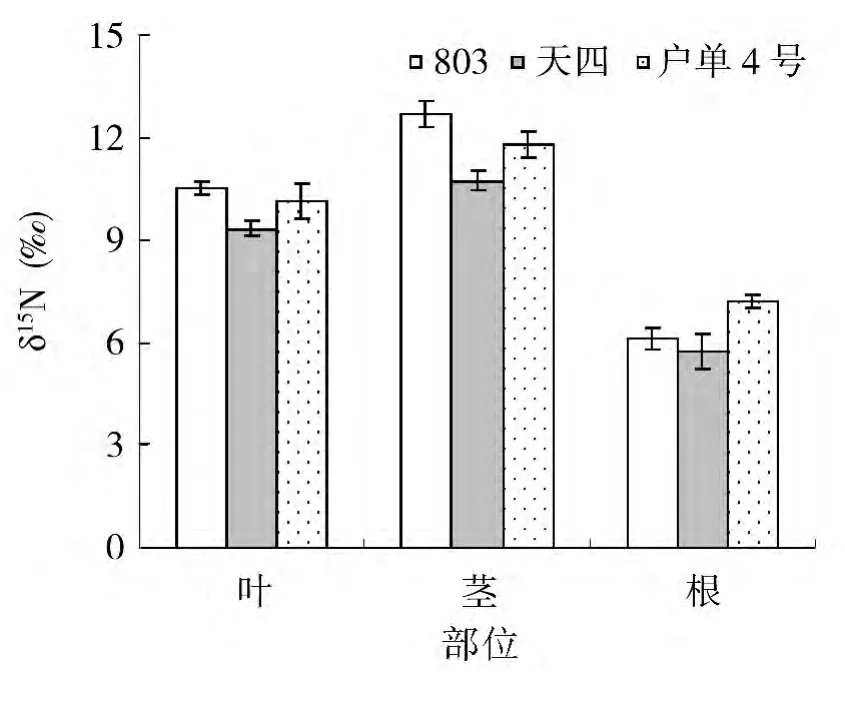
图2 杂交种及其亲本氮同位素组成在不同部位的组分
2.2 杂交种及其亲本的根系与茎、叶中氮同位素组成差异
根系是植物吸收矿质养分的主要通道,以往的研究表明,植物吸收的硝态氮可以在根系和叶片中同化,这会导致植物体内氮同位素分馏的发生。为了分析不同植物体内氮同位素分馏的差异性,本研究对比了根系与叶片、根系与茎中的氮同位素组成的差异(Δδ15N根-叶,Δδ15N(根-茎))。由图 3 可以看出,杂交种及其亲本Δδ15N根-叶差异较为明显,其中父本 803Δδ15N(根-叶)差异最大,达到 - 4.4‰,子代户单4号 Δδ15N(根-叶)的差异最小为 -3.0‰。结合根系和茎中的 Δδ15N(根-茎)的差异性可以看出(图4),其差异比 Δδ15N(根-叶)更大,父本803Δδ15N(根-茎)差异最大,达到-6.6‰,子代户单4号差异性最小,达到-4.6‰。但是无论是根系和叶片的差异还是根系和茎的差异,杂交种及其亲本均显示了一致的趋势,即杂交种及其亲本 Δδ15N(根-叶)和 Δδ15N(根-茎)均为户单4号<母本天四<父本803。

图3 杂交种及其亲本根系和茎中氮同位素组成差异
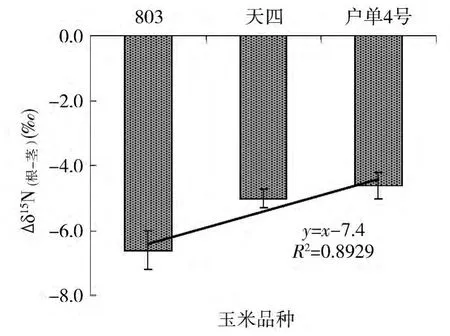
图4 杂交种及其亲本根系和叶中氮同位素组成差异
3 讨论
3.1 杂交种及其亲本氮素在体内的分配策略
当植物叶片和根系的氮源相同时,其氮同位素组成的差异就反应了植物体内氮素在传输和转化过程中的分馏效应[10]。本研究为室内水培试验,硝态氮是其生长的唯一氮源。结果表明植物根系、叶片和茎中的同位素组成为茎>叶片>根系,这说明在植物氮素的传输和转化过程中,确实存在氮同位素的分馏效应。Evans[3]研究认为,植物吸收的硝态氮首先在根系中同化,如果氮源充足,则会产生氮同位素的分馏作用,这主要是因为根系吸收的氮素一部分在根系中同化,由于15N和14N活性的差异,植物优先同化14N,那么剩余部分氮源的同位素组成偏正,导致植物叶片部分同化的氮素同位素组成偏正[3,11]。本研究采用的营养液为 1/2Hogland 营养液,其氮素浓度为7.5 mmol/L,相比以往的研究,浓度较高[3,12],从 Evans理论可以得出,植物叶片同位素组成应该大于根系氮同位素组成,本研究结果进一步验证了上述结论,说明在高浓度氮源的作用下,玉米体内氮素在根系和叶片中同时同化吸收。结合氮含量数据可以看出,玉米将更多的氮素利用在植物叶片上,这有利于植物高效的进行光合作用,同化更多的有机化合物。从玉米不同部位氮含量(图1)可以看出,植物的这种氮同位素差异和不同基因型玉米体内氮素的分配有关,子代户单4号将更多的氮素分配在了叶片部分,而父本803则将更多的氮素分配在了茎中。
Pardo等[10]对糖枫(sugar maple)和 山毛榉(A-merican Beech)根、茎和叶片中的氮同位素组成研究发现,其氮同位素组成的普遍规律为根系>茎>叶片。而Peuke等[13]在室内对德国的山毛榉的研究表明,氮同位素组成的普遍规律为叶片>茎>根系。Robinson[5]对野生大麦的研究也发现氮同位素组成为叶片>根系。本研究发现玉米杂交种及其亲本根、茎、叶中的氮同位素组成均表现为茎>叶片>根系,叶片中氮同位素组成大于根系氮同位素组成,与以往研究一致,不同的是,本研究中茎中氮同位素组成是最高的。说明在高氮肥条件下,玉米体内氮素的同化机制和其他植物具有机理上的差异性。由于目前还不能做到分析植物组织中不同氮素(有机氮、无机氮)的同位素组成,对于其差异性还难以从机理上进行解释。
3.2 杂交种及其亲本氮素分配的杂种优势效应
杂种优势的利用是作物育种上提高作物产量的主要手段之一,玉米是世界上杂种优势利用面积最大的作物[8]。对于本研究选择的实验材料,父本803为不抗旱型,母本天四为抗旱型,其子代户单4号为抗旱高产型。以往的研究者已经从根系吸水能力、根系的形态性状、水分利用等方面深入阐述了其子代的杂种优势效应[14~18]。本研究利用氮同位素技术进行的研究表明,无论是 Δδ15N(根-叶)还是 Δδ15N(根-茎),其绝对值均表现为户单4号<母本天四<父本803。这说明在氮素的吸收和同化过程中,父本803的分馏作用更大,而子代户单4号的分馏作用最小。分馏理论认为,分馏的产生往往是因为不同种类的氮素在传输和同化过程中活性不同导致,也就是植物对其利用的有效性差异导致,因此可以由此推断分馏作用越小则植物对氮素的利用率要更高一些,这也许就是子代具有杂种优势效应的原因之一。但是对于其机理上的原因还需要进一步深入研究才能够解释清楚。综合上述论述可以推断,在氮源相同的条件下,可以利用植物体的氮同位素组成研究植物的杂种优势效应。
4 结论
通过玉米研究杂交种及其亲本氮同位素组成的结果表明,在根、茎、叶中的氮同位素组成和氮含量明显不同,杂交种及其亲本体内氮同位素组成为茎>叶>根,这种差异性来源于不同植物在氮素的同化和分配上的差异性。对不同品种 Δδ15N(根-叶)和Δδ15N(根-茎)的研究显示,Δδ15N(根-叶)和 Δδ15N(根-茎)的绝对值均为父本803>母本天四>子代户单4号,说明这种差异性和玉米的杂种优势有关。本研究结果为进一步深入研究植物体内氮素的分配以及玉米的杂种优势提供了研究思路。
[1]许晨璐,孙晓梅,张守攻.基因差异表达与杂种优势形成机制探讨[J].遗传,2013,35(6):714 -726.
[2]张义荣,姚颖垠,彭惠茹,等.植物杂种优势形成的分子遗传机理研究进展[J].自然科学进展,2009,19(7):697-703.
[3]Evans RD.Physiological mechanisms influencing plant nitrogen isotope composition[J].Trends in Plant Science,2001,6(3):121 -126.
[4]Kolb KJ,Evans RD.Influence of nitrogen source and concentration on nitrogen isotope discrimination in two barley genotypes(Hordeum vulgare L.)[J].Plant Cell Environment,2001,26:1431 -1440.
[5]Robinson D,Handley LL,Scrimgeour CM,et al.Using stable isotope natural abundance(δ15N and δ13C)to integrate the stress responses of wild barley(Hordeum Spontaneum C.Koch.)genotypes[J].J Exp Bot,2000,51:41-50.
[6]Warner RL,Huffaker RC.Nitrate transport is independent of NADH and NAD(P)H nitrate reductases in barley seedings[J].Plant Physiology,1989,91,947 -953.
[7]王泽立,李新征,郭庆法,等.玉米抗旱性遗传与育种[J].玉米科学,1998,6(3):9 -13.
[8]李凤海.杂交当代玉米杂交效应初探[J].辽宁农业科学,1999(2):32-38.
[9]Hochholdinger F,Hoecker N.Towards the molecular basis of heterosis[J].Trends in Plant Science,2007,12(9):427-432.
[10]Pardo LH,Semaoune P,Schaberg PG,et al.Patterns in δ15N in roots,stems,and leaves of sugar maple and A-merican beech seedlings,saplings,and mature trees[J].Biogeochemistry,2013,112:275 – 291.
[11]Evans RD,Bloom AJ,Sukrapanna SS,et al.Nitrogen isotope compostion of tomato(Lycopersicon esculentum Mill.Cv.T-5)grown under ammonium or nitrate nutrition[J].Plant Cell Environ,1996,19:1317 -1323.
[12]Mariotti A,Mariotti F,Champigny ML,et al.Nitrogen isotope fractionation associated with nitrate reductase activity and uptake of nitrate by pearl millet[J].Plant Physiol,1982,69:880 -884.
[13]Peuke AD,Gessler A,Rennenberg H.The effect of drought on C and N stable isotope in different fractions of leaves,stems and roots of sensitive of and tolerant beech ecotypes[J].Plant Cell Environ,2006,29(5):823 -835.
[14]王周锋,张岁岐,刘小芳.玉米根系水流导度差异及其与解剖结构的关系[J].应用生态学报,2005,16(12):2349-2352.
[15]慕自新,张岁岐,郝文芳,等.玉米根系形态性状和空间分布对水分利用效率的调控[J].生态学报,2005,25(11):2895-2900.
[16]慕自新,张岁岐,梁爱华,等.玉米整株根系水导与其表型抗旱性的关系[J].作物学报,2005,31(2):203-208.
[17]吴安慧,张岁岐,邓西平,等.水分亏缺下玉米根系ZmPIP1亚族基因的表达[J].植物生理与分子生物学学报,2006,32(5):557 -562.
[18]周小平,张岁岐,杨晓青,等.玉米根系活力杂种优势及其与光合特性的关系[J].西北农业学报,2008,17(4):84-90.
猜你喜欢
西南农业学报(2022年5期)2022-06-06
蔬菜(2021年7期)2021-11-27
今日农业(2021年14期)2021-11-25
中国糖料(2021年3期)2021-07-13
桉树科技(2020年2期)2020-07-16
四川蚕业(2020年2期)2020-07-10
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
科学导报(2018年47期)2018-05-14
种子科技(2014年8期)2014-05-18
