
红榉不同种源种子形态性状变异
2015-12-21 00:50王旭军张日清许忠坤吴际友张玖荣
中南林业科技大学学报 2015年1期
王旭军,张日清,许忠坤,程 勇,吴际友,张玖荣
(1.湖南省林业科学院,湖南 长沙 410004;2.中南林业科技大学,湖南 长沙 410004;3.桑植县林业局,湖南 桑植 427100)
红榉不同种源种子形态性状变异
王旭军1,张日清2,许忠坤1,程 勇1,吴际友1,张玖荣3
(1.湖南省林业科学院,湖南 长沙 410004;2.中南林业科技大学,湖南 长沙 410004;3.桑植县林业局,湖南 桑植 427100)
研究了红榉自然分布区4个省份15个种源的种子长、宽、长宽比及千粒重等性状的变异。结果表明:(1)红榉种源间种子长、宽、长宽比和千粒重等性状的差异均达极显著水平,且种源水平上各性状的遗传力分别为0.901 8、0.921 0、0.922 1和0.900 8,单株水平上的遗传力分别为0.926 0、0.979 2、0.932 6和0.992 5,表明红榉种子形态性状存在着丰富的遗传变异,且受到中等程度以上的遗传控制;(2)相关分析表明,红榉种子长、宽及千粒重之间存在着极显著相关关系,且红榉种子性状无明显的经向和纬向变异,种子长主要受年均温和无霜期影响,种子宽主要受1月均温影响,而千粒重主要受年均温的影响;(3)聚类分析表明,红榉种子形态特征地理变异呈现区域板块变异模式和随机变异模式等。
红榉;种源;种子形态性状;变异
红榉为大叶榉Zelkova schneideriana Hand-Mazz.,为榆科榉属落叶大乔木树种,因其材色浅红而得名,属国家Ⅱ级重点保护野生植物。红榉生长较快,且材质坚硬有弹性,纹理美观有光泽,结构细致,为我国珍贵硬阔叶用材树种。同时,红榉树冠广阔,树形优美,叶色季相变化丰富,春叶嫩绿,夏叶深绿,秋叶橙红,观赏价值高,是深受人们喜爱的传统色叶园林树种。此外,红榉还是药用、化工原料等方面的重要原料树种[1-4]。但目前关于红榉的研究多集中于苗木繁育和丰产栽培等方面[5-9],而关于其遗传变异的研究报道较少[10]。物种分布愈广泛,其蕴含的遗传变异性也较大,而种子是其遗传变异的集中点[11]。研究表明,林木种子性状存在着一定的地理种源变异,往往与分布区的光照、温度、降水量及海拔、纬度等环境因素有关,而这些因素又通常是相互作用、相互影响的[12-13];而且对红榉种子地理变异研究可以作为一种早期的测定手段,在较短的时间内取得关于红榉地理变异在格局、大小与趋势等方面的一些重要资料,从而为红榉的遗传改良及种子生产和调拨提供参考[14-17]。
1 材料与方法
1.1 试验材料
供试的红榉种子于2012年10至11月分别采集于红榉自然分布区4个省的15个种源(见表1)。除湖南古丈和浙江桐乡为本地起源的红榉人工林外,其他林分均为天然林。由于每个天然种源林红榉数量少,且结实量也少,因此,在选优的基础上,为保证采到尽可能多的种子的前提下,每个种源地采种母树数量从5株到20株不等,且保证每个种源地各单株间距达50m以上。种子分单株采集,经自然干燥后置于通风阴凉处贮藏备用。
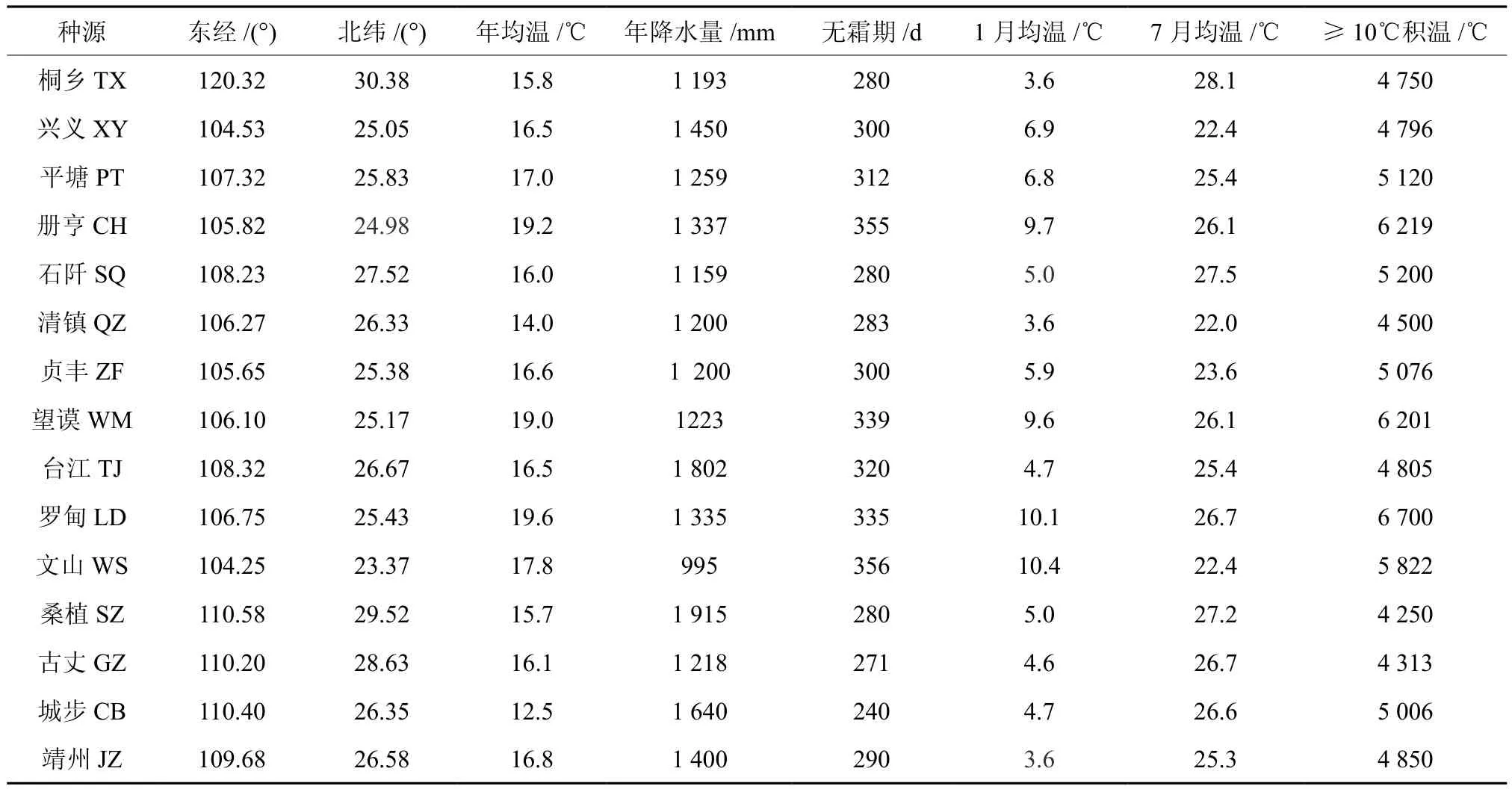
表1 不同红榉种源地理气候因子Table 1 Geographic meterological factors of different Z.schneideriana provenances
1.2 测定方法
种子形态性状等指标的测定以单株为单位,并以各种源内所有单株的均值来表示该种源种子形态性状的指标值。
每个红榉单株种子随机选取30粒,用电子游标卡尺测量种子长度和宽度等形态指标,以种子纵轴为长度,以腹面横向最大宽度为宽度,测量单位精确到0.01mm,并计算种子长宽比,3次重复。用百粒重法测定种子千粒重[18]。
1.3 数据分析
红榉种子原始数据文档用Microsoft of fi ce Excel2003整理,用SPSS16.0进行数据方差分析、相关分析和聚类分析等,而种源和家系遗传力采用公式H2=1-1/F,其中H2为性状遗传力,F为方差分析中的F检验值[19]。
2 结果与分析
2.1 不同种源红榉种子形态性状的变异
林木种子形态属一种较为稳定的性状,它比林木的根、茎、叶稳定性高[20]。对红榉种源间种子形态性状的统计分析(见表2)表明,红榉的平均种子长、宽、长宽比及千粒重分别为3.71mm、3.81mm、0.98和10.15g,且各形态性状指标的变异系数分别为3.74%、3.47%、3.28%和9.86%,均小于10%,说明红榉种源内种子形态性状比较稳定,变幅较小。不同种源间比较来看,种子最长的为城步种源,为4.35mm,最短的为望谟种源,为3.27mm,仅为城步种源的75%;种子最宽的为靖州种源,为4.60mm,最窄的也为望谟种源,为3.24mm,仅为靖州种源的70%;而种子长宽比最大的为城步种源,最小的为清镇种源,分别为1.12和0.78,这也说明红榉种子存在着圆形变异的趋势。而不同种源间千粒重比较来看,千粒重最大的为城步种源,达15.18g,最小的为望谟种源,仅为6.35g,还不及城步种源的一半。方差分析结果也表明,红榉种子各形态性状指标在种源间存在着极显著差异(见表2)。这意味着红榉种子形态性状在种源间存在着丰富的遗传变异,且红榉种源间种子长遗传力为0.901 8,种子宽的遗传力为0.921 0,长宽比的遗传力为0.922 1,千粒重的遗传力为0.900 8,表明这些性状受到较强的遗传控制,为红榉的遗传改良提供了可靠保障。
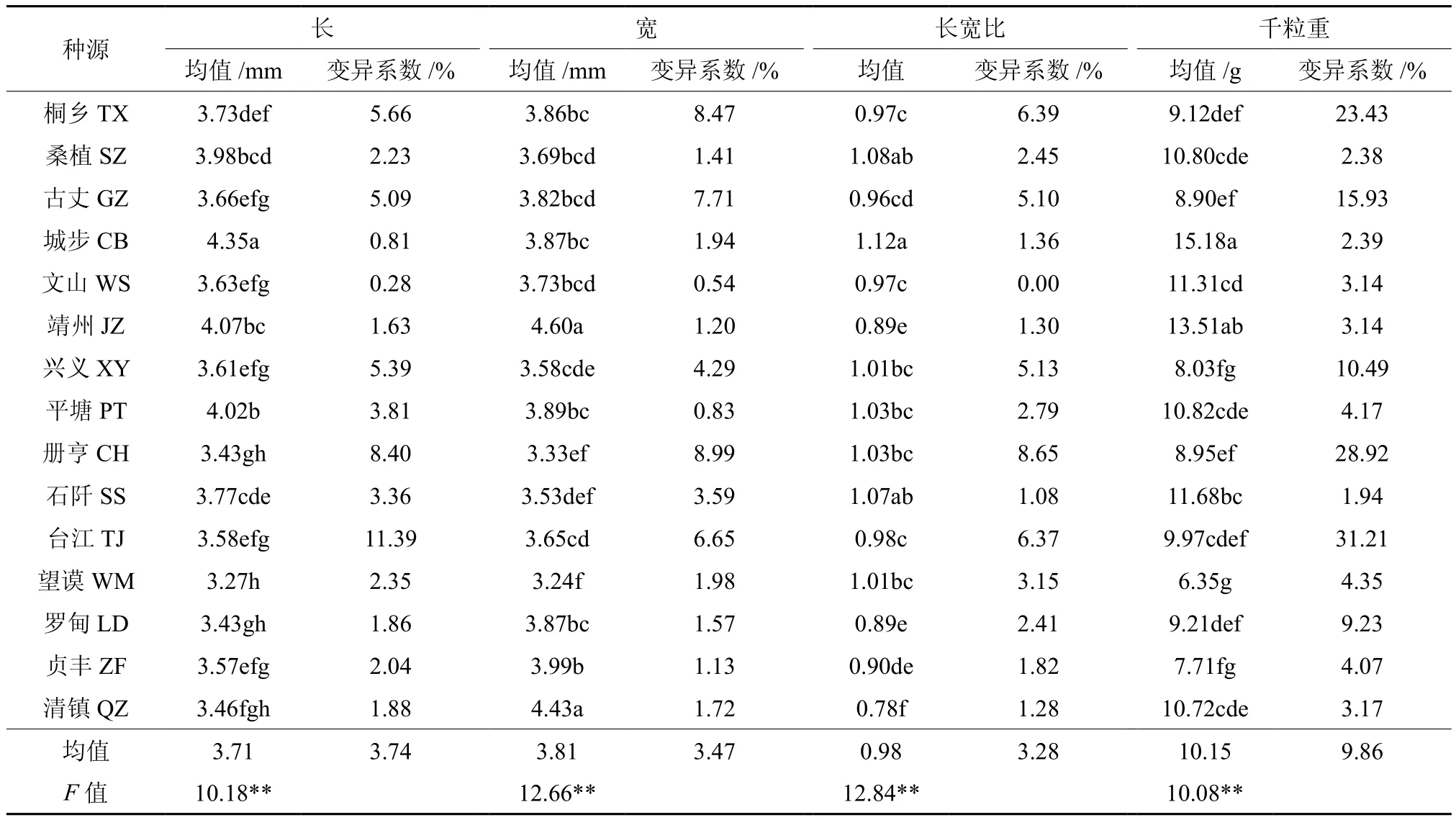
表2 不同种源红榉种子形态性状变异†Table 2 Variation of seed morphologic traits of different Z.schneideriana provenances
2.2 不同单株红榉种子形态性状的变异
对红榉单株间种子形态性状的统计分析(见表3)表明,红榉的平均种子长、宽、长宽比及千粒重分别为3.60mm、3.70mm、0.98和8.93g,且各形态性状指标的变异系数分别为2.83 %、1.92 %、2.84%和2.71 %,均小于10%,说明红榉单株内种子形态性状也比较稳定,变幅较小。不同单株间比较来看,种子长最长的为10号单株,达4.07mm,最短的为120号单株,仅3.15mm;种子宽最大的也为10号单株,为4.61mm,最窄的为114号单株,为3.00mm;种子长宽比最大的为116号单株,为1.16,最小的为129号单株,为0.78;千粒重最大的为65号单株,为13.38g,最轻的为114号单株,为5.71g,还不及65号单株的一半。方差分析表明,红榉各种子形态性状指标在不同单株间均存在极显著差异,这也说明红榉种子在单株水平上蕴藏着丰富的遗传变异,且红榉单株间种子长、种子宽、长宽比和千粒重的遗传力分别为0.926 0、0.979 2、0.932 6和0.992 5,表明这些性状也受到中等程度以上的遗传控制。
2.3 红榉种子形态性状的相关分析
探讨种子性状之间的相关关系,可以用来指导选择育种和林木种质资源保存中样本策略的制
定[21]。由表4可以看出,无论在种源水平还是单株水平上,红榉种子千粒重除与种子长宽比相关不显著外,与种子长、宽两个性状均达到了极显著正相关,表明种子的千粒重性状受种子长、种子宽影响很大,而且有随着种子长、宽的增大而显著递增趋势。因此,可以利用种子千粒重作为选择种子的间接指标;而种子长、宽与长宽比三个性状之间,两两相关也均达到了极显著水平。
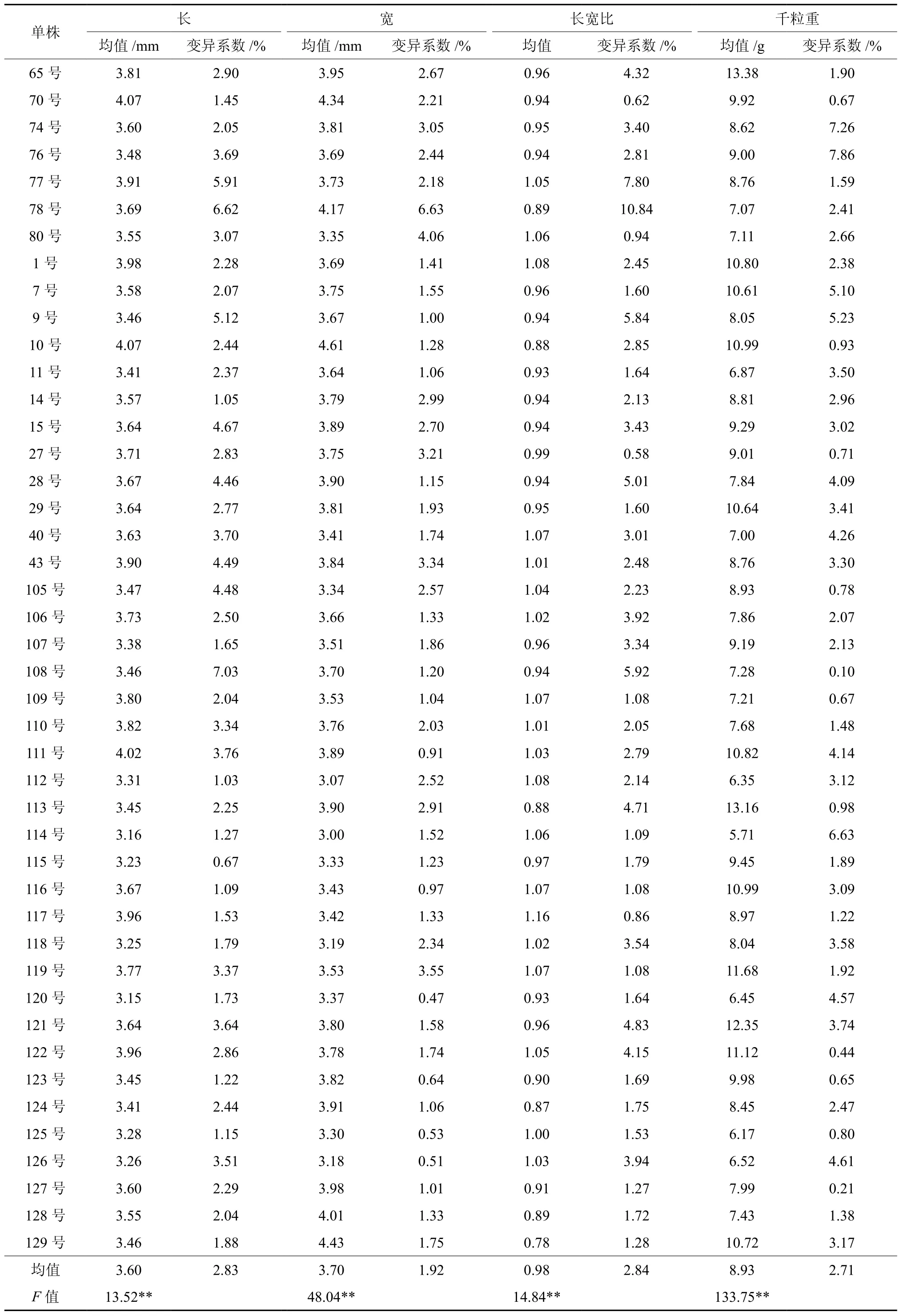
表3 不同单株红榉种子形态性状变异Table 3 Variation of seed morphologic traits of different Z.schneideriana individuals
2.4 不同种源红榉种子形态性状与地理气候因子的相关分析
通过对红榉种子性状与地理气候因子的相关分析,可以了解不同环境条件对种子性状发育的影响,为种子区划与良种筛选提供依据[22-25]。由表5可以看出,红榉种子各表型性状与经度和纬度呈较弱的正相关,说明经纬度对种子性状的影响不明显;年均温和无霜期与种子各性状都呈负相关,且与种子长、千粒重达到显著或极显著负相关;此外,1月均温对种子宽也达到显著相关水平,而其他因子与种子性状相关关系也不显著,对其影响不明显。总之,环境因素通过多因子综合作用和因子间交互作用引起红榉种子各形态特征变异。
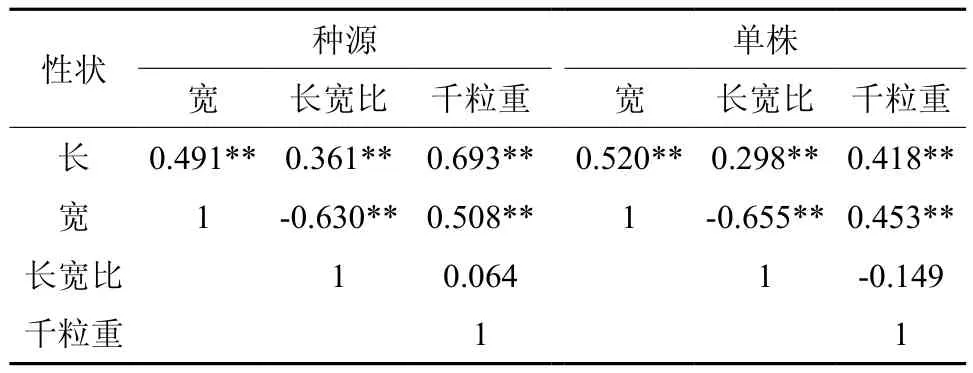
表4 红榉种子表型性状相关分析Table 4 Correlation coefficients among different morphologic traits
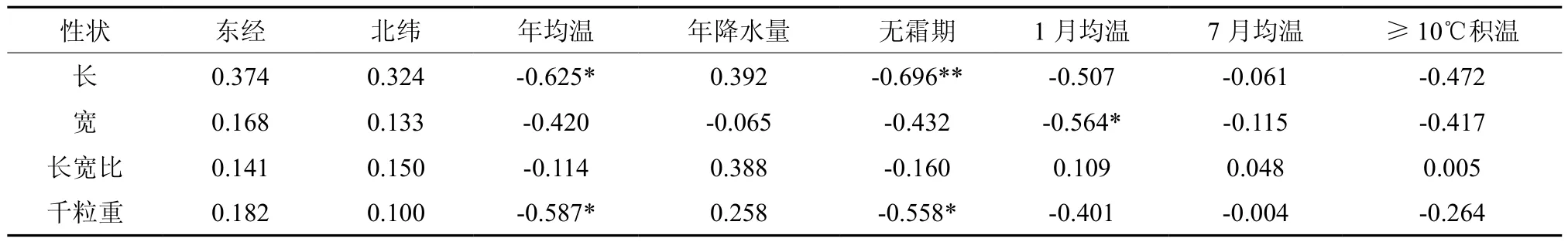
表5 红榉种子性状与地理气候因子的相关关系Table 5 Correlation coefficients between morphologic characters and geographical-meterological factors
2.5 不同种源红榉种子形态性状的聚类分析
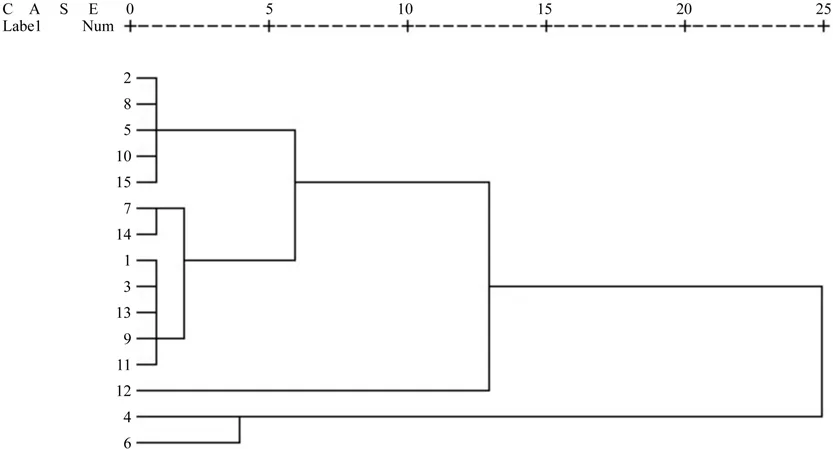
图1 红榉不同种源聚类树Fig.1 Dendrogram of different Z.schneideriana provenances
研究表明,以亲代特征为指标的地理变异模式与种源试验的结果基本相符,因此,对亲代表型性状进行聚类分析对于早期获得种源区划信息具有重要意义[26]。为了合理评价各红榉种源种子特性的差异并进行种源划分,以种子长、宽、长宽比和千粒重等为指标,采用欧式距离法对15个种源进行聚类分析(见图1)。由图1可知,可将红榉种源划分为四类,湖南桑植、贵州平塘、云南文山、贵州石阡、贵州清镇为一类;贵州兴义、贵州贞丰、浙江桐乡、湖南古丈、贵州罗甸、贵州册亨和贵州台江为一类;湖南城步和靖州为一类;而贵州望谟单独列为一类。由此分类结果可以看到,湖南城步和靖州地理位置较为接近,因而聚类为一类;而湖南桑植、云南文山及贵州平塘等地地理相隔遥远,且其年均温、降水量、无霜期等气候因子也相差较大,但聚类分析结果仍然将它们聚类为一类,同样的现象在浙江桐乡、湖南古丈等种源上也可以看到。广布种植物地理变异的形成与自然选择作用、基因流作用和基因飘动作用多种因素相关,在诸多因素综合作用下,植物会形成连续变异、区域板块变异以及随机变异等多种变异模式[27-28]。以上结果说明,红榉种子形态特征在部分地区呈现典型的区域板块化特征,部分地区呈现随机变异的特征。
3 结论与讨论
无论种源间还是单株间,红榉种子长、宽、长宽比及千粒重等性状均达到极显著差异。这说明红榉种源间(单株间)种子形态性状存在着丰富的遗传变异,遗传改良潜力巨大,为优良种源(单株)的选择提供了基础条件;且红榉种源间种子长、宽、长宽比及千粒重等性状的遗传力分别为0.901 8、0.921 0、0.922 1和0.900 8,单株间的遗传力分别为0.926 0、0.979 2、0.932 6和0.992 5。这说明红榉种子形态性状在种源、单株水平上均受到中等程度以上的遗传控制,为红榉的遗传改良提供了可靠保障。
种子千粒重反映了种子的大小和饱满程度,千粒重越大种子越饱满,其内含的营养物质越丰富,可以提供促发芽的物质越多,使发芽迅速整齐。千粒重对幼苗的生长和生物量也有较大影响。种子质量越大,幼苗越高大,生物量越高[29]。相关分析表明,无论种源间还是单株间,红榉种子千粒重与种子长、种子宽均达到极显著相关关系,表明种子的千粒重性状受种子长、种子宽影响很大,而且有随着种子长、宽的增大而显著递增趋势。因此,可以利用种子千粒重作为选择种子的间接指标。所以可以根据种子的质量来选择红榉不同产地的种子,对早期的红榉育苗具有重要的意义。
红榉各种源种子形态与地理气候因子的相关分析表明,红榉种子长、宽及千粒重之间存在着极显著相关关系,且红榉种子性状无明显的经向和纬向变异,种子长主要受年均温和无霜期影响,种子宽主要受1月均温影响,而千粒重主要受年均温的影响。
红榉种源种子形态性状的聚类分析结果表明,既存在地理分布相近的种源聚为一类的现象,也存在地理分布相隔较远的种源聚为一类的现象。基于上述结果,可以说红榉种子形态特征地理变异在一部分地区呈现区域板块变异模式,还有一部分地区则呈现随机变异模式。但是从遗传学角度出发,种源亲代表型性状的变异是自身遗传变异和外界环境饰变综合作用的结果,单单基于上述结果尚不能完全辨析红榉遗传结构变异模式。因此,对于红榉种源的地理变异模式辨析以及科学的种源区划,还有待于通过种源试验和分子标记手段进一步研究。
[1] 中国科学院植物研究所.中国高等植物图鉴(第一册)[M].北京:科学出版社,1972:469.
[2] 中国植物学会(中华人民共和国国务院正式批准公布).国家重点保护野生植物名录(第一批)[J].植物杂志,1999,5:4-11.
[3] 张立军,周丽君.大叶榉人工栽培技术研究[J].湖南林业科技,1999,26(4):18-23,47.
[4] 汪灵丹,张日清.榉树的研究进展[J].广西林业科学,2005,34(4):188-191,211.
[5] 王旭军,张日清,许忠坤,等.不同种源红榉苗期生长节律的研究[J].中南林业科技大学学报,2013,33(7):31-34.
[6] 刘海龙,张日清,汪灵丹,等.榉树嫩枝扦插技术的研究[J].中南林业科技大学学报,2013,33(9):11-13.
[7] 张日清,刘海龙,汪灵丹,等.榉树无菌播种技术[J].经济林研究,2013,31(1):139-142.
[8] 张日清,杨 婕,金晓玲,等.榉树实生苗苗期内源激素含量的动态变化[J].经济林研究,2011,29(4):1-5.
[9] 汪灵丹,张日清,金晓玲.大叶榉顶芽诱导与增殖培养[J].中南林业科技大学学报,2010,30(6):75-79.
[10] 黄利斌,汪企明,樊丛梅,等.榉树半同胞家系苗期性状变异的研究[J].江苏林业科技,2001,28(6):1-3.
[11] Michelle R L,Westoby M,Jurado E.Correlates of seed size variation:a comparison among fivet emperate floras[J].J.of Eco.,1995,83:517-530.
[12] 李晓洁,徐化成.白皮松种子发芽习性及其种源变异的研究[J].林业科学,1989,25(2):97-104.
[13] 宋丽华,王娅丽.几个臭椿种源种子的生物学特性变异研究[J].农业科学研究,2005,26(1):18-22.
[14] Langlet O.Ecological variability and taxonomy of forest trees [C]//Kozlowskii T T.Tree Growth.Renald Press Co,1962:332-373.
[15] Wright JW.Introduction to forest genetics[M].New York,San Francisco,London:Academic Press,1976:283-331.
[16] Wells O O.Geographic variation in Ponderosa pine[J].Silvae Genet.,1964,13(4):89-103.
[17] 舒 枭,杨志玲,杨 旭,等.不同产地厚朴种子性状的变异分析[J].林业科学研究,2010,23(3):457-461.
[18] 中华人民共和国国家标准.GB2772-1999林木种子检验规程[S].北京:中国标准出版社,2000.
[19] 沈熙环.林木育种学[M].北京:中国林业出版社,1990:55-70.
[20] 程诗明,顾万春.苦楝表型性状梯度变异的研究[J].林业科学,2006,42(5):29-35.
[21] 王军辉,顾万春,万 军,等.桤木不同种源球果及种子性状的遗传变异[J].东北林业大学学报,2006,34(2):1-4.
[22] 佘诚棋,方升佐,杨万霞.青钱柳种子形态特征的地理变异[J].南京林业大学学报:自然科学版,2008,32(4):63-66.
[23] 刘永红,杨培华,韩创举,等.油松不同种源种实性状的变异分析[J].浙江林学院学报,2008,25(2):163-168.
[24] 杨志玲,杨 旭,谭梓峰,等.厚朴不同种源及家系种子性状的变异[J].中南林业科技大学学报,2009,29(5):49-55.
[25] 黄雪方,金雅琴,李冬林.乌桕不同种源种子性状的地理变异[J].西南林业大学学报,2011,31(4):44-48.
[26] 徐化成,孙肇凤.油松种群地理分化的多变量分析[J].林业科学,1984,20(1):9-17.
[27] 徐化成.油松地理变异和种源选择[M].北京:中国林业出版社,1991:15.
[28] 魏胜利,王文全,秦淑英,等.甘草种源种子形态与萌发特性的地理变异研究[J].中国中药杂志,2008,33(8):869-873.
[29] 喻方圆,刘 远.聚乙二醇渗透处理对马尾松种子活力的影响[J].南京林业大学学报,2000,24(1):38-40.
Seed morphological trait variation of Zelkova schneideriana from different provenances
WANG Xu-jun1,ZHANG Ri-qing2,XU Zhong-kun1,CHENG Yong1,WU Ji-you1,ZHANG Jiu-rong3
(1.Hunan Academy of Forestry,Changsha 410004,Hunan,China; 2.Central South University of Forestry and Technology,Changsha 410004,Hunan,China; 3.Sangzhi Forestry Bureau,Sangzhi 427100,Hunan,China)
The variation patterns of four seed morphologic characters were studied by using Zelkova schneideriana Hand-Mazz.seeds as the tested materials,which were from 15 provenances of four provinces in natural distribution.The results show as follows:(1)Highly signi fi cant differences were found among different provenances in seed length,width,ratio of length to width,and 1000-grainweight,and theirs heritability were 0.901 8,0.921 0,0.922 1 and 0.900 8 at the provenance level,and 0.926 0,0.979 2,0.932 6 and 0.9925 at the individual level,respectively; this indicated that in these seed morphologic traits there were abundant genetic variation,and were controlled by moderate-higher genetic control.(2)There were highly signi fi cant correlation among seed length,seed width and 1000-grain-weight,and these seed morphologic traits demonstrated no obvious longitude or latitude variation tendency,the seed length was mainly affected by annual average temperature and frostless days,the seed width was affected by average temperature in January,While 1000-grain-weight was affected by annual average temperature.(3)Cluster analysis suggested that the geographic variation of seed morphologic traits of some distribution zones of Z.schneideriana represented regional block variation pattern,while the others showed random variation pattern.
Zelkova schneideriana Hand-Mazz.; provenance; seed morphologic trait; variation
S792.11
A
1673-923X(2015)01-0001-07
10.14067/j.cnki.1673-923x.2015.01.001
2013-11-15
国家林业公益性行业科研专项(200904011);国家“十二五”科技支撑计划项目(2011BAD38B03)
王旭军,副研究员,博士;E-mail:xjwang0514@163.com
王旭军,张日清,许忠坤,等.红榉不同种源种子形态性状变异[J].中南林业科技大学学报,2015,35(1):1-7.
[本文编校:谢荣秀]
猜你喜欢
南方农业(2022年17期)2022-10-21
养猪(2022年4期)2022-08-17
农业科技通讯(2022年7期)2022-07-19
安徽农学通报(2022年6期)2022-04-07
绿色科技(2021年21期)2021-11-26
辽宁林业科技(2021年1期)2021-03-16
农家科技中旬版(2020年4期)2020-07-06
广西农学报(2019年1期)2019-07-18
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
