
酪蛋白降解对牦牛乳硬质干酪苦味的影响
2015-12-20 07:56宋雪梅
食品科学 2015年19期
王 玲,梁 琪,宋雪梅,张 炎
(甘肃农业大学食品科学与工程学院,甘肃省功能乳品工程实验室,甘肃 兰州 730070)
酪蛋白降解对牦牛乳硬质干酪苦味的影响
王 玲,梁 琪*,宋雪梅,张 炎
(甘肃农业大学食品科学与工程学院,甘肃省功能乳品工程实验室,甘肃 兰州 730070)
针对牦牛乳硬质干酪的苦味缺陷,分别以小牛皱胃酶、微生物凝乳酶和木瓜蛋白酶制作的牦牛乳硬质干酪为研究对象,利用尿素聚丙烯酰胺凝胶电泳,研究牦牛乳硬质干酪pH 4.6水不溶性酪蛋白的降解程度,且对成熟过程中的牦牛乳硬质干酪苦味进行感官评价,探究牦牛乳硬质干酪pH 4.6水不溶性酪蛋白降解对其苦味的影响。结果表明:牦牛乳硬质干酪在成熟期间酪蛋白发生了明显的降解,且αs-酪蛋白均比β-酪蛋白降解速率快。经尿素聚丙烯酰胺凝胶电泳分离后,发现木瓜蛋白酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白在Pre-αs-酪蛋白区域有较强的蛋白带。木瓜蛋白酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白中αs-酪蛋白和β-酪蛋白降解程度均显著或极显著高于微生物凝乳酶和小牛皱胃酶制作的牦牛乳硬质干酪(P<0.05或P<0.01),木瓜蛋白酶制作的牦牛乳硬质干酪的苦味值极显著高于微生物凝乳酶和小牛皱胃酶制作的牦牛乳硬质干酪的苦味值(P<0.01),通过主成分分析得出3 种凝乳酶制作牦牛乳硬质干酪的苦味值和未降解β-酪蛋白和αs-酪蛋白含量成极显著负相关。这为控制牦牛乳硬质干酪品质提供了理论参考。
牦牛乳硬质干酪;酪蛋白降解;尿素聚丙烯酰胺凝胶电泳;苦味;主成分分析
牦牛乳是中国青藏高原地区的特色资源,蛋白质含量为5.6%,比荷斯坦牛乳高100.7%,而乳蛋白的组成以酪蛋白(casein,CN)为主,含量为84%[1],且主要是αs1-酪蛋白、αs2-酪蛋白、β-酪蛋白和κ-酪蛋白。酪蛋白的降解不仅影响干酪的产量和质地,一定程度上对干酪的风味也有较大的影响。大多数干酪成熟过程中,残留的凝乳酶促使酪蛋白降解,凝乳酶过度水解酪蛋白会产生苦味[2-4],苦味是蛋白质水解不平衡产生苦味肽的积累结果。已有大量从凝乳酶降解酪蛋白的产物αs1-酪蛋白和β-酪蛋白中分离出苦味肽的报道[5]。Lowrie等[6]研究发现切达干酪中的苦味是由酪蛋白降解产生的部分寡肽引起的,因为它们含有大量的疏水性残基。酪蛋白降解产物αs-酪蛋白和β-酪蛋白这两种酪蛋白含有较高的疏水性氨基酸,因此,这两种酪蛋白是苦味肽可能的来源。不同凝乳酶对牦牛乳硬质干酪酪蛋白降解程度不同。与动物凝乳酶相比,植物和微生物凝乳酶对β-酪蛋白降解影响更大[7-11]。许多研究发现同种凝乳酶水解αs-酪蛋白比β-酪蛋白降解程度更大。国外就动物凝乳酶制作干酪的酪蛋白降解研究报道较多,但对动物凝乳酶、微生物凝乳酶和植物凝乳酶分别制作干酪的酪蛋白降解研究甚少。
近些年研究发现尿素聚丙烯酰胺凝胶电泳(urea polyacrylamide gel electrophoresis,urea-PAGE)能够分析干酪酪蛋白降解程度,且能定量测定干酪成熟过程中酪蛋白的降解程度[12]。与传统的分析手段(高效液相色谱和聚丙烯酰胺凝胶电泳)相比,尿素聚丙烯酰胺凝胶电泳能在较低的凝胶浓度下达到很好的分离效果,能够很好地分析干酪成熟过程中酪蛋白降解的小分子多肽物质。我国正处于干酪发展的起步阶段,利用尿素聚丙烯酰胺凝胶电泳对不同凝乳酶制作干酪中酪蛋白降解产生苦味的研究尚未见报道,尤其是针对牦牛乳硬质干酪酪蛋白降解的相关研究还处于空白阶段。因此,本实验利用尿素聚丙烯酰胺凝胶电泳研究不同凝乳酶(动物、微生物和植物凝乳酶)分别制作牦牛乳硬质干酪酪蛋白降解程度对干酪苦味的影响,旨在对牦牛乳硬质干酪酪蛋白的降解规律有较为明确的认识,以期为今后指导牦牛乳干酪生产和控制干酪质量提供一定的科学依据。
1 材料与方法
1.1 材料与试剂
小牛皱胃酶(酶活力10 000 U/g) 兰州百灵生物技术有限公司;微生物凝乳酶(酶活力20 000 U/g) 甘肃华羚生物技术研究中心;木瓜蛋白酶(酶活力80 000 U/g)上海源叶生物科技有限公司;嗜热发酵剂:保加利亚乳杆菌、嗜热链球菌 丹麦丹尼斯克公司;αs-酪蛋白、β-酪蛋白 美国Sigma-Aldrich公司。
三羟基氨基甲烷、丙烯酰胺、N,N’-甲叉双丙烯酰胺、过硫酸铵、四甲基乙二胺、考马斯亮蓝G-250、溴酚蓝、盐酸、甲醇、冰乙酸等其他常用试剂均为分析纯。
1.2 仪器与设备
125L干酪压榨槽 上海翰强仪器设备厂;E-201-9 pH计 上海仪电科学仪器公司;TGL-20台式高速冷冻离心机 中国长沙湘仪离心机仪器有限公司;HWS26型电热恒温水浴锅 上海一恒科技有限公司;真空冷冻干燥机 上海姚氏仪器设备厂;电泳仪和电泳槽 美国Bio-Rad公司;Tanon Gis2010凝胶图像分析系统 上海天能科技有限公司。
1.3 方法
分别制作3 种酶活力相当的凝乳酶(小牛皱胃酶、微生物凝乳酶、木瓜蛋白酶)牦牛乳硬质干酪,用尿素聚丙烯酰胺凝胶电泳分别测定贮藏在10 ℃条件下的6 个成熟阶段(30、60、90、120、150、180 d)中3 种凝乳酶牦牛乳硬质干酪的pH 4.6水不溶性酪蛋白的降解程度,并对成熟过程中的干酪苦味进行感官评价。
1.3.1 牦牛乳硬质干酪制作工艺
原料乳→过滤→检验→巴氏杀菌(63 ℃,30 min)→冷却(35 ℃)→添加发酵剂(0.062 5 g/100 mL)→添加CaCl2(0.03 g/100 mL)→添加凝乳酶→凝乳→切割→二次加热→排乳清→搅拌、加盐(凝块的2%)→堆酿(30 min)→压榨成型(4~5 h)→真空包装→成熟
1.3.2 干酪pH 4.6水不溶性酪蛋白的制备
分别称取5 g干酪,加入15 mL去离子水均质。混合物于40 ℃条件下放置1 h,室温条件下放置15 min。在磁力搅拌的作用下,用1 mol/L盐酸调混合物至pH 4.6。混合物在4 ℃、4 000×g条件下离心20 min。沉淀用1 mmol/L醋酸铵(pH 4.6)冲洗一次,在4 ℃、3 000×g条件下离心10 min,收集沉淀进行真空冷冻干燥,干燥的粉末称质量,样品冷冻保存备用[13]。
1.3.3 尿素聚丙烯酰胺凝胶电泳[14-15]
分离胶:T 12.5%,C 4%;分离胶缓冲溶液:4.6 g三羟基氨基甲烷和26 g尿素,加入0.4 mL的浓盐酸,调节pH值至8.9,定容至100 mL。12 g丙烯酰胺、0.5 g N,N’-甲叉双丙烯酰胺、50 μL四甲基乙二胺和40 mg过硫酸铵溶于100 mL pH 8.9的分离胶缓冲溶液。
浓缩胶:T 4.2%,C 5%;浓缩胶缓冲溶液:0.75 g三羟基氨基甲烷和26 g尿素,加入0.4 mL的浓盐酸,调节pH值至7.6,定容至100 mL。2 g丙烯酰胺、0.1 g N,N’-甲叉双丙烯酰胺、25 μL四甲基乙二胺和20 mg过硫酸铵溶于100 mL pH 8.9的浓缩胶缓冲溶液。
电泳缓冲液:15 g三羟基氨基甲烷和73 g甘氨酸溶解在5 L的水中,预电泳电压为80 V,样品进入分离胶后调到120 V。
样品溶解液:0.75 g三羟基氨基甲烷和28 g尿素,加入0.4 mL的浓盐酸定容至100 mL。样品处理:分别取5 mg冻干样品溶解到3 mL的样品溶解液中,滴入2 滴溴酚蓝溶液,充分混匀,样品在沸水中煮沸5 min,待冷却后上样。电泳结束后用考马斯亮蓝染色,甲醇冰醋酸溶液脱色。用天能图像分析系统对电泳图片进行扫描分析,得谱带的积分光密度值和样品中各蛋白的百分含量。
1.3.4 牦牛乳硬质干酪苦味值的测定
感官评定小组由经过筛选的9 人组成(男女比例为4∶5,均为不吸烟者),品尝时,评价员用蒸馏水漱口,取1.5 cm×1.5 cm×1.5 cm大小的干酪置于口中,5~10 s后吐出。根据品尝的情况,然后按照表1的评分标准对每种干酪打分,评分值就是苦味值,得出的平均值表示干酪的苦味[16]。
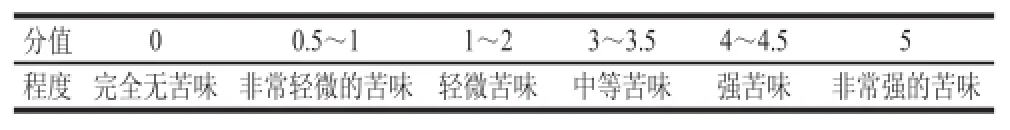
表1 苦味值的评分标准Table 1 Evaluation standards of bitterness
感官评价之前,分别取干酪样品在4 ℃过夜解冻,然后21 ℃放置1 h。以不同质量浓度的硫酸奎宁溶液为参比(0.002 3、0.004 6、0.009 4、0.018 8 g/L),对干酪的苦味进行感官评价。
1.4 数据处理
实验所得数据均用SPSS17.0统计分析软件进行处理,分别对凝乳酶种类和成熟时间进行单因素方差分析,LSD法进行差异显著性分析。对牦牛乳硬质干酪酪蛋白降解指标和干酪苦味值做了相关性分析,且对凝乳酶种类和成熟时间进行主成分分析。
2 结果与分析
2.1 尿素聚丙烯酰胺凝胶电泳结果

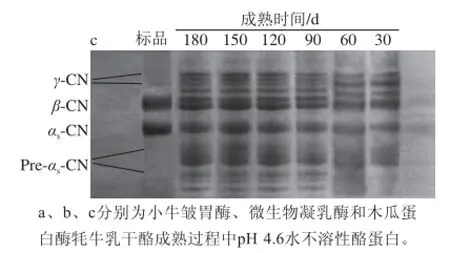
图1 3 种凝乳酶牦牛乳干酪成熟过程中pH 4.6水不溶性酪蛋白变化的尿素聚丙烯酰胺凝胶电泳图Fig.1 Urea-PAGE of pH 4.6-insoluble casein from yak milk hard cheeses prepared w ith calf rennet, microbial rennet and papain throughout ripening
由图1可知,干酪pH 4.6水不溶性酪蛋白分子质量由大到小划分为4 个条带:γ-CN、β-CN、αs-CN、Pre-αs-CN。3 种凝乳酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白降解的电泳条带不一致,原因是3 种凝乳酶对酪蛋白特定的酶切位点不同引起的[17]。木瓜蛋白酶牦牛乳硬质干酪的αs-CN和β-CN降解高于微生物和小牛皱胃酶,这与Sanjuán等[18]的研究结果一致。3 种凝乳酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白中αs-CN均比β-CN降解强烈,这主要是因为αs-CN的降解是由凝乳酶的作用引起的,β-CN的降解是由血纤维素蛋白溶酶引起的,也受到凝乳酶小幅度的降解,而排乳清时把纤维素蛋白溶酶的激活剂和抑制剂排除了,使得β-CN的降解程度较弱[18]。木瓜蛋白酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白在Pre-αs-CN区域有较强的蛋白带,这说明木瓜蛋白酶制作的牦牛乳硬质干酪中Pre-αs-CN区域的电泳迁移率最高,其次是微生物凝乳酶,小牛皱胃酶牦牛乳硬质干酪较弱。这是由于木瓜蛋白酶对αs-CN的降解比小牛皱胃酶和微生物凝乳酶更强烈。另外,木瓜蛋白酶和微生物凝乳酶牦牛乳硬质干酪中,Pre-αs-CN和γ-CN区域条带的颜色随着干酪的成熟逐渐加深。
2.2 牦牛乳硬质干酪未降解β-酪蛋白水平的变化

图2 3 种凝乳酶干酪成熟过程中未水解β-酪蛋白水平的变化Fig.2 Changes in residual β-CN of hard yak milk cheeses prepared with calf rennet, microbial rennet and papain during ri pening
由图2可知,牦牛乳硬质干酪成熟过程中,随成熟的进行,3 种凝乳酶对β-CN的降解速率趋于平缓。成熟终点时,小牛皱胃酶、微生物凝乳酶和木瓜蛋白酶制作的牦牛乳硬质干酪未水解β-CN含量分别是31.14%、25.74% 和19.78%。由方差分析可知,同一成熟时间,3 种凝乳酶对牦牛乳硬质干酪未水解β-CN含量影响极显著(P<0.01);同一凝乳酶,不同成熟时间对牦牛乳硬质干酪未水解β-CN的含量影响极显著(P<0.01)。牦牛乳硬质干酪整个成熟过程中,木瓜蛋白酶对β-CN降解极显著高于微生物凝乳酶和小牛皱胃酶(P<0.01),这与Galán等[19]研究荆棘蓟制作的羊乳干酪β-CN降解极显著高于小牛皱胃酶制作干酪的结果一致,这可能与引起木瓜蛋白酶牦牛乳硬质干酪的苦味较强的有关。微生物凝乳酶使β-CN的降解显著高于小牛皱胃酶(P<0.01),这和Lawrence等[17]研究米黑毛霉切达干酪β-CN的降解比小牛皱胃酶制作干酪强烈的结果吻合。牦牛乳硬质干酪成熟过程中,β-CN主要由受血纤溶蛋白酶影响,血纤溶蛋白酶首先使β-CN降解,然后被微生物蛋白酶缓慢水解为γ1[β-CN(f29~209)]、γ2[β-CN(f106~209)]、γ3[β-CN(f108~209)]。随成熟时间的延长,β-CN逐渐降解,γ-CN含量增加,干酪中残留的凝乳酶也会导致β-CN的微小降解。
2.3 牦牛乳硬质干酪未降解αs-酪蛋白水平的变化
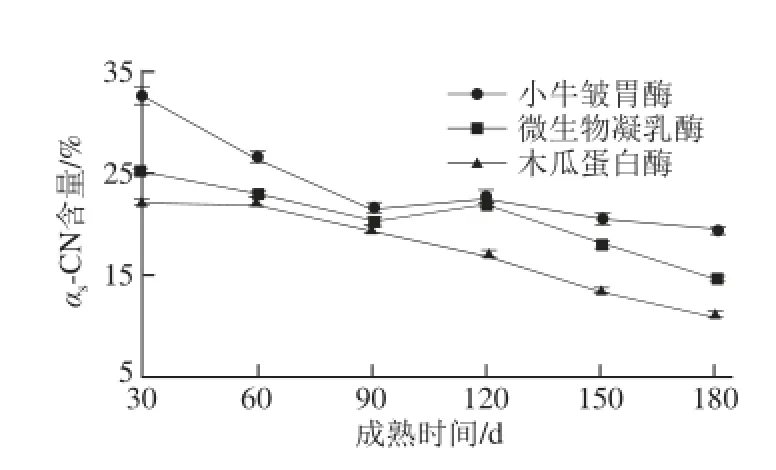
图3 3 种凝乳酶干酪成熟过程中未水解αs-酪蛋白的变化Fig.3 Changes in residual αs-CN of hard yak milk cheeses prepared with calf rennet, microbial rennet and papain during ripening
由图3可知,3 种凝乳酶牦牛乳硬质干酪成熟过程中,凝乳酶种类对牦牛乳硬质干酪的αs-CN降解有显著影响(P<0.05),成熟时间对其也有极显著影响(P<0.01 )。成熟30 d之后,木瓜蛋白酶牦牛乳硬质干酪αs-CN含量显著低于其他两种凝乳酶干酪(P<0.05)。血纤维素蛋白酶、植物凝乳酶和动物凝乳酶依次使αs-CN降解,这就使得木瓜蛋白酶牦牛乳硬质干酪的αs-CN降解程度显著高于小牛皱胃酶干酪[19]。αs-CN在Phe23-Phe24处断裂,形成αs1-CN(f1~23)和αs2-CN(f24~199)[20-25]小片段。在成熟180 d时,小牛皱胃酶、微生物凝乳酶和木瓜蛋白酶制作的牦牛乳硬质干酪未被水解的αs-CN含量分别为19.34%、14.43%和10.89%。3 种凝乳酶制作的牦牛乳硬质干酪成熟过程中,αs-CN和β-CN的降解程度并不相同,αs-CN的总体降解程度要大于β-CN。Visser[25]研究表明,在干酪Urea-PAGE电泳中,酪蛋白的降解速率主要是凝乳酶和乳中纤溶酶作用的结果。一般认为,酪蛋白的初级降解主要是凝乳酶的作用。
2.4 牦牛乳硬质干酪未降解αs-CN/β-CN值的变化
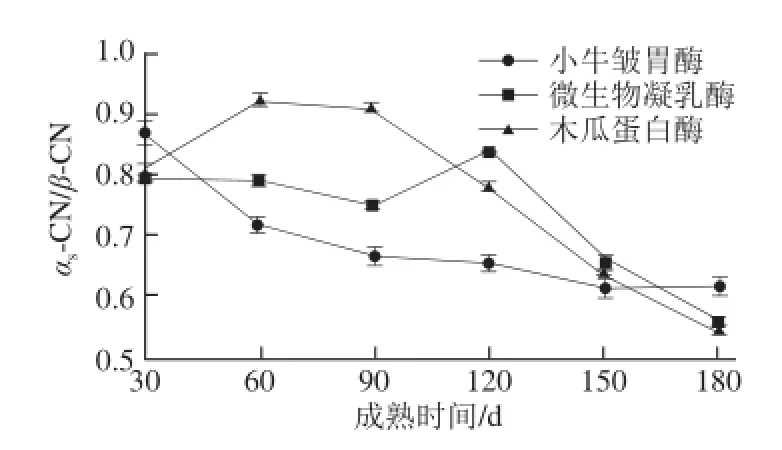
图4 3 种凝乳酶牦牛乳硬质干酪成熟过程中未水解αs--CCNN//β-CCNN值变化Fig.4 Changes in residual αs-C N/β-CN of yak milk hard cheeses prepared with calf rennet, microbial rennet and papain during ripening
由图4可知,成熟至180 d时,小牛皱胃酶、微生物凝乳酶和木瓜蛋白酶牦牛乳硬质干酪αs-CN/β-CN值分别从0.87、0.79和0.81降低到0.62、0.56和0.55。木瓜蛋白酶牦牛乳硬质干酪αs-CN/β-CN值最低,这与Fernández-Salguero等[26]研究动物凝乳酶和植物凝乳酶羊乳干酪报道的αs-CN/β-CN值从0.9%降低到0.5%一致。由方差分析可知,同一成熟时间,3 种凝乳酶牦牛乳硬质干酪未水解αs-CN/β-CN值差异显著(P<0.05)。成熟90 d之前,小牛皱胃酶牦牛乳硬质干酪未水解αs-CN/β-CN值差异显著(P<0.05),之后不显著(P>0.05)。成熟前90 d,微生物凝乳酶牦牛乳硬质干酪未水解αs-CN/β-CN值差异不显著(P>0.05),后期差异显著(P<0.05)。由此可以看出成熟时间和凝乳酶种类对牦牛乳硬质干酪未水解αs-CN/β-CN值有显著的影响。
2.5 牦牛乳硬质干酪苦味感官评价

图5 3 种凝乳酶牦牛乳硬质干酪苦味感官评价Fig.5 Sensory evaluation of the bitterness of hard yak milk cheeses prepared with calf rennet, microbial rennet and papain
由图5可知,随成熟时间的延长,3 种凝乳酶牦牛乳硬质干酪的苦味均呈缓慢加重趋势。成熟前90 d,同一种凝乳酶牦牛乳硬质干酪的苦味值差异不显著(P>0.05),成熟90 d之后,差异显著(P<0.05)。整个成熟过程中,凝乳酶种类对牦牛乳硬质干酪苦味值影响差异极显著(P<0.01)。牦牛乳硬质干酪成熟180 d时,小牛皱胃酶、微生物凝乳酶和木瓜蛋白酶牦牛乳硬质干酪苦味值分别从1.5、2和3升高到3、4和5。
2.6 牦牛乳硬质干酪酪蛋白降解片段与苦味值的相关性分析
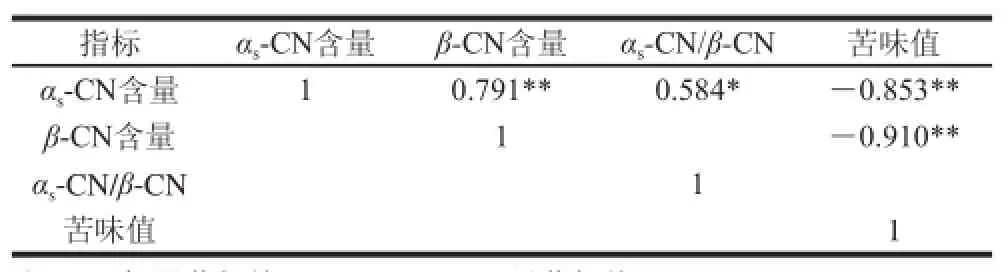
表2 牦牛乳硬质干酪酪蛋白降解指标与苦味值的相关性Table 2 Correlation between bitterness and casein degradation indices of hard yak milk cheese
由表2可知,牦牛乳硬质干酪未降解的αs-CN含量和β-CN含量成极显著正相关(r=0.791)、苦味值成极显著负相关(r=-0.853),且与αs-CN/β-CN值成显著正相关(r=0.584)。牦牛乳硬质干酪未降解的β-CN含量与苦味值成极显著负相关(r=-0.910)。未降解的β-CN含量对牦牛乳硬质干酪的苦味值的影响最大,其次是αs-CN含量,未降解酪蛋白αs-CN/β-CN值和牦牛乳硬质干酪的苦味值无相关性。
2.7 牦牛乳硬质干酪酪蛋白片段的主成分分析

图6 3 种凝乳酶牦牛乳硬质干酪成熟过程中酪蛋白片段的主成分得分图Fig.6 Score plots obtained from principal component analysis for casein fractions from hard yak milk cheeses prepared with calf rennet, microbial rennet and papain
由图6可知,第一主成分占累计贡献率的65.79%,反映了凝乳酶种类对未降解αs-CN含量差异的显著性有极大的贡献,但凝乳酶种类对未降解β-CN含量差异的显著性也有贡献。第二主成分占累计贡献率的33.95%,反映了成熟时间对牦牛乳硬质干酪β-CN含量的影响。在牦牛乳硬质干酪成熟180 d时,小牛皱胃酶牦牛乳硬质干酪中未降解β-CN的含量最高。同理,成熟时间对未降解αs-CN含量和αs-CN/β-CN值的差异显著性有极大的贡献。小牛皱胃酶牦牛乳硬质干酪pH 4.6水不溶性酪蛋白部分主要分布在PC 1的评分正值附近、PC 2的评分负值附近,而微生物凝乳酶和木瓜蛋白酶制作的牦牛乳硬质干酪pH 4.6水不溶性酪蛋白部分主要分布在PC 1的评分正值附近、PC 2的评分正值附近。说明木瓜蛋白酶和微生物凝乳酶对牦牛乳硬质干酪αs-CN和β-CN降解速率更大。
3 结 论
苦味缺陷是牦牛乳硬质干酪优质资源开发的制约因素。干酪成熟过程中,残留凝乳酶影响干酪酪蛋白的降解,其降解程度直接关系着干酪的苦味。通过研究不同凝乳酶制作的牦牛乳硬质干酪成熟期间酪蛋白的降解规律,发现不同凝乳酶牦牛乳硬质干酪酪蛋白中未降解的αs-CN、β-CN含量以及干酪苦味值差异均极显著(P<0.01),且牦牛乳硬质干酪酪蛋白中αs-CN均比β-CN降解强烈。木瓜凝乳酶制作的牦牛乳硬质干酪酪蛋白中的αs-CN和β-CN降解程度显著或极显著高于微生物凝乳酶和小牛皱胃酶牦牛乳硬质干酪(P<0.05或P<0.01),且β-CN的降解对牦牛乳硬质干酪的苦味值的影响最大,其次是αs-CN,αs-CN/β-CN值与干酪的苦味无关,表明干酪中残留的凝乳酶降解酪蛋白中的β-CN产生苦味肽,使干酪产生苦味。本实验为调控牦牛乳硬质干酪风味缺陷提供了一定的理论依据,在干酪生产中应选择蛋白酶活力较小而凝乳能力较强的凝乳酶和能够降解苦味肽的外肽酶。由于引起干酪苦味肽形成的原因极其复杂,导致引起干酪苦味的物质仍然不是非常清楚,因此,干酪中苦味的产生机理及苦味成分的组成还有待进一步研究。
[1] 杨楠, 梁琪, 杨敏, 等. 牦牛脱脂乳中酪蛋白受热处理温度及时间影响的研究[J]. 食品工业科技, 2013, 34(5): 121-125.
[2] VICENTE M S, IBANEZ F C, BARCINA Y, et al. Casein breakdown during ripening of Idiazabal cheese: influence of starter and rennet type[J]. Journal of the Science of Food and Agriculture, 2001, 81(2): 210-215.
[3] PINO A, PRADOS F, GALAN E, et al. Proteolysis during the ripening of goats’ milk cheese made with p lant coagulant or calf rennet[J]. Food Research International, 2009, 42(3): 324-330.
[4] IRIGOYEN A, IZCO J M, IBANEZ F C, et al. Evaluation of the effect of rennet type on casein proteolysis in an ovine milk cheese by means of capillary electrophoresis[J]. Journal of Chromatography A, 2000, 881(1): 59-67.
[5] VALÉRIE G, STEFANIA C, CONCETTA P, et al. Uncommonly thorough hydrolysis of peptides during ripening of Ragusano cheese revealed by tandem mass spectrometry[J]. Journal of Agricultural and Food Chemistry, 2011, 59(23): 12443-12452.
[6] LOWRIE R J, LAWRENCE R C. Cheddar cheese flavour. IV. A new hypothesis to account for the development of bitterness[J]. International Journal of Dairy Technology, 1972, 37(3): 113-125.
[7] TEJADA L, ABELLÁN A, CAYUELA J M, et al. Proteolysis in goats’milk cheese made with calf rennet and plant coagulant[J]. International Dairy Journal, 2008, 18(2): 139-146.
[8] ŞENGÜL M, ERKAYA T, DERVIŞOĞLU M, et al. Compositional, biochemical and textural changes during ripening of Tulum cheese made with different coagulants[J]. International Journal of Dairy Technology, 2014, 67(3): 373-383.
[9] YA SAR K, GUZELER N. Effects of coagulant type on the physicochemical and organoleptic properties of Kashar cheese[J]. International Journal of Dairy Technology, 2011, 64(3): 372-379.
[10] SOUSA M J, MALCATA F X. Advances in the role of a plant coagulant (Cynara cardunculus) in vitro and during ripening of cheeses from several milk species[J]. Le Lait, 2002, 82(2): 151-170.
[11] HAYALOGLU A A, BRECHANY E Y, DEEGAN K C, et al. Characterization of the chemistry, bi ochemistry and volatile profile of Kufl u cheese, a mou ld-ripened variety[J]. LWT-Food Science and Technology, 2008, 41(7): 1323-1334.
[12] MCSWEENEY P L H, FOX P F. Chemical methods for the characterization of proteolysis in cheese during ripening[J]. Le Lait, 1997, 77(1): 41-76.
[13] HAYALOGLU A A, GUVEN M, FOX P F. Proteolysis in Turkish White-brined cheese ma de with defined strains of Lactococcus[J]. International Dairy Journal, 2004, 14(7): 599-610.
[14] ANDREWS A T. Proteinases in normal bovine milk and their action on caseins[J]. Journal of Dairy Research, 1983, 50(2): 45-55.
[15] SHALABI S I, FOX P F. Electrophoretic analysis of cheese, comparison of methods[J]. Irish Journal of Food Science and Technology, 1987, 11(3): 135-151.
[16] KUKMAN I L, ZELENIK-BLATNIK M, ABRAM V. Isolation of low-molecular-mass hydrophobic bitter peptides in soybean protein hydrolysates by reversed-phase high-performance liquid chromatography[J]. Journal of Chromatography A, 1995, 704(1): 113-120.
[17] LAWRENCE R C, CREAMER L K, GILLES J. Texture development during cheese ripening[J]. Journal of Dairy Science, 1987, 70(8): 1748-1760.
[18] SANJUÁN E, MILLÁN R, SAAVEDRA P, et al. Infl uence of anim al and vegetable rennet on the physicochemical characteristics of Los Pedroches cheese during ripening[J]. Food Chemistry, 2002, 78(3): 281-289.
[19] GALÁN E, PRADOS F, PINO A, et al. Infl uence of different amounts of vegetable coagulant from cardoon Cynara cardunculus and calf rennet on the proteolysis and sensory characteristics of cheeses made with sheep milk[J]. International Dairy Journal, 2008, 18(1): 93-98.
[20] GARCÍA V, ROVIRA S, TERUEL R, et al. Effect of vegetable coagulant, microbial coagulant and calf rennet on physicochemical, proteolysis, sensory and texture profi les of fresh goats cheese[J]. Dairy Science and Technology, 2012, 92(6): 691-707.
[21] LARSEN M D, KRISTIANSEN K R, HANSEN T K. Characterization of the proteolytic activity of starter cultures of Penicillium roqueforti for production of blue veined cheeses[J]. International Journal of Food Microbiology, 1998, 43(3): 215-221.
[22] 张娜, 郭庆启, 黄文秀, 等. 微胶囊化蛋白酶在干酪制备中的应用及干酪成熟过程中电泳分析[J]. 食品科学, 2014, 35(5): 134-138. doi: 10.7506/spkx1002-6630-201405027.
[23] BARAĆ M B, SMILJANIĆ M, PEŠIĆ M B, et al. Primary proteolysis of white brined goat cheese monitored by high molarity Tris buffer SDS-PAGE system[J]. Mljekarstvo, 2013, 63(3): 122-131.
[24] O’MAHONY J A, SOUSA M J, MCSWEENEY P L H. Proteolysis in miniature Cheddar-type cheeses made using blends of chymosin and Cynara cardunculus proteinases as coagulant[J]. International Journal of Dairy Technology, 2003, 56(1): 52-58.
[25] VISSER S. Proteolytic enzymes and their relation to cheese ripening and flavor: an overview[J]. Journal of Dairy Science, 1993, 76(1): 329-350.
[26] FERNÁNDEZ-SALGUERO J, SANJUÁN E. Infl uence of vegetable and animal rennet on proteolysis during ripening in ewes’ milk cheese[J]. Food Chemistry, 1999, 64(2): 177-183.
Effect of Casein Degradation on Bitterness of Hard Yak Milk Cheese
WANG Ling, LIANG Qi*, SONG Xuemei, ZHANG Yan (Functional Dairy Product Engineering Laboratory of Gansu, College of Food Science and Engineering, Gansu Agricultural University, Lanzhou 730070, China)
In view of the bitter taste defect in hard yak milk cheese, calf rennet, microbial rennet and papain were used to prepare yak milk hard cheese in the present study to explore the degradation degree of pH 4.6 water-insoluble casein of hard yak milk cheese, as evaluated by urea-polyacrylamide gel electrophoresis (urea-PAGE) and sensory evaluation of bitterness during the ripening period, as well as explore the effect of degradation degree of pH 4.6 water-insoluble casein on bitterness of hard yak milk cheese. The results indicated that hard yak milk cheese casein revealed signifi cant degradation during the ripening period; moreover, αs-casein underwent much more extensive and faster degradation than β-casein. The pH 4.6 water-insoluble casein of hard yak milk cheese made with papain showed higher i ntensity ban ds in the Pre-αs-casein region as shown by urea-PAGE, while the degradation degree of αs-casein and β-casein was signifi cantly higher than that of pH 4.6 water-insoluble casein from hard yak milk cheeses pre pared with microbial rennet and calf rennet (P < 0.05). The bitterness of hard yak milk cheese prepared with papain was signifi cantly higher than that obtained from microbial rennet and calf rennet (P < 0.01). These results suggested that the contents of residual αs-casein and β-casein showed a highly signifi cant negative correlation with bitterness for three yak milk hard cheeses by principal component analysis. This investigation can provided a theor etical basis for the quality control of hard yak milk cheese.
hard yak milk cheese; casein degradation; urea-polyacrylamide gel electrophoresis (urea-PAGE); bitterness; principal component analysis
TS252.53
A
1002-6630(2015)19-0001-06
10.7506/spkx1002-6630-201519001
2014-12-19
国家自然科学基金地区科学基金项目(31260383)
王玲(1988-),女,硕士研究生,研究方向为乳制品加工。E-mail:1041842191@qq.com
*通信作者:梁琪(1969-),女,教授,博士,研究方向为食品品质、乳品科学。E-mail:liangqi@gsau.edu.cn
猜你喜欢
中国饲料(2022年5期)2022-04-26
中国兽医杂志(2020年3期)2020-09-07
今日畜牧兽医(2020年6期)2020-02-15
卫生职业教育(2017年13期)2017-07-25
饲料博览(2017年8期)2017-04-04
中国酿造(2016年12期)2016-03-01
中国畜禽种业(2016年4期)2016-01-27
Asian Agricultural Research(2015年3期)2015-02-02
食品工业科技(2014年3期)2014-03-22
食品工业科技(2014年15期)2014-03-11
