
不同梭梭种群同化枝的解剖结构特征及其与生态因子的关系分析
2016-03-24 06:32昝丹丹吕新华
西北植物学报 2016年2期
关键词:梭梭
昝丹丹,庄 丽,黄 刚,刘 鸯,吕新华
(石河子大学 生命科学学院,新疆石河子 832000)
不同梭梭种群同化枝的解剖结构特征及其与生态因子的关系分析
昝丹丹,庄丽*,黄刚,刘鸯,吕新华
(石河子大学 生命科学学院,新疆石河子 832000)
摘要:该研究于中国西北干旱区按自然降水梯度从东向西选择6个梭梭种群,取其当年生同化枝作为试验材料,采用石蜡切片法,应用光学显微技术测定同化枝9项解剖结构指标,分析不同生境下梭梭同化枝解剖结构的变异特点以及与生态因子的关系,为干旱区生态环境保护和梭梭资源的保护与利用提供理论依据。结果表明:(1)干旱荒漠环境的梭梭同化枝解剖结构具有连续的栅栏组织、花环结构、含晶细胞和贮水组织等结构特征,但其表皮细胞、角质层厚度、气孔位置和气下室与旱生植物结构有差异,主要表现为表皮细胞和角质层较薄,气孔半下陷且气下室不发达。(2)梭梭同化枝各种群间的9项解剖结构指标都存在显著差异,其中变异系数较大的是维管束个数/直径和角质层厚度,变异系数分别为22.78%和15.20%;相关分析表明:同化枝直径、栅栏细胞切向长、贮水组织厚度和维管柱直径均与经度、纬度、海拔、一月均温、七月均温、年均温、年降水量和年相对湿度等8个生态因子有不同程度的相关性,其中与经度、海拔、七月均温和年平均相对湿度显著相关。(3)6个梭梭种群通过聚类分析可分为两大类,分析结果与所观察到的生境类型、地理分布的划分基本一致;随着自然降水量自东向西减少,梭梭的抗旱性相应逐渐增强。
关键词:西北干旱区;梭梭;同化枝;解剖结构;生态因子
梭梭(Haloxylonammodendron)属藜科(Chenopodiaceae)梭梭属(HaloxylonBunge)超旱生小乔木,呈高大灌木状,为国家濒危三级保护植物[1]。以梭梭为建群种的梭梭荒漠,在亚、非荒漠区有大面积分布,并形成水平地带性独特景观。在中国梭梭荒漠主要分布在西北荒漠地区,约占全国荒漠总面积(不包括山地)的十分之一[2-3]。梭梭主侧根发达,叶退化为同化枝进行光合作用,对干旱有较强的适应性,是中国西部荒漠化地区植树造林首选树种,也是极好的薪炭材和饲用植物。近几年中国学者对梭梭的研究多集中在分类学[4-6],分子生物学[7-9]及生理生态学[10-15]等方面,早期对该植物同化枝解剖结构的研究己有报道[16-18],但针对不同种群间梭梭同化枝解剖结构比较的研究未见报道。
植物叶片是植物进行光合作用和呼吸作用的重要器官,与周围环境紧密联系,是植物体暴露于大气环境中面积最大的器官,对水分、温度、光照、降水量、海拔等环境因子的变化敏感、可塑性大,主要表现为叶形的变化、叶片的大小、厚度及解剖结构的不同,而叶片的形态结构特征是植物适应环境的具体表现[19]。叶片结构在不同种类的植物之间具有明显的差异,由于环境条件的不同,同种植物叶片的解剖结构表现出明显的差异[20]。同化枝是一种特殊的光合器官,常常存在于适应干旱环境的植物中,是旱生结构的主要特征之一。同化枝为了减少蒸腾面积,叶片退化为膜质鳞片,取代叶进行光合作用;幼茎呈绿色,肉质特化[21]。由于同化枝是介于叶与茎之间,具有独特的结构特点,本研究通过对不同生境下梭梭同化枝在解剖结构的变化进行研究,分析了梭梭同化枝结构变异的特点及与生态因子的相关性,旨在于揭示其与生态因子的关系,为干旱区生态环境保护和梭梭资源的保护与利用提供理论依据。
1材料和方法
1.1实验材料
2014年8月,以内蒙古阿拉善、甘肃民勤、新疆奇台、阜康、石河子和精河为实验材料采集地,以生长健康且基径、株高、冠幅基本一致的梭梭植株为取材对象,采用随机抽样,6个梭梭自然种群各采9株,比较不同种群梭梭同化枝解剖结构。采样点的经纬度和海拔用GPS定位记录,均温、降水量及相对湿度等生态因子数据由当地相关气象单位提供,表1中气象数据为3年数据的均值。
1.2同化枝解剖结构的测定
用解剖刀截取位于同一高度的当年生同化枝,截成5 mm小段后立即放入FAA固定液(酒精∶福尔马林∶冰醋酸=90∶5∶5)中进行固定,以保证植物组织结构完整,固定时间为24 h以上。取固定好的同化枝进行脱水、透明、浸蜡、包埋,用Leica切片机横向切割同化枝,将得到的石蜡切片进行番红-固绿对染,中性树胶封片[22],在Olympus BX51显微镜下观察拍照,选择有代表性的石蜡切片,记录同化枝的横切面特征。用Digimizer测量软件分别测量同化枝直径、角质层厚度、栅栏细胞径向长、栅栏细胞切向长、栅栏细胞密度、维管柱厚度等指标。测量值为10个数值的平均值。
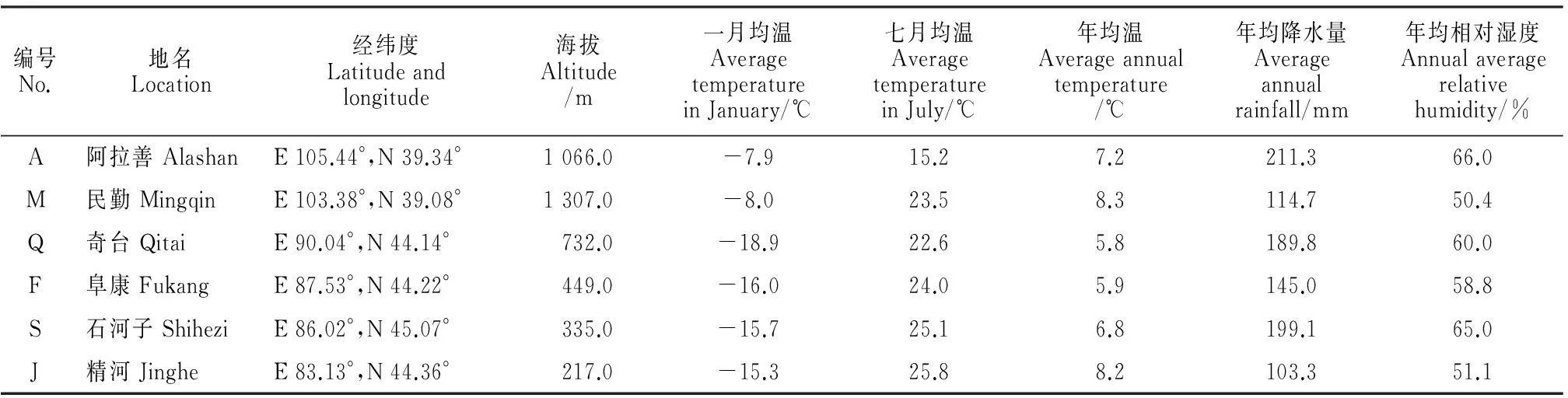
表1 供试材料来源
1.3数据处理
采用Microsoft Excel 2010和SPSS 20.0软件进行统计分析。单因素方差分析法分析各种群结构指标的大小和差异性;采用双变量相关分析法对同化枝解剖结构指标与生态因子之间进行相关分析;利用类平均法(UPGMA)进行分层聚类分析。
2结果与分析
2.1同化枝解剖结构特征
6个梭梭种群的同化枝横切面结构相似,均呈近圆形(图1,a~f),具有以下典型的旱生特征。梭梭的叶退化,只保有瘤状突起,由同化枝代替叶进行光合作用。当年生同化枝呈淡绿色,具节和节间。梭梭同化枝解剖结构由表皮、皮层和维管柱3部分组成,其中表皮光滑,无被毛,表皮细胞的形状差异不大,排列紧密;角质层较薄,最大厚度仅有1.46 μm;表皮上有半下陷的气孔分布,气孔下分布有气室但不发达(图1,g)。表皮下有1层排列疏松的下皮细胞;下皮细胞内侧的栅栏组织由单层形状细长的栅栏细胞紧密排列组成,其中富含叶绿素。栅栏组织内侧是一层由排列紧密的粘液细胞组成的花环细胞,其内含叶绿体整个结构呈波浪形轮廓(图1,h)。花环细胞内侧是起着贮水保水作用的贮水组织,贮水组织中央是大维管柱,其外套一圈厚壁组织,木质部导管孔径小,贮水组织中散生小的维管束;髓射线狭窄,不发达。在栅栏组织、贮水组织和髓内均分布含晶细胞(图1,i~l)。
2.26个梭梭种群的同化枝解剖结构特征比较
对不同梭梭种群的解剖结构特征进行比较(表2),其中除导管孔径之外,其他特征都具有显著差异(P<0.05)。同化枝直径变化范围是506.47~785.9 μm,变异系数为7.42%,最大的是J种群,最小的是M种群。角质层厚度最大的是J群,最小的是A种群,变化范围是1.18~1.46 μm;栅栏细胞径向长最长的是F种群,长度为15.66 μm,最短的是S种群,长度为9.51 μm;A种群栅栏细胞切向长最短,S种群的最长;栅栏细胞密度在8~13个/μm,最大的是M种群,最小的是F种群;3个指标中栅栏细胞切向长的变异系数13.62%大于另外2个;小维管束个数/直径的变化范围是5~9个,其中变异系数为22.78%;贮水组织厚度的变化范围119.52~211.04 μm,维管柱直径变化范围139.15~269.28 μm,J种群的这2个指标均为最大,A种群的贮水组织厚度最薄,F种群维管柱直径最小;不同种群的导管孔径无显著差异。
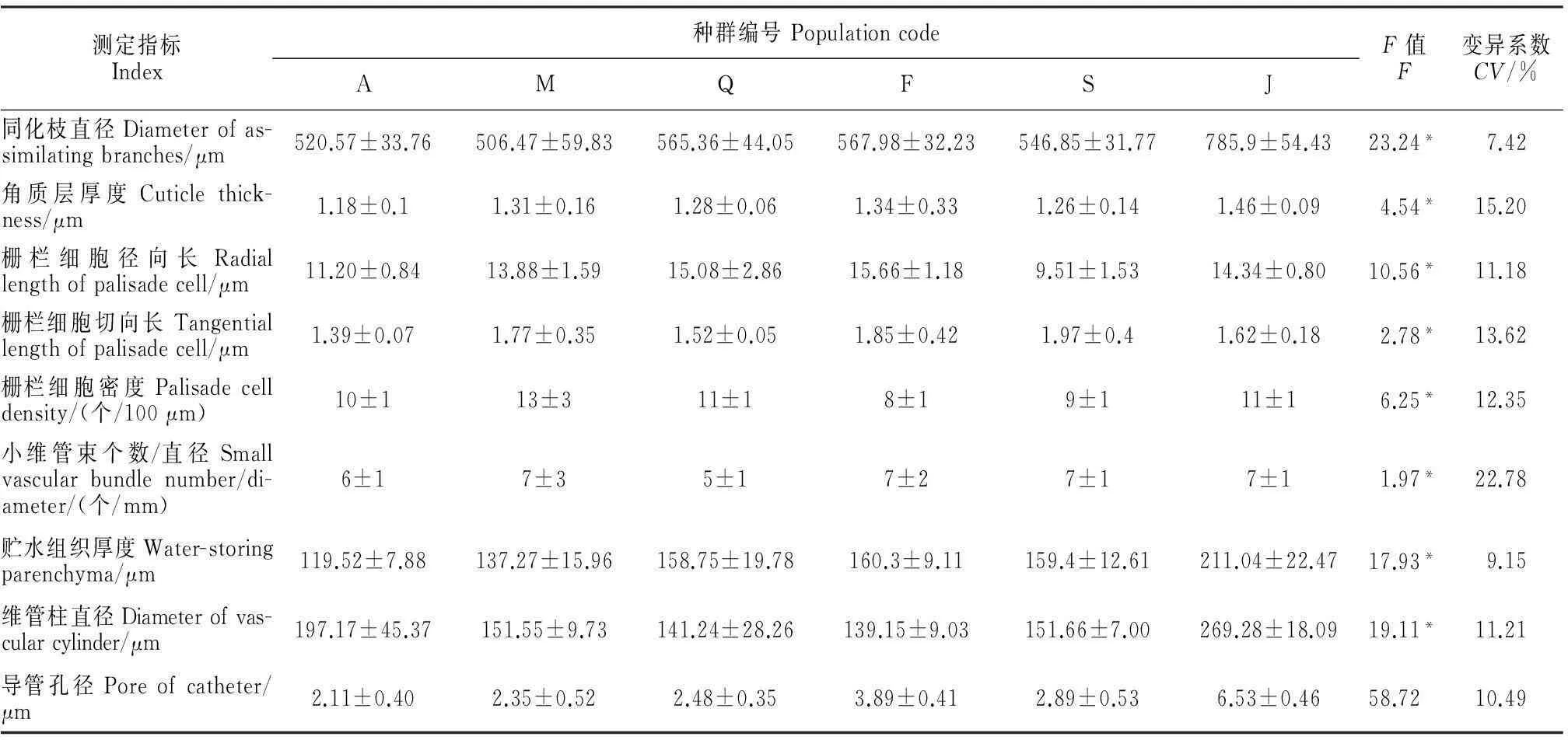
表2 6个种群梭梭同化枝解剖结构特征值及方差分析
注:数据为平均数±标准差;*表示差异显著性(P<0.05)。
Note:Value is mean±SD;* indicates the significant difference(P<0.05).
2.3同化枝解剖结构指标与生态因子的相关关系
对所测定的解剖结构的9个指标与8个生态因子进行双变量相关分析,结果(表3)表明:同化枝直径与七月均温以及年均相对湿度呈显著负相关,这说明随着七月均温和年均相对湿度的升高,同化枝直径减小;栅栏细胞切向长与七月均温呈极显著正相关关系,维管柱直径与七月均温呈显著正相关,与年平均相对湿度呈极显著正相关关系,说明随着七月均温和年平均相对湿度的增加,栅栏细胞切向长与维管柱直径增大;贮水组织厚度与经度和海拔呈极显著负相关关系,即随着经度以及海拔的降低,贮水组织厚度呈现升高趋势,符合植物抗旱性特点。
2.4不同梭梭种群的聚类分析
根据同化枝的解剖指标特征(表2),利用类平均法(UPGMA)对6个梭梭种群进行聚类分析,在欧式距离为5时,这些梭梭种群聚类为两大类(图2),Q、F、S三个居群先聚合在一起,紧接着与A和M种群相聚,这5个种群聚为第一类;这一类所处地生境主要为沙壤土,年均降水量接近,同化枝的解剖特征主要表现为栅栏细胞径向长和栅栏细胞切向长较长以及栅栏细胞密度较大。第二类为J种群,J种群所在地的生境为盐碱沙土,年均降水量为103.3 mm;此类的特点是角质层厚度和贮水组织厚度较厚,同化枝直径、维管柱直径以及导管孔径较大。
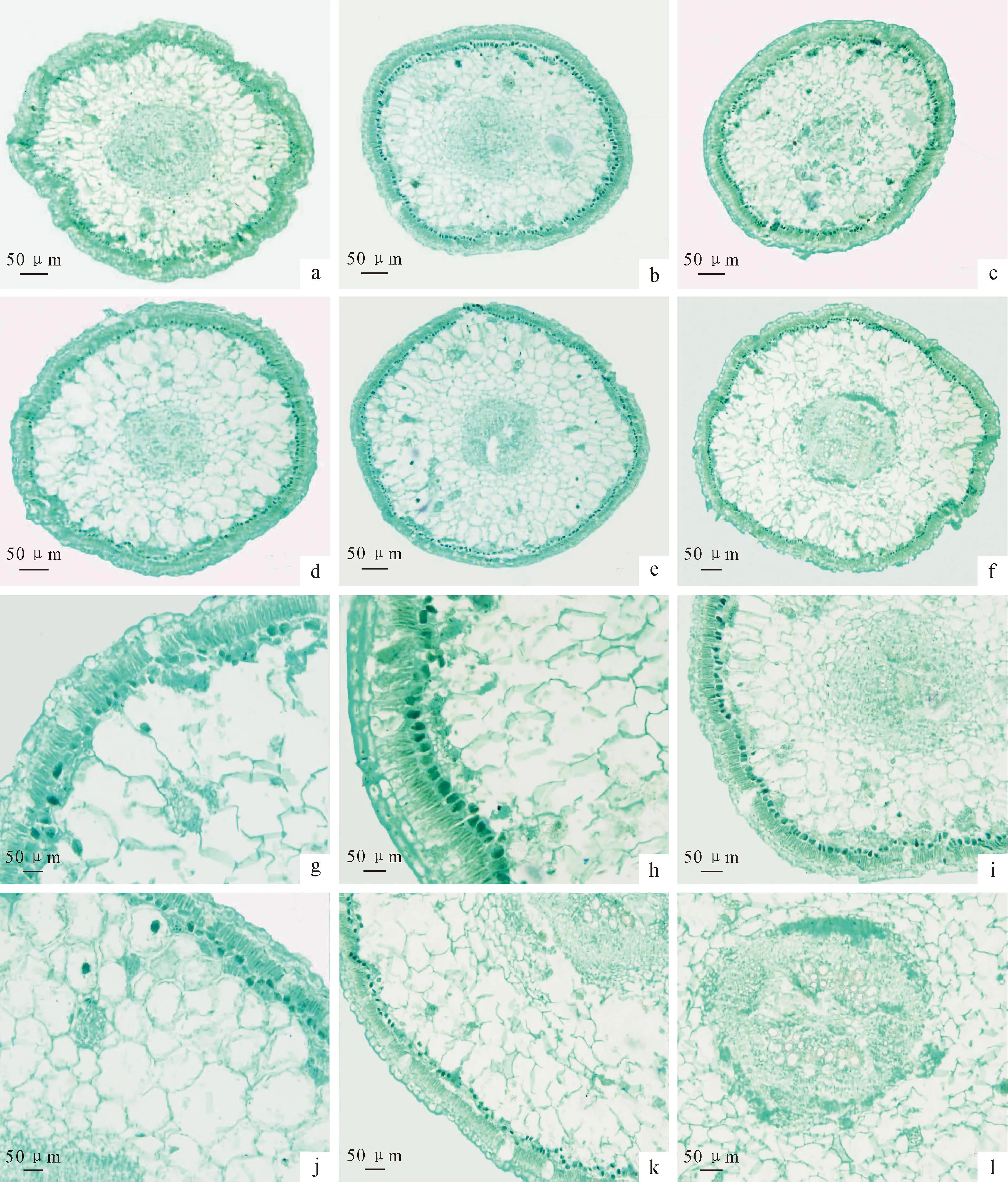
梭梭同化枝横切面的完整结构:a~f分别代表A、M、Q、F、S、J种群;g~l.局部示意图

测定指标Index经度Latitude纬度Longitude海拔Altitude一月均温AveragetemperatureinJanuary七月均温AveragetemperatureinJuly年均温Averageannualtemperature年均降水量Averageannualrainfall年均相对湿度Annualaveragerelativehumidity同化枝直径Diameterofassimilatingbranches-0.6680.463-0.682-0.358-1.000*-0.551-0.551-1.000*角质层厚度Cuticlethickness-0.2390.268-0.267-0.249-0.629-0.166-0.621-0.156栅栏细胞径向长Radiallengthofpalisadecell-0.1950.114-0.014-0.320-0.660-0.586-0.586-0.660栅栏细胞切向长Tangentiallengthofpalisadecell-0.6640.626-0.433-0.6050.966**-0.539-0.539-0.542栅栏细胞密度Densityofpalisadecell0.433-0.5420.6000.431-0.034-0.428-0.428-0.034小维管束个数/直径Smallvascularbundlenumber/diameter-0.1940.044-0.2640.2600.359-0.571-0.5710.359贮水组织厚度Water-storingparenchyma-0.861*0.698-0.803*-0.593-0.6290.565-0.566-0.629维管柱直径Diameterofvascularcylinder-0.198-0.026-0.3200.1621.000*-0.394-0.3941.000**导管孔径Poreofcatheter-0.7440.4830.332-0.3030.5210.341-0.621-0.156
注:*表示0.05水平相关性显著;**表示0.01水平相关性极显著。
Note:* indicates the significant correlation(P<0.05);** indicate the extremely significant correlation(P<0.01).

A、M、Q、F、S、J表示梭梭种群编号;0~25表示不同类之间的距离
3讨论
3.1同化枝解剖结构的差异性
植物的生长与分布是植物对外界环境长期适应的结果,为了更好地适应高温、缺水等干旱环境,植物叶片不仅在内部的生理生化特点产生变化,在自身的形态结构特征均具有相应的变异[23]。不同的环境会导致叶的厚度、栅栏组织、海绵组织、胞间隙等形态的改变,其中叶片厚度是植物保水性能的重要指标。栅栏组织细胞因含有大量的叶绿体,因此栅栏组织的厚度以及与叶片厚度的比值是植物光合作用能力强弱的指标。维管束在植物体中起到营养物质的运输和机械支持作用,因此,维管束越发达,植物的生命力越旺盛,植物的抗逆性越强[24-25]。文中所选的梭梭同化枝取代叶行使同化功能,是梭梭进化的顶峰。梭梭同化枝在结构上与环栅型叶有许多共同点,表皮细胞表皮外有角质层,连续紧密的栅栏组织、花环结构、粘液细胞、含晶细胞,贮水组织等解剖特征不仅可以有效地提高光合效率,同时可使体内的水势始终低于土壤水势,防止细胞组织失水,提高植物的保水和吸水力,在一定程度上增强了同化枝的耐旱性。梭梭表皮细胞和角质层较薄,且气孔半下陷,虽与旱生植物的结构有差异,但因为梭梭是典型的超旱生植物,其必定有适应荒漠环境的特殊机制。以上结果与公维昌[26]对梭梭植物同化枝的解剖学研究结果相似。
本研究发现,采自同一时间段(2014年8月)的不同梭梭种群其同化枝解剖结构特征指数具有明显的差异,从侧面反映了梭梭具有适应不同生境的能力。梭梭的同化枝解剖结构特征受环境影响较大,在9个解剖结构特征中,角质层厚度和小维管束个数/直径差异最大,变异系数分别为15.2%和22.78%,表明在不同环境条件选择下,这两个特征的可塑性最大;而同化枝直径和贮水组织厚度的差异相对较小,变异系数分别为7.42%和9.15%,表明这2个特征相对稳定,受环境条件的影响不大。
3.2同化枝解剖结构与生态因子的关系
在长久的进化过程中,每一种植物存在于一个适合其生长发育的地理环境内,形成了与生态因子(如温度、降水、光照等)相互适应的分布格局。通常认为,决定大尺度植被分布格局及植物形态结构差异的因素主要是温度和降水等气候因子。而在小尺度上,影响植物形态结构差异主要因素是微环境,如地形条件(海拔)、土壤水分条件及其他生物因素[27-29]。本研究采用相关分析法分析梭梭同化枝解剖结构与生态因子关系的研究与前人的研究有相同的结果。即在经度、纬度、海拔、一月均温、七月均温、年均温、年降水量和年平均相对湿度8个生态因子中,每个解剖结构指标都与这些因子有着不同程度的相关性,那么必定有一种或多种同化枝结构与这些因子呈现相关关系。本研究结果显示随着七月均温和年均相对湿度的升高,同化枝直径减小,栅栏细胞切向长与维管柱直径增大;贮水组织厚度随着经度和海拔的降低而呈现升高趋势。其中同化枝直径、维管柱直径和贮水组织厚度与经度、海拔和七月均温有显著相关关系,栅栏细胞切向长和维管柱直径与七月均温和年平均相对湿度有极显著相关关系;因此推测这几个结构指标与梭梭的抗旱特性有关,为梭梭抗旱结构的主要特征,而七月均温、年均相对湿度、经度和海拔是影响同化枝结构的关键因子。
3.3不同梭梭种群的抗旱性聚类分析
按照邢毅等[30]研究得出的具有相似生境特点的种群具有相似表型特征,本研究采用聚类方法对梭梭同化枝的解剖研究结果进行了聚类,最终这6个种群分为两大类,第一大类包括A、M、F、Q、S五个种群,表现出相似性,其主要特点是;栅栏细胞径向长、栅栏细胞切向长以及栅栏细胞密度较大;其生境是荒漠地区,生长在沙垄之上;土壤为沙壤土,年均降水量接近;第2类是J种群,该种群的生境是平原戈壁,土壤主要为盐碱沙土和戈壁,其同化枝直径、角质层厚度、贮水组织厚度、维管柱直径以及导管孔径都较大。通常认为,同化枝的角质层厚度、贮水组织厚度和维管柱直径越厚,抗旱性越强,从这个角度来说,J种群的抗旱能力较其他种群强,这有待于进行抗旱生理试验来验证。
4结论
(1)梭梭同化枝解剖结构对干旱荒漠环境的积极适应性,主要表现为具有连续的栅栏组织、花环结构、含晶细胞和贮水组织等结构;但其表皮细胞、角质层厚度、气孔位置和气下室与旱生植物结构有差异,主要表现在表皮细胞和角质层较薄,气孔半下陷且气下室不发达。
(2)不同种群梭梭同化枝解剖结构具有差异性。同化枝直径、栅栏细胞切向长、维管柱直径和贮水组织厚度与经度、海拔、七月均温和年平均相对湿度有显著相关关系,这4个指标与梭梭的抗旱特性有关,为梭梭抗旱结构的主要特征。
(3)6个梭梭种群的聚类结果与所观察到生境类型、地理分布的划分基本一致。按自然降水梯度自东向西,随着降雨量的减少,梭梭的抗旱性增强。
参考文献:
[1]国家环境保护局自然保护司保护区与物种管理处.珍稀濒危植物保护与研究[M].北京:中国环境科学出版社,1991:157-170.
[2]郭泉水,郭志华,阎洪,等.我国以梭梭属植物为优势的潜在荒漠植被分布[J].生态学报,2005,25(4):848-853.
GUO Q S,GUO Z H,YAN H,etal.Study on potential distribution ofHaloxylonplants dominated desert vegetation in China[J].ActaEcologicaSinica,2005,25(4):848-853.
[3]胡式之.中国西北地区的梭梭荒漠[J].植物生态学与地植物学丛刊,1963,1(1):81-109.
HU S Z.HaloxylonDesert in Northwest China[J].PlantEcologyandGeobotanicaSinica,1963,1(1):81-109.
[4]努尔古丽·阿木提.新疆藜科植物系统分类学研究[D].乌鲁木齐:新疆大学,2013.
[5]何静,高翔,吕光辉,等.艾比湖自然保护区植物群落的数量分类及土壤环境解释[J].新疆农业科学,2010,5:1 030-1 035.
HE J,GAO X,LÜ G H,etal.The numerical classification of vegetation community and soil interpretation of Nature Reserve in the Ebinur Take Wetland[J].XinjiangAgriculturalSciences,2010,5:1 030-1 035.
[6]周志强,魏晓雪,刘彤.新疆奇台荒漠植物群落的数量分类及土壤环境解释[J].生物多样性,2007,3:264-270.
ZHOU Z Q,WEI X X,LIU T.The numerical classification of desert vegetation and soil interpretation in Qitai County,Xinjiang[J].BiodiversityScience,2007,15(3):264-270.
[7]甘晓燕,巩檑,石磊,等.梭梭液泡膜焦磷酸酶HaVVP基因克隆及序列分析[J].西北农业学报,2014,23(11):198-203.
GAN X Y,GONG L,SHI L,etal.Cloning and sequence analysis of a cDNA encoding vacuolar H+-PPase gene fromHaloxylonammodendron[J].ActaBot.Boreal.-Occident.Sin.,2014,23(11):198-203.
[8]张萍,董玉芝,魏岩,等.利用ISSR标记对新疆梭梭遗传多样性的研究[J].西北植物学报,2006,26(7):1 337-1 341.
ZHANG P,DONG Y Z,etal.ISSR analysis of genetic diversity ofHaloxylonammodendron(C.A.Mey.) Bunge in Xinjiang[J].ActaBot.Boreal.-Occident.Sin.,2006,26(7):1 337-1 341.
[9]石磊,甘晓燕,夏兴雷,等.梭梭CMO基因的克隆与植物表达载体的构建[J].西北植物学报,2010,30(8):1 514-1 519.
SHI L,GAN X Y,XIA X L,etal.Cloning of choline monooxygenase gene fromHaloxylonammodendronand construction of its plant expression vector[J].ActaBot.Boreal.-Occident.Sin.,2010,30(8):1 514-1 519.
[10]巩檑,甘晓燕,石磊,等.梭梭NAC转录因子HaNAC1克隆及序列分析[J].西北农业学报,2014,23(12):168-174.
GONG L,GAN X Y,SHI L,etal.Cloning and sequence analysis of a NAC-like gene HaNAC1 fromHaloxylonammodendron[J].ActaAgriculturaeBoreali-OccidentalisSinica,2014,23(12):168-174.
[11]郭泉水,谭德远,刘玉军,等.梭梭对干旱的适应及抗旱机理研究进展[J].林业科学研究,2004,17(6):796-803.
GUO Q S,TAN D Y,LIU Y J,etal.Advance in studies ofHaloxylonBunge’s mechanism of adapation and resistance to drought[J].ForestResearch,2004,17(6):796-803.
[12]陈芳,纪永福,张锦春,等.民勤梭梭人工林天然更新的生态条件[J].生态学杂志,2010,29(9):1 691-1 695.
CHEN F,JI Y F,etal.Ecological conditions for natural regeneration of artificialHaloxylonplantationsin Minqin oasis[J].ChineseJournalofEcology,2010,29(9):1 691-1 695.
[13]史胜青,齐力旺,孙晓梅,等.梭梭抗旱性相关研究现状及对今后研究的建议[J].世界林业研究,2006,19(5):27-32.
SHI S Q,QI L W,SUN X M,etal.Research progress and suggestion on drought resistance ofHaloxylonammodendron[J].WorldForestryResearch,2006,19(5):27-32.
[14]格日勒,梅花,吉如和,等.阿拉善荒漠梭梭资源状况与恢复利用[J].草原与草业,2014,26(4):18-21.
GE R L,MEI H,JI R H,etal.Plant ecology and geobotany series alxa desertHaloxylonresourcesutilization and recovery[J].GrasslandandPrataculture,2014,26(4):18-21.
[15]楚新正,马倩,马晓飞,等.梭梭(Haloxylonammodendron)主根周围土壤特征[J].中国沙漠,2014,34(1):170-175.
CHU X ZH,MA Q,MA X F,etal.Characteristics of the soil around the main root ofHaloxylonammodendron[J].JournalofDesertResearch,2014,34(1):170-175.
[16]罗秀英,邓彦斌.新疆几种旱生植物叶(同化枝)的解剖结构观察[J].新疆大学学报(自然科学版),1986,1:77-84.
LUO X Y,DENG Y B.Anatomical observation of leaves and assimilative branches on several xerophytes structure in Xinjiang[J].JournalofXinjiangUniversity(Nat.Sci.Edi.),1986,1:77-84.
[17]刘家琼.我国荒漠不同生态类型植物的旱生结构[J].植物生态学与地植物学丛刊,1982,6(4):314-325.
LIU J Q.The xeromorphic structure of different typical plants in deserts of China[J].ActaPhytoecologicaetGeobotanicaSinica,1982,6(4):314-325.
[18]董占元,姚云峰,赵金仁,等.梭梭[Haloxylonammodendron(C.A.Mey) Bunge]光合枝细胞组织学观察及其抗逆性特征[J].干旱区资源与环境,2000,14(S1):78-83.
DONG ZH Y,YAO Y F,ZHAO J R,etal.Anatomical observations on the plotosynthatic branch ofHaloxylonammodendrom(C.A.Mey) Bunge and It is the character of drought and salt resistance[J].JournalofAridLandResourcesandEnvironment,2000,14(S1):78-83.
[19]公维昌,庄丽,赵文勤,等.多枝柽柳与梭梭光合器官形态解剖结构的生态适应性[J].中国沙漠,2011,31(1):129-136.
GONG W C,ZHUANG L,ZHAO W Q,etal.Ecological adaptation of morphological and anatomical structure of photosynthetic organs ofTamarixramosissimaandHaloxylonammodendron[J].JournalofDesertResearch,2011,31(1):129-136.
[20]周朝彬,辛慧慧,宋于洋.梭梭次生木质部解剖特征及其可塑性研究[J].西北林学院学报,2014,29(2):207-212.
ZHOU C B,XIN H H,etal.Secondary xylem anatomical structure and its plasticity ofHaloxylonammodendron[J].JournalofNorthwestForestryUniversity,2014,29(2):207-212.
[21]闻志彬,张明理.30种藜科C4植物同化器官解剖结构及δ13C值[J].中国沙漠,2014,34(4):1 015-1 022.
WEN ZH B,ZHANG M L.Anatomical types and13C/12C values of assimilating organs in 30 C4species from chenopodiaceae in Xinjiang,China[J].JournalofDesertResearch,2014,34(4):1 015-1 022.
[22]赵小仙,李毅,苏世平,等.3个地理种群蒙古沙拐枣同化枝解剖结构及抗旱性比较[J].中国沙漠,2014,34(5):1 293-1 300.
ZHAO X X,LI Y,SU S P,etal.Drought resistance analysis based on anatomical structures of assimilating shoots ofCalligonummongolicumfrom three geographic populations[J].JournalofDesertResearch,2014,34(5):1 293-1 300.
[23]王勋陵,王静.植物的形态结构与环境[M].兰州:兰州大学出版社,1989:57-87.
[24]周仪.植物形态解剖实验(修订版)[M].北京:北京师范大学出版社,1993:37-68.
[25]郗金标,张福锁,田长彦.新疆盐生植物[M].北京:科学出版社,2006:7-60.
[26]公维昌,庄丽,赵文勤,等.2种盐生植物解剖结构的生态适应性[J].生态学报,2009,29(12):6 764-6 771.
GONG W C,ZHUANG L,ZHAO W Q,etal.Anatan ical structure and ecological adaptability of two kinds of halophytes (HaloxylonammondendronChenopodiaceae andTamarixramosissiaAmaran thaceae)[J].ActaEcologicaSinica,2009,29(12):6 764-6 771.
[27]康萨如拉,牛建明,张庆,等.短花针茅叶片解剖结构及与气候因子的关系[J].草业学报,2013,22(1):77-86.
KANGSHA R L,NIU J M,etal.Anatomical structure ofStipabrevifloraleaves and its relationship with environmental factors[J].ActaPrataculturaeSinica,2013,22(1):77-86.
[28]胡建莹,郭柯,董鸣.高寒草原优势种叶片结构变化与生态因子的关系[J].植物生态学报,2008,32(2):370-378.
HU J Y,GUO K,DONG M.Variation of leaf structure of two dominant species in alpine grassland and the relationship between leaf structure and ecological factors[J].JournalofPlantEcology(Chinese Version),2008,32(2):370-378.
[29]邓彦斌,姜彦成,刘健.新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究[J].植物生态学报,1988,12(2):69-75.
DENG Y B,JIANG Y CH,LIU J.The xeromorphic and saline morphic structure of leaves and assimilating branches in ten Chenopodiacea species in Xinjiang[J].ActaPhytoecologicaSinica,1988,12(2):69-75.
[30]邢毅,赵祥,董宽虎,等.不同居群达乌里胡枝子形态变异研究[J].草业学报,2008,17(4):26-31.
XING Y,ZHAO X,DONG K H,etal.A study on morphological variation of different populations ofLespedezadavurica[J].ActaPratacultureSinica,2008,17(4):26-31.
(编辑:潘新社)
Relationship between Anatomical Differences and Ecological Factors of the Assimilating Shoots ofHaloxylonammodendron
ZAN Dandan,ZHUANG Li*,HUANG Gang,LIU Yang,LÜ Xinhua
(College of Life Science,Shihezi University,Shehezi,Xinjiang 832000,China)
Abstract:The annual assimilating shoots of 6 Haloxylon ammodendron species were selected as experimental materials,which were gotten from arid region of northwest China according to precipitation gradient which is from east to west.9 anatomical structures were measured.We analyzed the relationship between anatomical differences and ecological factors of the assimilating shoots of H.ammodendron,to provide theoretical basis for the protection and utilization of the ecological environment and H.ammodendron resources in arid areas of China.Results showed that:(1)the anatomical structure of the assimilating shoots of H.ammodendron in the arid desert environment with continuous wreath of palisade tissue,structure,crystal cell and water storage organization structure,but the epidermis cells,cutin layer thickness,hole position under the gas chamber difference with xerophyte structure,mainly displays in the epidermal cells and corneous layer is thinner,porosity nuder half wubsidence and gas chamber is not developed.(2)9 anatomical structures of the assimilating shoots of each characteristic of 6 H.ammodendron species have significant differences by variance analysis.The variability coefficients are greater in the number of vascular bundle and cuticle thickness with the values of 22.78% and 15.20%,respectively;Correlation analysis showed that each index has different degree of correlation with eight ecological factors,which are longitude,latitude,altitude,average temperature in January and July,annual average temperature,annual precipitation and annual average relative humidity.The diameter of assimilating shoots thickness,palisade cell cut to length,water storage tissue thickness and vascular cylinder diameter have different degrees of correlation with eight ecological factors.(3)6 populations of H.ammodendron could be divided into 2 categories by cluster analysis.The results of cluster analysis were identical with the habitat types and geographical distribution of H.ammodendron;as fewer natural rainfall from east to west,the drought resistance of H.ammodendron gradually strengthened.
Key words:arid area of northwest China;Haloxylon ammodendron;assimilating shoots;anatomical structure;ecological factors
中图分类号:Q944.56;Q948.11
文献标志码:A
作者简介:昝丹丹(1989-),女,硕士研究生,主要从事资源植物研究。E-mail:827444294@qq.com*通信作者:庄丽,教授,主要从事资源植物与生态保护方面的研究。E-mail:2230688993@qq.com
基金项目:国家重点基础研究发展计划(973项目)(2014CB954203);国家自然科学基金(31360139,41561010,31560177,31460070)
收稿日期:2015-11-04;修改稿收到日期:2016-01-06
文章编号:1000-4025(2016)02-0309-07
doi:10.7606/j.issn.1000-4025.2016.02.0309
猜你喜欢
世界热带农业信息(2022年7期)2022-06-30
哈哈画报(2022年4期)2022-04-19
科学大众(2020年17期)2020-10-27
农民致富之友(2020年12期)2020-05-11
中成药(2020年4期)2020-04-29
生态学报(2020年6期)2020-04-20
今日中国·中文版(2020年3期)2020-03-09
中学时代(2019年7期)2019-11-13
中学生天地(B版)(2019年4期)2019-05-08
绿色中国(2015年6期)2015-07-03
